Potential Impact of Climate Change on the Forest Coverage and the Spatial Distribution of 19 Key Forest Tree Species in Italy under RCP4.5 IPCC Trajectory for 2050s


 , , , , , and
, , , , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Spatial Data and Climatic Scenarios
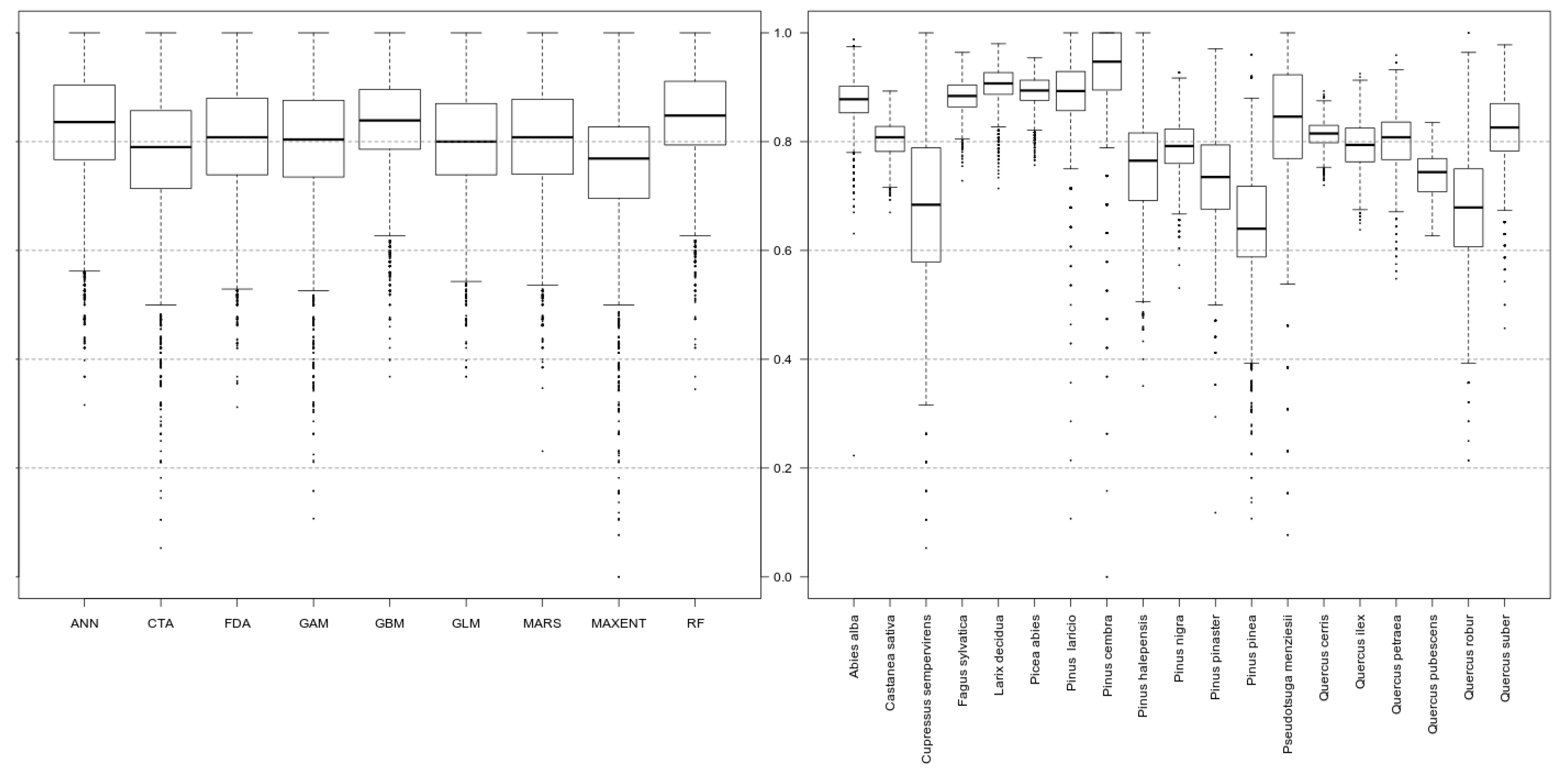
2.2. Species Distribution Ensemble Modelling
2.3. Suitability Maps Analysis and Uncertainties Quantification
3. Results
4. Discussion
4.1. Species-Specific Requirements against a Changing Climate
4.2. SDM as a Tool for Forest Management Options in the Italian Framework
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Availability of Data and Materials
References
- Deal, R.L.; Smith, N.; Gates, J. Ecosystem services to enhance sustainable forest management in the US: Moving from forest service national programmes to local projects in the Pacific Northwest. Forestry 2017, 90, 632–639. [Google Scholar] [CrossRef]
- Ray, D.; Petr, M.; Mullett, M.; Bathgate, S.; Marchi, M.; Beauchamp, K. A simulation-based approach to assess forest policy options under biotic and abiotic climate change impacts: A case study on Scotland’s National Forest Estate. For. Policy Econ. 2019, 103, 17–27. [Google Scholar] [CrossRef]
- Benito Garzón, M.; Robson, T.M.; Hampe, A. ΔTraitSDM: Species distribution models that account for local adaptation and phenotypic plasticity. New Phytol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Fréjaville, T.; Fady, B.; Kremer, A.; Ducousso, A.; Benito Garzón, M. Inferring phenotypic plasticity and local adaptation to climate across tree species ranges using forest inventory data. Glob. Ecol. Biogeogr. 2019, 28, 1–34. [Google Scholar] [CrossRef]
- Di Biase, R.M.; Fattorini, L.; Marchi, M. Statistical inferential techniques for approaching forest mapping. A review of methods. Ann. Silvic. Res. 2018, 42, 46–58. [Google Scholar] [CrossRef]
- Broome, A.; Bellamy, C.; Rattey, A.; Ray, D.; Quine, C.P.; Park, K.J. Niches for Species, a multi-species model to guide woodland management: An example based on Scotland’s native woodlands. Ecol. Indic. 2019, 103, 410–424. [Google Scholar] [CrossRef]
- Falk, W.; Mellert, K.H. Species distribution models as a tool for forest management planning under climate change: Risk evaluation of Abies alba in Bavaria. J. Veg. Sci. 2011, 22, 621–634. [Google Scholar] [CrossRef]
- Lenoir, J.; Gégout, J.C.; Marquet, P.A.; De Ruffray, P.; Brisse, H. A significant upward shift in plant species optimum elevation during the 20th century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef]
- O’Neill, G.A.; Hamann, A.; Wang, T.L. Accounting for population variation improves estimates of the impact of climate change on species’ growth and distribution. J. Appl. Ecol. 2008, 45, 1040–1049. [Google Scholar] [CrossRef]
- Williams, M.I.; Dumroese, R.K. Preparing for Climate Change: Forestry and Assisted Migration. J. For. 2013, 111, 287–297. [Google Scholar] [CrossRef]
- Cudlín, P.; Klopčič, M.; Tognetti, R.; Mališ, F.; Alados, C.L.; Bebi, P.; Grunewald, K.; Zhiyanski, M.; Andonowski, V.; Porta, N.L.; et al. Drivers of treeline shift in different European mountains. Clim. Res. 2017, 73, 135–150. [Google Scholar] [CrossRef]
- Chen, I.C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef] [PubMed]
- Marchi, M.; Nocentini, S.; Ducci, F. Future scenarios and conservation strategies for a rear-edge marginal population of Pinus nigra Arnold in Italian central Apennines. For. Syst. 2016, 25, e072. [Google Scholar] [CrossRef]
