Wind Resistance of Eastern Baltic Silver Birch (Betula pendula Roth.) Suggests Its Suitability for Periodically Waterlogged Sites

 , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Sample Trees
2.2. Static Pulling Tests
2.3. Data Processing and Analysis
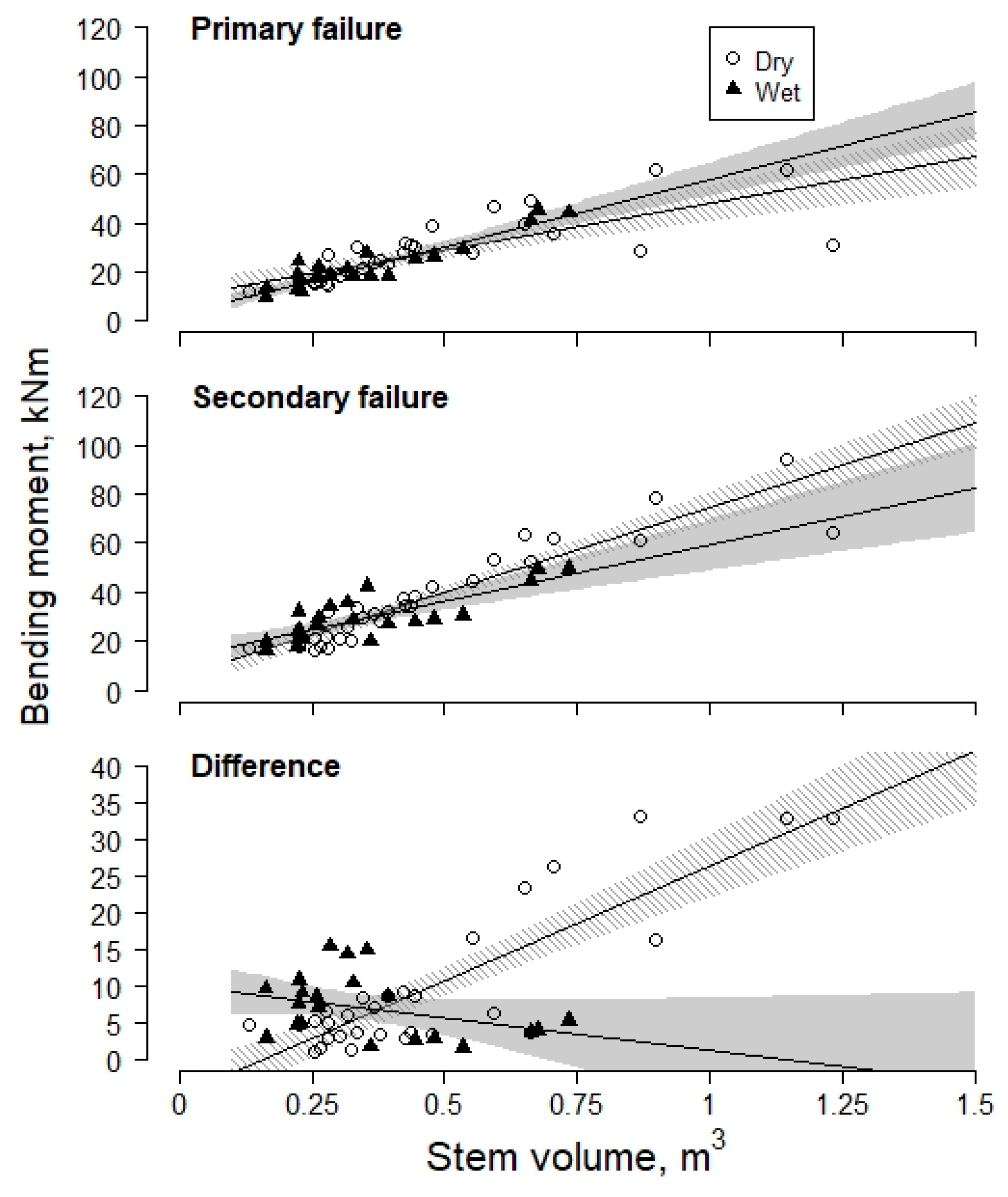
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest disturbances under climate change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef]
- Csilléry, K.; Kunstler, G.; Courbaud, B.; Allard, D.; Lassègues, P.; Haslinger, K.; Gardiner, B. Coupled effects of wind-storms and drought on tree mortality across 115 forest stands from the Western Alps and the Jura mountains. Glob. Chang. Biol. 2017, 23, 5092–5107. [Google Scholar] [CrossRef]
- Cawley, K.M.; Campbell, J.; Zwilling, M.; Jaffé, R. Evaluation of forest disturbance legacy effects on dissolved organic matter characteristics in streams at the Hubbard Brook Experimental Forest, New Hampshire. Aquat. Sci. 2014, 76, 611–622. [Google Scholar] [CrossRef]
- Gardiner, B.; Berry, P.; Moulia, B. Review: Wind impacts on plant growth, mechanics and damage. Plant Sci. 2016, 245, 94–118. [Google Scholar] [CrossRef] [PubMed]
- Laapas, M.; Lehtonen, I.; Venäläinen, A.; Peltola, H.M. The 10-Year return levels of maximumwind speeds under frozen and unfrozen soil forest conditions in Finland. Climate 2019, 7, 62. [Google Scholar] [CrossRef]
- Venäläinen, A.; Lehtonen, I.; Laapas, M.; Ruosteenoja, K.; Tikkanen, O.P.; Viiri, H.; Ikonen, V.P.; Peltola, H. Climate change induces multiple risks to boreal forests and forestry in Finland: A literature review. Glob. Chang. Biol. 2020, 26, 4178–4196. [Google Scholar] [CrossRef] [PubMed]
- Jackson, T.; Shenkin, A.; Moore, J.; Bunce, A.; Van Emmerik, T.; Kane, B.; Burcham, D.; James, K.; Selker, J.; Calders, K.; et al. An architectural understanding of natural sway frequencies in trees. J. R. Soc. Interface 2019, 16, 20190116. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Défossez, P.; Dupont, S. A root-to-foliage tree dynamic model for gusty winds during windstorm conditions. Agric. For. Meteorol. 2020, 287, 107949. [Google Scholar] [CrossRef]
- Schelhaas, M.J.; Nabuurs, G.J.; Schuck, A. Natural disturbances in the European forests in the 19th and 20th centuries. Glob. Chang. Biol. 2003, 9, 1620–1633. [Google Scholar] [CrossRef]
- Gregow, H.; Laaksonen, A.; Alper, M.E. Increasing large scale windstorm damage in Western, Central and Northern European forests, 1951–2010. Sci. Rep. 2017, 7, 46397. [Google Scholar] [CrossRef]