- Boisvert-Marsh, L.; Périé, C.; De Blois, S. Shifting with climate? Evidence for recent changes in tree species distribution at high latitudes. Ecosphere 2014, 5, 1–33. [Google Scholar] [CrossRef]
- Monleon, V.J.; Lintz, H.E. Evidence of tree species’ range shifts in a complex landscape. PLoS ONE 2015, 10, e0118069. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, K.L. What is close-to-nature silviculture in a changing world? Forestry 2016, 89, 1–6. [Google Scholar] [CrossRef]
- Puettmann, K.J.; Wilson, S.M.G.; Baker, S.C.; Donoso, P.J.; Drössler, L.; Amente, G.; Harvey, B.D.; Knoke, T.; Lu, Y.; Nocentini, S.; et al. Silvicultural alternatives to conventional even-aged forest management—What limits global adoption? For. Ecosyst. 2015, 2, 8. [Google Scholar] [CrossRef]
- Nocentini, S.; Buttoud, G.; Ciancio, O.; Corona, P. Managing forests in a changing world: The need for a systemic approach. A review. For. Syst. 2017, 26, 1. [Google Scholar] [CrossRef]
- Ruddell, S.; Sampson, R.; Smith, M.; Giffen, R.; Cathcart, J.; Hagan, J.; Sosland, D.; Godbee, J.; Heissenbuttel, J.; Lovett, S.; et al. The role for sustainably managed forests in climate change mitigation. J. For. 2007, 105, 314–319. [Google Scholar]
- MacDicken, K.G.; Sola, P.; Hall, J.E.; Sabogal, C.; Tadoum, M.; de Wasseige, C. Global progress toward sustainable forest management. For. Ecol. Manag. 2015, 352, 47–56. [Google Scholar] [CrossRef]
- Luyssaert, S.; Marie, G.; Valade, A.; Chen, Y.Y.; Njakou Djomo, S.; Ryder, J.; Otto, J.; Naudts, K.; Lansø, A.S.; Ghattas, J.; et al. Trade-offs in using European forests to meet climate objectives. Nature 2018, 562, 259–262. [Google Scholar] [CrossRef] [PubMed]
- Bellassen, V.; Luyssaert, S. Carbon sequestration: Managing forests in uncertain times. Nature 2014, 506, 153–155. [Google Scholar] [CrossRef] [PubMed]
- Fady, B.; Aravanopoulos, F.A.; Alizoti, P.; Mátyás, C.; von Wühlisch, G.; Westergren, M.; Belletti, P.; Cvjetkovic, B.; Ducci, F.; Huber, G.; et al. Evolution-based approach needed for the conservation and silviculture of peripheral forest tree populations. For. Ecol. Manag. 2016, 375, 66–75. [Google Scholar] [CrossRef]
- Roces-Díaz, J.V.; Jiménez-Alfaro, B.; Álvarez-Álvarez, P.; Álvarez-García, M.A. Environmental niche and distribution of six deciduous tree species in the spanish atlantic region. iFor. Biogeosci. For. 2014, 8, 214–221. [Google Scholar] [CrossRef]
- Pecchi, M.; Marchi, M.; Giannetti, F.; Bernetti, I.; Bindi, M.; Moriondo, M.; Maselli, F.; Fibbi, L.; Corona, P.; Travaglini, D.; et al. Reviewing climatic traits for the main forest tree species in Italy. iFor. Biogeosci. For. 2019, 12, 173–180. [Google Scholar] [CrossRef]
- Olthoff, A.; Martínez-Ruiz, C.; Alday, J.G. Distribution patterns of forest species along an Atlantic-Mediterranean environmental gradient: An approach from forest inventory data. Forestry 2016, 89, 46–54. [Google Scholar] [CrossRef]
- Marchi, M.; Ducci, F. Some refinements on species distribution models using tree-level national forest inventories for supporting forest management and marginal forest population detection. iFor. Biogeosci. For. 2018, 11, 291–299. [Google Scholar] [CrossRef]
- Hampe, A.; Petit, R.J. Conserving biodiversity under climate change: The rear edge matters. Ecol. Lett. 2005, 8, 461–467. [Google Scholar] [CrossRef]
- Zhang, J.; Nielsen, S.E.; Chen, Y.; Georges, D.; Qin, Y.; Wang, S.S.; Svenning, J.C.; Thuiller, W. Extinction risk of North American seed plants elevated by climate and land-use change. J. Appl. Ecol. 2017, 54, 303–312. [Google Scholar] [CrossRef]
- Journé, V.; Barnagaud, J.Y.; Bernard, C.; Crochet, P.A.; Morin, X. Correlative climatic niche models predict real and virtual species distributions equally well. Ecology 2019, 101, 1–14. [Google Scholar] [CrossRef]
- Pecchi, M.; Marchi, M.; Burton, V.; Giannetti, F.; Moriondo, M.; Bernetti, I.; Bindi, M.; Chirici, G. Species distribution modelling to support forest management. A literature review. Ecol. Model. 2019, 411, 108817. [Google Scholar] [CrossRef]
- Booth, T.H. Species distribution modelling tools and databases to assist managing forests under climate change. For. Ecol. Manag. 2018, 430, 196–203. [Google Scholar] [CrossRef]
- Iverson, L.R.; Prasad, A.M.; Hale, B.J.; Sutherland, E.K. Atlas of Current and Potential Future Distributions of Common Trees of the Eastern United States; US Department of Agriculture, Forest Service, Northeastern Research Station: Madison, WI, USA, 1999.
- Badeau, V.; Dupouey, J.; Cluzeau, C.; Drapier, J.; Badeau, V.; Dupouey, J.; Cluzeau, C.; Drapier, J.; Mod, C.L.B. Modélisation et Cartographie de l’ aire Climatique Potentielle des Grandes Essences Forestières. 2004. Available online: https://hal.inrae.fr/hal-02834220 (accessed on 3 April 2020).
- Tang, Y.; Winkler, J.A.; Viña, A.; Wang, F.; Zhang, J.; Zhao, Z.; Connor, T.; Yang, H.; Zhang, Y.; Zhang, X.; et al. Expanding ensembles of species present-day and future climatic suitability to consider the limitations of species occurrence data. Ecol. Indic. 2020, 110, 105891. [Google Scholar] [CrossRef]
- Beaumont, L.J.; Hughes, L.; Pitman, A.J. Why is the choice of future climate scenarios for species distribution modelling important? Ecol. Lett. 2008, 11, 1135–1146. [Google Scholar] [CrossRef]
- Jarnevich, C.S.; Young, N.E. Not so normal normals: Species distribution model results are sensitive to choice of climate normals and model type. Climate 2019, 7, 37. [Google Scholar] [CrossRef]
- Goberville, E.; Beaugrand, G.; Hautekèete, N.C.; Piquot, Y.; Luczak, C. Uncertainties in the projection of species distributions related to general circulation models. Ecol. Evol. 2015, 5, 1100–1116. [Google Scholar] [CrossRef]
- Garbolino, E.; Sanseverino-Godfrin, V.; Hinojos-Mendoza, G. Describing and predicting of the vegetation development of Corsica due to expected climate change and its impact on forest fire risk evolution. Saf. Sci. 2015. [Google Scholar] [CrossRef]
- Kindt, R. Ensemble species distribution modelling with transformed suitability values. Environ. Model. Softw. 2018, 100, 136–145. [Google Scholar] [CrossRef]
- Crimmins, S.M.; Dobrowski, S.Z.; Mynsberge, A.R. Evaluating ensemble forecasts of plant species distributions under climate change. Ecol. Model. 2013, 266, 126–130. [Google Scholar] [CrossRef]
- Hao, T.; Elith, J.; Guillera-Arroita, G.; Lahoz-Monfort, J.J. A review of evidence about use and performance of species distribution modelling ensembles like BIOMOD. Divers. Distrib. 2019, 25, 839–852. [Google Scholar] [CrossRef]
- Lowe, J.A.; Bernie, D.; Bett, P.; Bricheno, L.; Brown, S.; Calvert, D.; Clark, R.; Eagle, K.; Edwards, T.; Fosser, G.; et al. UKCP18 Science Overview Report Version 2.0; Met Office: Exeter, UK, 2019.