- Hanewinkel, M.; Cullmann, D.A.; Schelhaas, M.J.; Nabuurs, G.J.; Zimmermann, N.E. Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Chang. 2013, 3, 203–207. [Google Scholar] [CrossRef]
- Nabuurs, G.J.; Verkerk, P.J.; Schelhaas, M.J.; Ramón González Olabarria, J.; Trasobares, A.; Cienciala, E. Climate-Smart Forestry: Mitigation impacts in three European regions. In From Science to Policy 6; European Forest Institute: Joensuu, Finland, 2018; p. 32. [Google Scholar]
- Johnstone, J.F.; McIntire, E.J.B.; Pedersen, E.J.; King, G.; Pisaric, M.J.F. A sensitive slope: Estimating landscape patterns of forest resilience in a changing climate. Ecosphere 2010, 1. [Google Scholar] [CrossRef]
- Yousefpour, R.; Hanewinkel, M.; Le Moguédec, G. Evaluating the suitability of management strategies of pure Norway spruce forests in the Black Forest area of Southwest Germany for adaptation to or mitigation of climate change. Environ. Manag. 2010, 45, 387–402. [Google Scholar] [CrossRef] [PubMed]
- Nicoll, B.C.; Ray, D. Adaptive growth of tree root systems in response to wind action and site conditions. Tree Physiol. 1996, 16, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Nicoll, B.C.; Gardiner, B.A.; Rayner, B.; Peace, A.J. Anchorage of coniferous trees in relation to species, soil type, and rooting depth. Can. J. For. Res. 2011, 36, 1871–1883. [Google Scholar] [CrossRef]
- Lindner, M.; Fitzgerald, J.B.; Zimmermann, N.E.; Reyer, C.; Delzon, S.; van der Maaten, E.; Schelhaas, M.J.; Lasch, P.; Eggers, J.; van der Maaten-Theunissen, M.; et al. Climate change and European forests: What do we know, what are the uncertainties, and what are the implications for forest management? J. Environ. Manag. 2014, 146, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Suvanto, S.; Peltoniemi, M.; Tuominen, S.; Strandström, M.; Lehtonen, A. High-resolution mapping of forest vulnerability to wind for disturbance-aware forestry. For. Ecol. Manag. 2019, 453, 117619. [Google Scholar] [CrossRef]
- Martín, J.A.; Esteban, L.G.; de Palacios, P.; Fernández, F.G. Variation in wood anatomical traits of Pinus sylvestris L. between Spanish regions of provenance. Trees Struct. Funct. 2010, 24, 1017–1028. [Google Scholar] [CrossRef]
- Donat, M.G.; Renggli, D.; Wild, S.; Alexander, L.V.; Leckebusch, G.C.; Ulbrich, U. Reanalysis suggests long-term upward trends in European storminess since 1871. Geophys. Res. Lett. 2011, 38. [Google Scholar] [CrossRef]
- Reyer, C.P.O.; Bathgate, S.; Blennow, K.; Borges, J.G.; Bugmann, H.; Delzon, S.; Faias, S.P.; Garcia-Gonzalo, J.; Gardiner, B.; Gonzalez-Olabarria, J.R.; et al. Are forest disturbances amplifying or canceling out climate change-induced productivity changes in European forests? Environ. Res. Lett. 2017, 12, 034027. [Google Scholar] [CrossRef]