- Moriondo, M.; Bindi, M. Comparison of temperatures simulated by GCMs, RCMs and statistical downscaling: Potential application in studies of future crop development. Clim. Res. 2006, 30, 149–160. [Google Scholar] [CrossRef][Green Version]
- Beale, C.M.; Lennon, J.J. Incorporating uncertainty in predictive species distribution modelling. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Moreno, A.; Hasenauer, H. Spatial downscaling of European climate data. Int. J. Climatol. 2016, 36, 1444–1458. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I.; Angulo, M.; El Kenawy, A. A New Global 0.5° Gridded Dataset (1901–2006) of a Multiscalar Drought Index: Comparison with Current Drought Index Datasets Based on the Palmer Drought Severity Index. J. Hydrometeorol. 2010, 11, 1033–1043. [Google Scholar] [CrossRef]
- Fréjaville, T.; Benito Garzón, M. The EuMedClim Database: Yearly Climate Data (1901–2014) of 1 km Resolution Grids for Europe and the Mediterranean Basin. Front. Ecol. Evol. 2018, 6, 1–5. [Google Scholar] [CrossRef]
- Harris, I.; Osborn, T.J.; Jones, P.; Lister, D. Version 4 of the CRU TS monthly high-resolution gridded multivariate climate dataset. Sci. Data 2020, 7, 109. [Google Scholar] [CrossRef]
- Wang, T.; Hamann, A.; Spittlehouse, D.; Carroll, C. Locally downscaled and spatially customizable climate data for historical and future periods for North America. PLoS ONE 2016, 11, e0156720. [Google Scholar] [CrossRef]
- Lin, H.Y.; Hu, J.M.; Chen, T.Y.; Hsieh, C.F.; Wang, G.; Wang, T. A dynamic downscaling approach to generate scale-free regional climate data in Taiwan. Taiwania 2018, 63, 251–266. [Google Scholar] [CrossRef]
- Isaac-Renton, M.G.; Roberts, D.R.; Hamann, A.; Spiecker, H. Douglas-fir plantations in Europe: A retrospective test of assisted migration to address climate change. Glob. Chang. Biol. 2014, 20, 2607–2617. [Google Scholar] [CrossRef]
- Noce, S.; Collalti, A.; Santini, M. Likelihood of changes in forest species suitability, distribution, and diversity under future climate: The case of Southern Europe. Ecol. Evol. 2017, 7, 9358–9375. [Google Scholar] [CrossRef]
- Attorre, F.; Alfo, M.; De Sanctis, M.; Francesconi, F.; Valenti, R.; Vitale, M.; Bruno, F. Evaluating the effects of climate change on tree species abundance and distribution in the Italian peninsula. Appl. Veg. Sci. 2011, 14, 242–255. [Google Scholar] [CrossRef]
- Fattorini, L. Design-based methodological advances to support national forest inventories: A review of recent proposals. iFor. Biogeosci. For. 2014, 83, 6. [Google Scholar] [CrossRef]
- Borghetti, M.; Chirici, G. Raw data from the Italian National Forest Inventory are on-line and publicly available. For. J. Silvic. For. Ecol. 2016, 13, 33–34. [Google Scholar] [CrossRef]
- Maselli, F.; Pasqui, M.; Chirici, G.; Chiesi, M.; Fibbi, L.; Salvati, R.; Corona, P. Modeling primary production using a 1 km daily meteorological data set. Clim. Res. 2012, 54, 271–285. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J. Dismo: Species Distribution Modeling, R package version 1.0-15; 2015. Available online: http://CRAN.R-project.org/package=dismo (accessed on 25 February 2020).