- Peltola, H.; Ikonen, V.P.; Gregow, H.; Strandman, H.; Kilpeläinen, A.; Venäläinen, A.; Kellomäki, S. Impacts of climate change on timber production and regional risks of wind-induced damage to forests in Finland. For. Ecol. Manag. 2010, 260, 833–845. [Google Scholar] [CrossRef]
- ALRahahleh, L.; Kilpeläinen, A.; Ikonen, V.P.; Strandman, H.; Asikainen, A.; Venäläinen, A.; Kaurola, J.; Kangas, J.; Peltola, H. Effects of using certain tree species in forest regeneration on volume growth, timber yield, and carbon stock of boreal forests in Finland under different CMIP5 projections. Eur. J. For. Res. 2018, 137, 573–591. [Google Scholar] [CrossRef]
- Gardiner, B.; Schuck, A.R.T.; Schelhaas, M.J.; Orazio, C.; Blennow, K.; Nicoll, B. Living with Storm Damage to Forests What Science Can Tell Us; European Forest Institute: Joensuu, Finland, 2013; Volume 131, ISBN 978-952-5980-09-7. [Google Scholar]
- Díaz-Yáñez, O.; Mola-Yudego, B.; González-Olabarria, J.R.; Pukkala, T. How does forest composition and structure affect the stability against wind and snow? For. Ecol. Manag. 2017, 401, 215–222. [Google Scholar] [CrossRef]
- Suvanto, S.; Henttonen, H.M.; Nöjd, P.; Mäkinen, H. Forest susceptibility to storm damage is affected by similar factors regardless of storm type: Comparison of thunder storms and autumn extra-tropical cyclones in Finland. For. Ecol. Manag. 2016, 381, 17–28. [Google Scholar] [CrossRef]
- Donis, J.; Kitenberga, M.; Snepsts, G.; Dubrovskis, E.; Jansons, A. Factors affecting windstorm damage at the stand level in hemiboreal forests in Latvia: Case study of 2005 winter storm. Silva Fenn. 2018, 52, 10009. [Google Scholar] [CrossRef]
- Snepsts, G.; Kitenberga, M.; Elferts, D.; Donis, J.; Jansons, A. Stem damage modifies the impact of wind on Norway Spruces. Forests 2020, 11, 463. [Google Scholar] [CrossRef]
- Kellomäki, S.; Maajärvi, M.; Strandman, H.; Kilpeläinen, A.; Peltola, H. Model computations on the climate change effects on snow cover, soil moisture and soil frost in the boreal conditions over Finland. Silva Fenn. 2010, 44, 213–233. [Google Scholar] [CrossRef]
- Hynynen, J.; Niemistö, P.; Viherä-Aarnio, A.; Brunner, A.; Hein, S.; Velling, P. Silviculture of birch (Betula pendula Roth and Betula pubescens Ehrh.) in Northern Europe. Forestry 2010, 83, 103–119. [Google Scholar] [CrossRef]
- Rosenvald, K.; Ostonen, I.; Truu, M.; Truu, J.; Uri, V.; Vares, A.; Lõhmus, K. Fine-root rhizosphere and morphological adaptations to site conditions in interaction with tree mineral nutrition in young silver birch (Betula pendula Roth.) stands. Eur. J. For. Res. 2011, 130, 1055–1066. [Google Scholar] [CrossRef]
- Tenkanen, A.; Keski-Saari, S.; Salojärvi, J.; Oksanen, E.; Keinänen, M.; Kontunen-Soppela, S. Differences in growth and gas exchange between southern and northern provenances of silver birch (Betula pendula Roth) in northern Europe. Tree Physiol. 2020, 40, 198–214. [Google Scholar] [CrossRef]
- Jansons, Ā.; Matisons, R.; Šēnhofa, S.; Katrevičs, J.; Jansons, J. High-frequency variation of tree-ring width of some native and alien tree species in Latvia during the period 1965–2009. Dendrochronologia 2016, 40, 151–158. [Google Scholar] [CrossRef]
- Peltola, H.; Kellomäki, S.; Hassinen, A.; Granander, M. Mechanical stability of Scots pine, Norway spruce and birch: An analysis of tree-pulling experiments in Finland. For. Ecol. Manag. 2000, 135, 143–153. [Google Scholar] [CrossRef]
- Tsuda, Y.; Semerikov, V.; Sebastiani, F.; Vendramin, G.G.; Lascoux, M. Multispecies genetic structure and hybridization in the Betula genus across Eurasia. Mol. Ecol. 2017, 26, 589–605. [Google Scholar] [CrossRef] [PubMed]
- Stubbs, C.J.; Cook, D.D.; Niklas, K.J. A general review of the biomechanics of root anchorage. J. Exp. Bot. 2019, 70, 3439–3451. [Google Scholar] [CrossRef] [PubMed]
- Ahti, T.; Hämet-Ahti, L.; Jalas, J. Vegetation zones and their sections in northwestern Europe. Ann. Bot. Fenn. 1968, 5, 169–211. [Google Scholar]
- Rendenieks, Z.; Nikodemus, O.; Brumelis, G. The implications of stand composition, age and spatial patterns of forest regions with different ownership type for management optimisation in northern Latvia. For. Ecol. Manag. 2015, 335, 216–224. [Google Scholar] [CrossRef]
- Avotniece, Z.; Klavins, M.; Rodinovs, V. Changes of extreme climate events in Latvia. Environ. Clim. Technol. 2012, 9, 4–11. [Google Scholar] [CrossRef]
- Jaagus, J.; Briede, A.; Rimkus, E.; Sepp, M. Changes in precipitation regime in the Baltic countries in 1966–2015. Theor. Appl. Climatol. 2018, 131, 433–443. [Google Scholar] [CrossRef]
- Jaagus, J.; Briede, A.; Rimkus, E.; Remm, K. Precipitation pattern in the Baltic countries under the influence of large-scale atmospheric circulation and local landscape factors. Int. J. Climatol. 2009, 30. [Google Scholar] [CrossRef]
- Draveniece, A. Okeāniskās un kontinentālās gaisa masas Latvijā. Latv. Veģetācija 2007, 14, 135. [Google Scholar]
- Schoen, J.M.; Ashley, W.S. A climatology of fatal convective wind events by storm type. Weather Forecast. 2011, 26, 109–121. [Google Scholar] [CrossRef]