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Marchi, M.; Sinjur, I.; Bozzano, M.; Westergren, M. Evaluating WorldClim Version 1 (1961–1990) as the Baseline for Sustainable Use of Forest and Environmental Resources in a Changing Climate. Sustainability 2019, 11, 3043. [Google Scholar] [CrossRef]
- Bucchignani, E.; Mercogliano, P.; Panitz, H.J.; Montesarchio, M. Climate change projections for the Middle East–North Africa domain with COSMO-CLM at different spatial resolutions. Adv. Clim. Chang. Res. 2018, 9, 66–80. [Google Scholar] [CrossRef]
- Fibbi, L.; Moriondo, M.; Chiesi, M.; Bindi, M.; Maselli, F. Impacts of climate change on the gross primary production of Italian forests. Ann. For. Sci. 2019, 76. [Google Scholar] [CrossRef]
- Araújo, M.B.; New, M. Ensemble forecasting of species distributions. Trends Ecol. Evol. 2007, 22, 42–47. [Google Scholar] [CrossRef]
- Leroy, B.; Delsol, R.; Hugueny, B.; Meynard, C.N.; Barhoumi, C.; Barbet-Massin, M.; Bellard, C. Without quality presence–absence data, discrimination metrics such as TSS can be misleading measures of model performance. J. Biogeogr. 2018, 45, 1994–2002. [Google Scholar] [CrossRef]
- Marmion, M.; Parviainen, M.; Luoto, M.; Heikkinen, R.K.; Thuiller, W. Evaluation of consensus methods in predictive species distribution modelling. Divers. Distrib. 2009, 15, 59–69. [Google Scholar] [CrossRef]
- Thuiller, W.; Georges, D.; Engler, R. Biomod2: Ensemble Platform for Species Distribution Modeling; R Development Core Team: Vienna, Austria, 2020. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Development Core Team: Vienna, Austria, 2020. [Google Scholar]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography Cop. 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting pseudo-absences for species distribution models: How, where and how many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- Harris, R.M.B.; Grose, M.R.; Lee, G.; Bindoff, N.L.; Porfirio, L.L.; Fox-Hughes, P. Climate projections for ecologists. Wiley Interdiscip. Rev. Clim. Chang. 2014, 5, 621–637. [Google Scholar] [CrossRef]
- Keenan, R.J. Climate change impacts and adaptation in forest management: A review. Ann. For. Sci. 2015, 72, 145–167. [Google Scholar] [CrossRef]
- Sørland, S.L.; Schär, C.; Lüthi, D.; Kjellström, E. Bias patterns and climate change signals in GCM-RCM model chains. Environ. Res. Lett. 2018. [Google Scholar] [CrossRef]
- Buras, A.; Menzel, A. Projecting tree species composition changes of european forests for 2061–2090 under RCP 4.5 and RCP 8.5 scenarios. Front. Plant Sci. 2019, 9. [Google Scholar] [CrossRef]
- Lelieveld, J.; Hadjinicolaou, P.; Kostopoulou, E.; Chenoweth, J.; El Maayar, M.; Giannakopoulos, C.; Hannides, C.; Lange, M.A.; Tanarhte, M.; Tyrlis, E.; et al. Climate change and impacts in the Eastern Mediterranean and the Middle East. Clim. Chang. 2012, 114, 667–687. [Google Scholar] [CrossRef]
- Giannakopoulos, C.; Le Sager, P.; Bindi, M.; Moriondo, M.; Kostopoulou, E.; Goodess, C. Climatic changes and associated impacts in the Mediterranean resulting from a 2 °C global warming. Glob. Planet. Chang. 2009, 68, 209–224. [Google Scholar] [CrossRef]
- Liu, S.; Liang, X.; Gao, W.; Stohlgren, T.J. Regional climate model downscaling may improve the prediction of alien plant species distributions. Front. Earth Sci. 2014, 8, 457–471. [Google Scholar] [CrossRef]
- Wang, T.; Campbell, E.M.; O’Neill, G.A.; Aitken, S.N. Projecting future distributions of ecosystem climate niches: Uncertainties and management applications. For. Ecol. Manag. 2012, 279, 128–140. [Google Scholar] [CrossRef]
- Wang, T.; Wang, G.; Innes, J.; Nitschke, C.; Kang, H. Climatic niche models and their consensus projections for future climates for four major forest tree species in the Asia-Pacific region. For. Ecol. Manag. 2016, 360, 357–366. [Google Scholar] [CrossRef]
- Fourcade, Y.; Besnard, A.G.; Secondi, J. Paintings predict the distribution of species, or the challenge of selecting environmental predictors and evaluation statistics. Glob. Ecol. Biogeogr. 2018, 27, 245–256. [Google Scholar] [CrossRef]
- Gomes, V.H.F.; Ijff, S.D.; Raes, N.; Amaral, I.L.; Salomão, R.P.; Coelho, L.D.S.; Matos, F.D.D.A.; Castilho, C.V.; Filho, D.D.A.L.; López, D.C.; et al. Species Distribution Modelling: Contrasting presence-only models with plot abundance data. Sci. Rep. 2018, 8, 1–2. [Google Scholar] [CrossRef]