- Rädler, A.T.; Groenemeijer, P.H.; Faust, E.; Sausen, R.; Púčik, T. Frequency of severe thunderstorms across Europe expected to increase in the 21st century due to rising instability. NPJ Clim. Atmos. Sci. 2019, 2. [Google Scholar] [CrossRef]
- Bušs, K. Latvijas PSR Meža Tipoloģijas Pamati; LRZTIPI: Riga, Latvia, 1976. [Google Scholar]
- Krisans, O.; Matisons, R.; Rust, S.; Burnevica, N.; Bruna, L.; Elferts, D.; Kalvane, L.; Jansons, A. Presence of root rot reduces stability of Norway spruce (Picea abies): Results of static pulling tests in Latvia. Forests 2020, 11, 416. [Google Scholar] [CrossRef]
- Krisans, O.; Saleniece, R.; Rust, S.; Elferts, D.; Kapostins, R.; Jansons, A.; Matisons, R. Effect of bark-stripping on mechanical stability of Norway Spruce. Forests 2020, 11, 357. [Google Scholar] [CrossRef]
- Krišāns, O.; Samariks, V.; Donis, J.; Jansons, Ā. Structural Root-Plate Characteristics of Wind-Thrown Norway Spruce in Hemiboreal Forests of Latvia. Forests 2020, 11, 1143. [Google Scholar] [CrossRef]
- Detter, A.; Richter, K.; Rust, C.; Rust, S. Aktuelle Untersuchungen zum Primärversagen von grünem Holz—Current studies on primary failure in green wood. In Proceedings of the Jahrbuch der Baumpflege, Augsburg, Germany, 6–9 September 2015; pp. 156–167. [Google Scholar]
- Detter, A.; Rust, S.; Rust, C.; Maybaum, G. Determining strength limits for standing tree stems from bending tests. In Proceedings of the 18th International Nondestructive Testing and Evaluation of Wood Symposium, Madison, WI, USA, 24–27 September 2013; Ross, R.J., Wang, X., Eds.; U.S. Department of Agriculture, ForestService, Forest Products Laboratory: Madison, WI, USA, 2013; p. 226. [Google Scholar]
- Detter, A.; van Wassenaer, P.; Rust, S. Stability recovery in London Plane trees 8 years after primary anchorage failure | Request PDF. Arboric. Urban. For. 2019, 45, 279–288. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: http://www.r-project.org/ (accessed on 5 December 2019).
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Bartoń, K. MuMIn: Multi-Model Inference. 2016. R package version 1.43.17. Available online: https://CRAN.R-project.org/package=MuMIn. (accessed on 24 November 2020).
- Pallardy, S.G. Physiology Of Woody Plants; Academic Press: Cambridge, MA, USA, 2007. [Google Scholar]
- Bonnesoeur, V.; Constant, T.; Moulia, B.; Fournier, M. Forest trees filter chronic wind-signals to acclimate to high winds. New Phytol. 2016, 210, 850–860. [Google Scholar] [CrossRef]
- Mayr, S.; Bertel, C.; Dämon, B.; Beikircher, B. Static and dynamic bending has minor effects on xylem hydraulics of conifer branches (Picea abies, Pinus sylvestris). Plant. Cell Environ. 2014, 37, 2151–2157. [Google Scholar] [CrossRef]
- Stenlid, J.; Redfern, D.B. Spread within the Tree and Stand. In Heterobasidion Annosum. Biology, Ecology, Impact and Controltle; Woodward, S., Stenlid, J., Karjalainen, R., Hüttemann, A., Eds.; CAB International: Wallingford, CT, USA, 1998; pp. 125–143. [Google Scholar]


{kind=link}
| Site No. | Locality | Composition (%) | N | DBH (cm) | H (m) | Vstem (m3) | Root Depth (m) | GWCsoil (%) | Soil Density (kg m−3) | Soil Type |
|---|---|---|---|---|---|---|---|---|---|---|
| Dry | 39 | 22.5 ± 4.4 | 24.3 ± 3.5 | 0.47 ± 0.25 | 0.74 ± 0.19 | 16.9 ± 8.4 | 1352 ± 98 | |||
| 1 | Auce | B(100) | 5 | 21.4 ± 3.7 | 21.2 ± 1.7 | 0.36 ± 0.14 | 0.72 ± 0.10 | 18.2 ± 0.3 | 1467 ± 70 | Loamy |
| 2 | Auce | B(70), A(20), G(10) | 4 | 25.9 ± 0.6 | 24.5 ± 1.7 | 0.58 ± 0.07 | 0.79 ± 0.40 | 26.6 ± 4.9 | 1345 ± 124 | Loamy |
| 3 | Kalsnava | B(100) | 5 | 18.7 ± 3.5 | 21.6 ± 1.9 | 0.28 ± 0.11 | 0.64 ± 0.10 | 7.8 ± 1.2 | 1357 ± 52 | Loamy |
| 4 | Kalsnava | B(100) | 5 | 21.1 ± 3.9 | 24.4 ± 1.1 | 0.40 ± 0.17 | 0.84 ± 0.24 | 6.5 ± 0.7 | 1446 ± 133 | Loamy |
| 5 | Kalsnava | B(70), O(30) | 5 | 20.6 ± 1.7 | 22.5 ± 1.3 | 0.34 ± 0.07 | 0.70 ± 0.12 | 11.6 ± 1.5 | 1288 ± 116 | Loamy |
| 6 | Kalsnava | B(90), S(10) | 5 | 21.3 ± 2.8 | 22.0 ± 2.6 | 0.37 ± 0.14 | 0.64 ± 0.11 | 13.6 ± 0.9 | 1306 ± 110 | Loamy podzolic |
| 7 | Skede | B(80), G 20) | 10 | 25.7 ± 5.6 | 29.2 ± 1.4 | 0.71 ± 0.32 | 0.79 ± 0.32 | 26.3 ± 4.3 | 1256 ± 79 | Silty |
| Periodically waterlogged | 32 | 19.8 ± 3.6 | 21.7 ± 2.4 | 0.32 ± 0.15 | 0.62 ± 0.20 | 38.9 ± 19.3 | 1268 ± 108 | |||
| 8 | Auce | B(70), S(30) | 6 | 25.3 ± 2.6 | 25.1 ± 2.2 | 0.57 ± 0.14 | 0.85 ± 0.06 | 24.6 ± 0.7 | 1348 ± 132 | Loamy glayed |
| 9 | Kalsnava | B(70), G(30) | 3 | 19.9 ± 4.4 | 18.5 ± 1.7 | 0.28 ± 0.15 | 0.67 ± 0.12 | 30.3 ± 0.6 | 1201 ± 67 | Loamy glayed |
| 10 | Kalsnava | B 100) | 4 | 19.0 ± 1.9 | 20.7 ± 0.7 | 0.27 ± 0.06 | 0.45 ± 0.21 | 83.7 ± 0.9 | 1276 ± 192 | Loamy glayed |
| 11 | Skede | B(90), G(10) | 9 | 18.7 ± 2.2 | 21.3 ± 1.2 | 0.27 ± 0.06 | 0.54 ± 0.22 | 36.4 ± 11.2 | 1269 ± 68 | Silty glayed |
| 12 | Skede | B(80), G(20) | 10 | 17.7 ± 2.3 | 21.2 ± 1.5 | 0.24 ± 0.07 | 0.48 ± 0.10 | 34.3 ± 9.5 | 1248 ± 84 | Silty glayed |
| Site No. | BBMPF (kNm) | BBMSF (kNm) | BBMdif (kNm) | Stem Breakage, N |
|---|---|---|---|---|
| Dry | 27.36 ± 12.35 | 37.04 ± 18.36 | 9.68 ± 8.92 | 2 |
| 1 | 22.06 ± 7.39 | 28.14 ± 8.59 | 6.08 ± 2.07 | 0 |
| 2 | 30.50 ± 6.65 | 43.57 ± 8.84 | 13.07 ± 3.86 | 0 |