- Valladares, F.; Matesanz, S.; Guilhaumon, F.; Araújo, M.B.; Balaguer, L.; Benito-Garzón, M.; Cornwell, W.; Gianoli, E.; van Kleunen, M.; Naya, D.E.; et al. The effects of phenotypic plasticity and local adaptation on forecasts of species range shifts under climate change. Ecol. Lett. 2014, 17, 1351–1364. [Google Scholar] [CrossRef]
- Sáenz-Romero, C.; Lindig-Cisneros, R.A.; Joyce, D.G.; Beaulieu, J.; Bradley, J.S.C.; Jaquish, B.C. Assisted migration of forest populations for adapting trees to climate change. Revista Chapingo Serie Ciencias Forestales y del Ambiente 2016, 22, 303–323. [Google Scholar] [CrossRef]
- Peterson St-Laurent, G.; Hagerman, S.; Kozak, R. What risks matter? Public views about assisted migration and other climate-adaptive reforestation strategies. Clim. Chang. 2018, 151, 573–587. [Google Scholar] [CrossRef]
- Aubin, I.; Garbe, C.M.; Colombo, S.; Drever, C.R.; McKenney, D.W.; Messier, C.; Pedlar, J.; Saner, M.A.; Venier, L.; Wellstead, A.M.; et al. Why we disagree about assisted migration: Ethical implications of a key debate regarding the future of Canada’s forests. For. Chron. 2011, 87, 755–765. [Google Scholar] [CrossRef]
- Zhang, W.; Huang, D.; Wang, R.; Liu, J.; Du, N. Altitudinal patterns of species diversity and phylogenetic diversity across temperate mountain forests of northern China. PLoS ONE 2016, 11, e0159995. [Google Scholar] [CrossRef]
- Littell, J.S.; Peterson, D.L.; Tjoelker, M. Douglas-fir growth in mountain ecosystems: Water limits tree growth from stand to region. Ecol. Monogr. 2008, 78, 349–368. [Google Scholar] [CrossRef]
- Vacchiano, G.; Motta, R. An improved species distribution model for Scots pine and downy oak under future climate change in the NW Italian Alps. Ann. For. Sci. 2015, 72, 321–334. [Google Scholar] [CrossRef]
- Rumpf, S.B.; Hülber, K.; Klonner, G.; Moser, D.; Schütz, M.; Wessely, J.; Willner, W.; Zimmermann, N.E.; Dullinger, S. Range dynamics of mountain plants decrease with elevation. Proc. Natl. Acad. Sci. USA 2018, 115, 1848–1853. [Google Scholar] [CrossRef] [PubMed]
- Rumpf, S.B.; Hülber, K.; Zimmermann, N.E.; Dullinger, S. Elevational rear edges shifted at least as much as leading edges over the last century. Glob. Ecol. Biogeogr. 2019, 28, 533–543. [Google Scholar] [CrossRef]
- Rogora, M.; Frate, L.; Carranza, M.L.; Freppaz, M.; Stanisci, A.; Bertani, I.; Bottarin, R.; Brambilla, A.; Canullo, R.; Carbognani, M.; et al. Assessment of climate change effects on mountain ecosystems through a cross-site analysis in the Alps and Apennines. Sci. Total Environ. 2018, 624, 1429–1442. [Google Scholar] [CrossRef] [PubMed]
- Hanewinkel, M.; Cullmann, D.A.; Schelhaas, M.J.; Nabuurs, G.J.; Zimmermann, N.E. Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Chang. 2013, 3, 203–207. [Google Scholar] [CrossRef]
- Ruiz-Labourdette, D.; Nogués-Bravo, D.; Ollero, H.S.; Schmitz, M.F.; Pineda, F.D. Forest composition in Mediterranean mountains is projected to shift along the entire elevational gradient under climate change. J. Biogeogr. 2012, 39, 162–176. [Google Scholar] [CrossRef]
- Lindner, M.; Fitzgerald, J.B.; Zimmermann, N.E.; Reyer, C.; Delzon, S.; van der Maaten, E.; Schelhaas, M.J.; Lasch, P.; Eggers, J.; van der Maaten-Theunissen, M.; et al. Climate change and European forests: What do we know, what are the uncertainties, and what are the implications for forest management? J. Environ. Manag. 2014, 146, 69–83. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.T.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Abatzoglou, J.T.; Williams, A.P. Impact of anthropogenic climate change on wildfire across western US forests. Proc. Natl. Acad. Sci. USA 2016. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, C.; Salvati, L.; Corona, P.; Romano, R.; Marchi, M. The background context matters: Local-scale socioeconomic conditions and the spatial distribution of wildfires in Italy. Sci. Total Environ. 2019, 654, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Fargeon, H.; Pimont, F.; Martin-StPaul, N.; De Caceres, M.; Ruffault, J.; Barbero, R.; Dupuy, J.L. Projections of fire danger under climate change over France: Where do the greatest uncertainties lie? Clim. Chang. 2020. [Google Scholar] [CrossRef]
- Casalegno, S.; Amatulli, G.; Camia, A.; Nelson, A.; Pekkarinen, A. Vulnerability of Pinus cembra L. in the Alps and the Carpathian mountains under present and future climates. For. Ecol. Manag. 2010, 259, 750–761. [Google Scholar] [CrossRef]
- Castaldi, C.; Marchi, M.; Vacchiano, G.; Corona, P. Douglas-fir climate sensitivity at two contrasting sites along the southern limit of the European planting range. J. For. Res. 2019. [Google Scholar] [CrossRef]
- Vitali, V.; Büntgen, U.; Bauhus, J. Silver fir and Douglas fir are more tolerant to extreme droughts than Norway spruce in south-western Germany. Glob. Chang. Biol. 2017, 23, 5108–5119. [Google Scholar] [CrossRef] [PubMed]
- Dyderski, M.K.; Paź, S.; Frelich, L.E.; Jagodziński, A.M. How much does climate change threaten European forest tree species distributions? Glob. Chang. Biol. 2018, 24, 1150–1163. [Google Scholar] [CrossRef]
- Cheaib, A.; Badeau, V.; Boe, J.; Chuine, I.; Delire, C.; Dufrêne, E.; François, C.; Gritti, E.S.; Legay, M.; Pagé, C.; et al. Climate change impacts on tree ranges: Model intercomparison facilitates understanding and quantification of uncertainty. Ecol. Lett. 2012, 15, 533–544. [Google Scholar] [CrossRef]
- Perkins, D.; Uhl, E.; Biber, P.; du Toit, B.; Carraro, V.; Rötzer, T.; Pretzsch, H. Impact of climate trends and drought events on the growth of oaks (Quercus robur L. and Quercus petraea (Matt.) Liebl.) within and beyond their natural range. Forests 2018, 9, 108. [Google Scholar] [CrossRef]
- Kim, H.N.; Jin, H.Y.; Kwak, M.J.; Khaine, I.; You, H.N.; Lee, T.Y.; Ahn, T.H.; Woo, S.Y. Why does Quercus suber species decline in Mediterranean areas? J. Asia Pacific Biodivers. 2017, 10, 337–341. [Google Scholar] [CrossRef]
- Vitale, M.; Mancini, M.; Matteucci, G.; Francesconi, F.; Valenti, R.; Attorre, F. Model-based assessment of ecological adaptations of three forest tree species growing in Italy and impact on carbon and water balance at national scale under current and future climate scenarios. iFor. Biogeosci. For. 2012, 5, 235–246. [Google Scholar] [CrossRef]
- Märkel, U.; Dolos, K. Tree species site suitability as a combination of occurrence probability and growth and derivation of priority regions for climate change adaptation. Forests 2017, 8, 181. [Google Scholar] [CrossRef]
- Mamet, S.D.; Brown, C.D.; Trant, A.J.; Laroque, C.P. Shifting global Larix distributions: Northern expansion and southern retraction as species respond to changing climate. J. Biogeogr. 2019, 46, 30–44. [Google Scholar] [CrossRef]
- Vitasse, Y.; Bottero, A.; Rebetez, M.; Conedera, M.; Augustin, S.; Brang, P.; Tinner, W. What is the potential of silver fir to thrive under warmer and drier climate? Eur. J. For. Res. 2019, 138, 547–560. [Google Scholar] [CrossRef]
- Eilmann, B.; de Vries, S.M.G.; den Ouden, J.; Mohren, G.M.J.; Sauren, P.; Sass-Klaassen, U. Origin matters! Difference in drought tolerance and productivity of coastal Douglas-fir (Pseudotsuga menziesii (Mirb.)) provenances. For. Ecol. Manag. 2013, 302, 133–143. [Google Scholar] [CrossRef]
- Gray, L.K.; Rweyongeza, D.; Hamann, A.; John, S.; Thomas, B.R. Developing management strategies for tree improvement programs under climate change: Insights gained from long-term field trials with lodgepole pine. For. Ecol. Manag. 2016, 377, 128–138. [Google Scholar] [CrossRef]
- Klein, T.; Cahanovitc, R.; Sprintsin, M.; Herr, N.; Schiller, G. A nation-wide analysis of tree mortality under climate change: Forest loss and its causes in Israel 1948–2017. For. Ecol. Manag. 2019, 432, 840–849. [Google Scholar] [CrossRef]
- Freire, J.A.; Rodrigues, G.C.; Tomé, M. Climate change impacts on Pinus pinea L. Silvicultural System for cone production and ways to contour those impacts: A review complemented with data from permanent plots. Forests 2019, 10, 169. [Google Scholar] [CrossRef]
- Silvério, E.; Duque-Lazo, J.; Navarro-Cerrillo, R.M.; Pereña, F.; Palacios-Rodríguez, G. Resilience or Vulnerability of the Rear-Edge Distributions of Pinus halepensis and Pinus pinaster Plantations Versus that of Natural Populations, under Climate-Change Scenarios. For. Sci. 2019, 66, 178–190. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Predictor | Sum of Squares | Prop. of Explained Variance | df | F Value | Pr (>F) | |
|---|---|---|---|---|---|---|
| Climate scenario | 3 × 1010 | 0.20 | 7 | 6.2045 | 0.000137 | *** |