| 3 | 20.75 ± 7.78 | 24.18 ± 8.40 | 3.43 ± 1.63 | 1 |
| 4 | 29.90 ± 14.69 | 34.30 ± 13.56 | 4.40 ± 1.36 | 0 |
| 5 | 22.19 ± 7.00 | 25.65 ± 7.91 | 3.47 ± 2.04 | 0 |
| 6 | 24.39 ± 13.10 | 29.78 ± 14.12 | 5.38 ± 2.69 | 0 |
| 7 | 34.84 ± 15.95 | 55.99 ± 21.79 | 21.15 ± 9.72 | 1 |
| Periodically waterlogged | 20.56 ± 9.66 | 27.67 ± 10.13 | 7.11 ± 4.05 | 7 |
| 8 | 34.06 ± 11.09 | 37.11 ± 12.33 | 3.06 ± 1.42 | 0 |
| 9 | 18.41 ± 6.01 | 21.77 ± 5.67 | 3.37 ± 1.23 | 2 |
| 10 | 19.39 ± 6.06 | 29.31 ± 10.97 | 9.92 ± 5.96 | 1 |
| 11 | 16.77 ± 4.46 | 26.20 ± 5.30 | 9.43 ± 2.15 | 4 |
| 12 | 16.99 ± 7.92 | 24.45 ± 10.52 | 7.46 ± 3.81 | 0 |
| BBMPF | BBMSF | BBMdif | Root depth | |||||
|---|---|---|---|---|---|---|---|---|
| Random effects | Variance | SD | Variance | SD | Variance | SD | Variance | SD |
| Site | 2.61 | 0.98 | 2.88 | 0.71 | 7.12 | 2.67 | 0.31 | 0.02 |
| Residual | 35.01 | 5.92 | 43.79 | 6.61 | 12.22 | 3.49 | 24.11 | 0.15 |
| Fixed effects | χ2-value | p-value | χ2-value | p-value | χ2-value | p-value | χ2-value | p-value |
| Vstem (covariate) | 134.64 | <0.001 | 285.13 | <0.001 | 49.15 | <0.001 | 31.07 | <0.001 |
| Site type | 0.55 | 0.81 | 0.03 | 0.869 | 0.06 | 0.798 | 2.02 | 0.154 |
| Vstem by site type | 5.04 | 0.025 | 4.89 | 0.041 | 4.55 | 0.042 | 3.19 | 0.07 |
| Marginal R2 | 0.69 | 0.82 | 0.56 | 0.44 | ||||
| Model p-value | <0.001 | <0.001 | <0.001 | <0.001 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krišāns, O.; Matisons, R.; Kitenberga, M.; Donis, J.; Rust, S.; Elferts, D.; Jansons, Ā. Wind Resistance of Eastern Baltic Silver Birch (Betula pendula Roth.) Suggests Its Suitability for Periodically Waterlogged Sites. Forests 2021, 12, 21. https://doi.org/10.3390/f12010021
Krišāns O, Matisons R, Kitenberga M, Donis J, Rust S, Elferts D, Jansons Ā. Wind Resistance of Eastern Baltic Silver Birch (Betula pendula Roth.) Suggests Its Suitability for Periodically Waterlogged Sites. Forests. 2021; 12(1):21. https://doi.org/10.3390/f12010021
Chicago/Turabian StyleKrišāns, Oskars, Roberts Matisons, Māra Kitenberga, Jānis Donis, Steffen Rust, Didzis Elferts, and Āris Jansons. 2021. "Wind Resistance of Eastern Baltic Silver Birch (Betula pendula Roth.) Suggests Its Suitability for Periodically Waterlogged Sites" Forests 12, no. 1: 21. https://doi.org/10.3390/f12010021
APA StyleKrišāns, O., Matisons, R., Kitenberga, M., Donis, J., Rust, S., Elferts, D., & Jansons, Ā. (2021). Wind Resistance of Eastern Baltic Silver Birch (Betula pendula Roth.) Suggests Its Suitability for Periodically Waterlogged Sites. Forests, 12(1), 21. https://doi.org/10.3390/f12010021