| Threshold | 1 × 1011 | 0.80 | 1 | 168.565 | 4.49 × 10−14 | *** |
| Climatic Scenario | LS Threshold | Number of Pixels | N. of Pixel Variation | Mean Elevation | Elevation SD | Minimum Elevation | Maximum Elevation |
|---|---|---|---|---|---|---|---|
| Current | 500 | 277,469 | - | 570.2 | 584.0 | 0 | 4322 |
| 600 | 270,012 | - | 565.3 | 561.8 | 0 | 3536 | |
| 700 | 259,095 | - | 565.9 | 544.1 | 0 | 3536 | |
| 800 | 241,857 | - | 563.5 | 522.7 | 0 | 3154 | |
| 900 | 202,913 | - | 580.5 | 496.9 | 0 | 2974 | |
| CC | 500 | 272,212 | −2% | 551.0 | 560.3 | 0 | 3786 |
| 600 | 253,718 | −6% | 551.8 | 538.2 | 0 | 3050 | |
| 700 | 232,846 | −10% | 557.3 | 524.3 | 0 | 3033 | |
| 800 | 191,699 | −21% | 572.0 | 514.0 | 0 | 3033 | |
| 900 | 117,636 | −42% | 580.1 | 498.2 | 0 | 2841 | |
| COSMO | 500 | 302,091 | +9% | 535.2 | 586.3 | 0 | 4783 |
| 600 | 161,849 | −40% | 794.8 | 622.7 | 0 | 4322 | |
| 700 | 117,167 | −55% | 911.1 | 622.1 | 0 | 3840 | |
| 800 | 83,045 | −66% | 1005.3 | 610.5 | 0 | 3536 | |
| 900 | 38,627 | −81% | 1122.9 | 560.5 | 2 | 3536 | |
| HD | 500 | 271,421 | −2% | 559.0 | 571.4 | 0 | 4322 |
| 600 | 249,366 | −8% | 556.8 | 537.2 | 0 | 3478 | |
| 700 | 227,487 | −12% | 562.3 | 523.2 | 0 | 3093 | |
| 800 | 186,541 | −23% | 570.0 | 498.9 | 0 | 3033 | |
| 900 | 107,279 | −47% | 526.5 | 444.1 | 0 | 2921 | |
| HE | 500 | 264,667 | −5% | 571.2 | 575.2 | 0 | 4412 |
| 600 | 243,331 | −10% | 567.0 | 541.5 | 0 | 3346 | |
| 700 | 220,858 | −15% | 574.7 | 530.2 | 0 | 3093 | |
| 800 | 175,290 | −28% | 588.2 | 511.6 | 0 | 3033 | |
| 900 | 97,472 | −52% | 561.2 | 468.0 | 0 | 2921 | |
| HG | 500 | 266,667 | −4% | 563.0 | 575.6 | 0 | 4783 |
| 600 | 248,089 | −8% | 553.3 | 538.4 | 0 | 3478 | |
| 700 | 225,522 | −13% | 557.4 | 523.1 | 0 | 3346 | |
| 800 | 183,055 | −24% | 555.1 | 504.9 | 0 | 3033 | |
| 900 | 111,688 | −45% | 513.8 | 441.4 | 0 | 2921 | |
| MG | 500 | 263,520 | −5% | 553.5 | 562.4 | 0 | 3786 |
| 600 | 245,959 | −9% | 548.4 | 536.0 | 0 | 3213 | |
| 700 | 225,935 | −13% | 541.2 | 514.4 | 0 | 3038 | |
| 800 | 183,215 | −24% | 553.1 | 503.0 | 0 | 2974 | |
| 900 | 103,091 | −49% | 549.1 | 495.1 | 0 | 2810 | |
| MP | 500 | 266,133 | −4% | 558.3 | 561.1 | 0 | 3840 |
| 600 | 245,089 | −9% | 558.2 | 534.0 | 0 | 3478 | |
| 700 | 222,756 | −14% | 563.2 | 520.9 | 0 | 3216 | |
| 800 | 170,854 | −29% | 588.6 | 516.9 | 0 | 2974 | |
| 900 | 90,737 | −55% | 579.7 | 493.0 | 0 | 2680 |
| Predictor | df | Sum of Squares | Prop. of Explained Variance | F Value | Pr (>F) | |
|---|---|---|---|---|---|---|
| Altitude | 1 | 1.72 × 107 | 0.45 | 1401.5 | <2.2 × 10−16 | *** |
| Longitude | 1 | 2.96 × 106 | 0.08 | 241.161 | <2.2 × 10−16 | *** |
| Latitude | 1 | 1.33 × 107 | 0.35 | 1086.71 | <2.2 × 10−16 | *** |
| Fortype | 18 | 4.21 × 106 | 0.11 | 19.0496 | <2.2 × 10−16 | *** |
| Gtot | 1 | 1.15 × 105 | 3.04 × 10−3 | 9.4081 | 0.00217 | ** |
| TypeFor | 3 | 1.58 × 105 | 4.15 × 10−3 | 4.2801 | 0.00502 | ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pecchi, M.; Marchi, M.; Moriondo, M.; Forzieri, G.; Ammoniaci, M.; Bernetti, I.; Bindi, M.; Chirici, G. Potential Impact of Climate Change on the Forest Coverage and the Spatial Distribution of 19 Key Forest Tree Species in Italy under RCP4.5 IPCC Trajectory for 2050s. Forests 2020, 11, 934. https://doi.org/10.3390/f11090934
Pecchi M, Marchi M, Moriondo M, Forzieri G, Ammoniaci M, Bernetti I, Bindi M, Chirici G. Potential Impact of Climate Change on the Forest Coverage and the Spatial Distribution of 19 Key Forest Tree Species in Italy under RCP4.5 IPCC Trajectory for 2050s. Forests. 2020; 11(9):934. https://doi.org/10.3390/f11090934
Chicago/Turabian StylePecchi, Matteo, Maurizio Marchi, Marco Moriondo, Giovanni Forzieri, Marco Ammoniaci, Iacopo Bernetti, Marco Bindi, and Gherardo Chirici. 2020. "Potential Impact of Climate Change on the Forest Coverage and the Spatial Distribution of 19 Key Forest Tree Species in Italy under RCP4.5 IPCC Trajectory for 2050s" Forests 11, no. 9: 934. https://doi.org/10.3390/f11090934
APA StylePecchi, M., Marchi, M., Moriondo, M., Forzieri, G., Ammoniaci, M., Bernetti, I., Bindi, M., & Chirici, G. (2020). Potential Impact of Climate Change on the Forest Coverage and the Spatial Distribution of 19 Key Forest Tree Species in Italy under RCP4.5 IPCC Trajectory for 2050s. Forests, 11(9), 934. https://doi.org/10.3390/f11090934