Predicting Suitable Habitats of Camptotheca acuminata Considering Both Climatic and Soil Variables



Abstract
1. Introduction
2. Materials and Methods
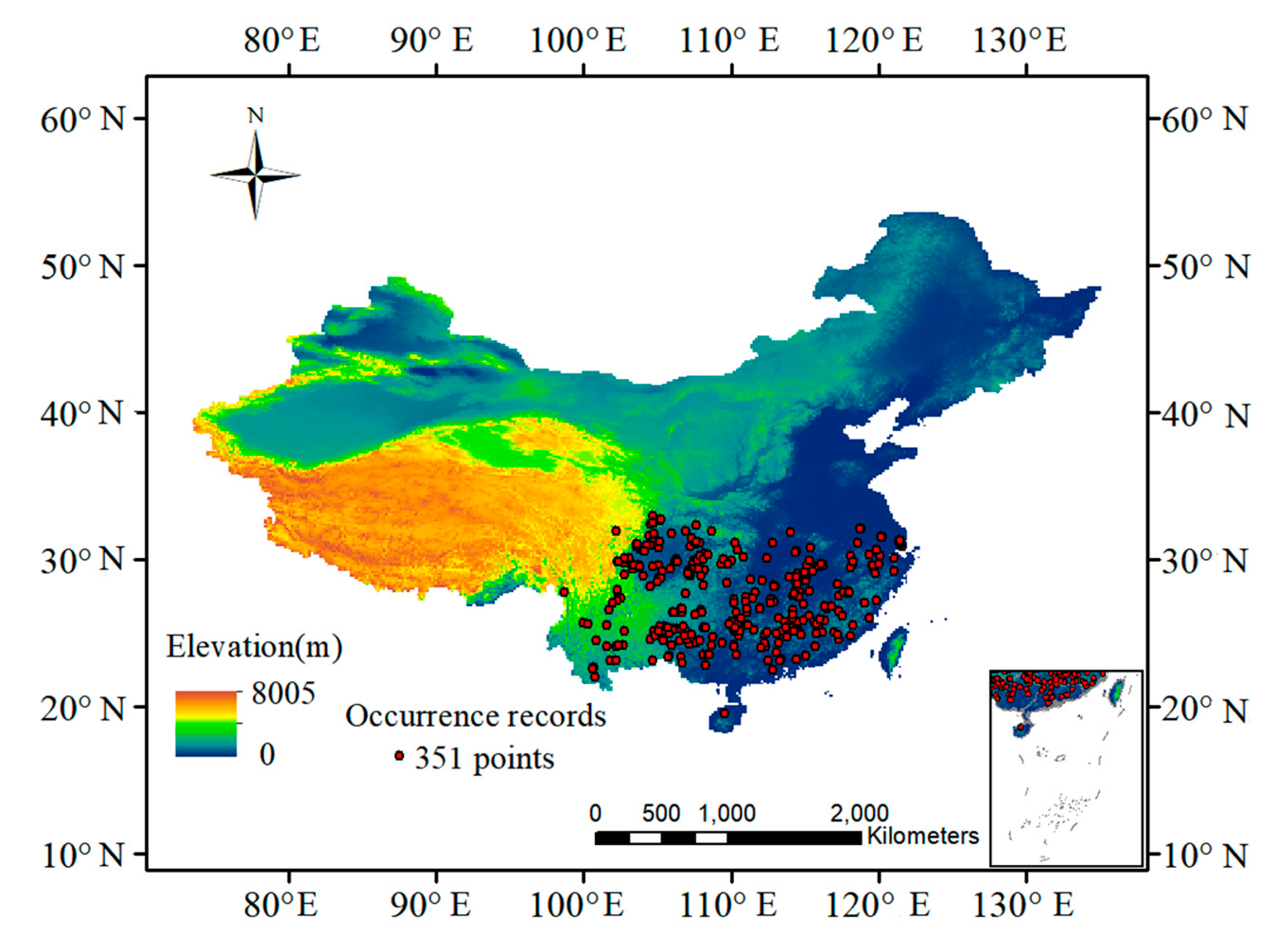
2.1. Species Occurrence Data
2.2. Environment Data
2.3. Model Development
2.4. Model Evaluation
2.5. Model Predictions and Assessments of Climate Change Impacts
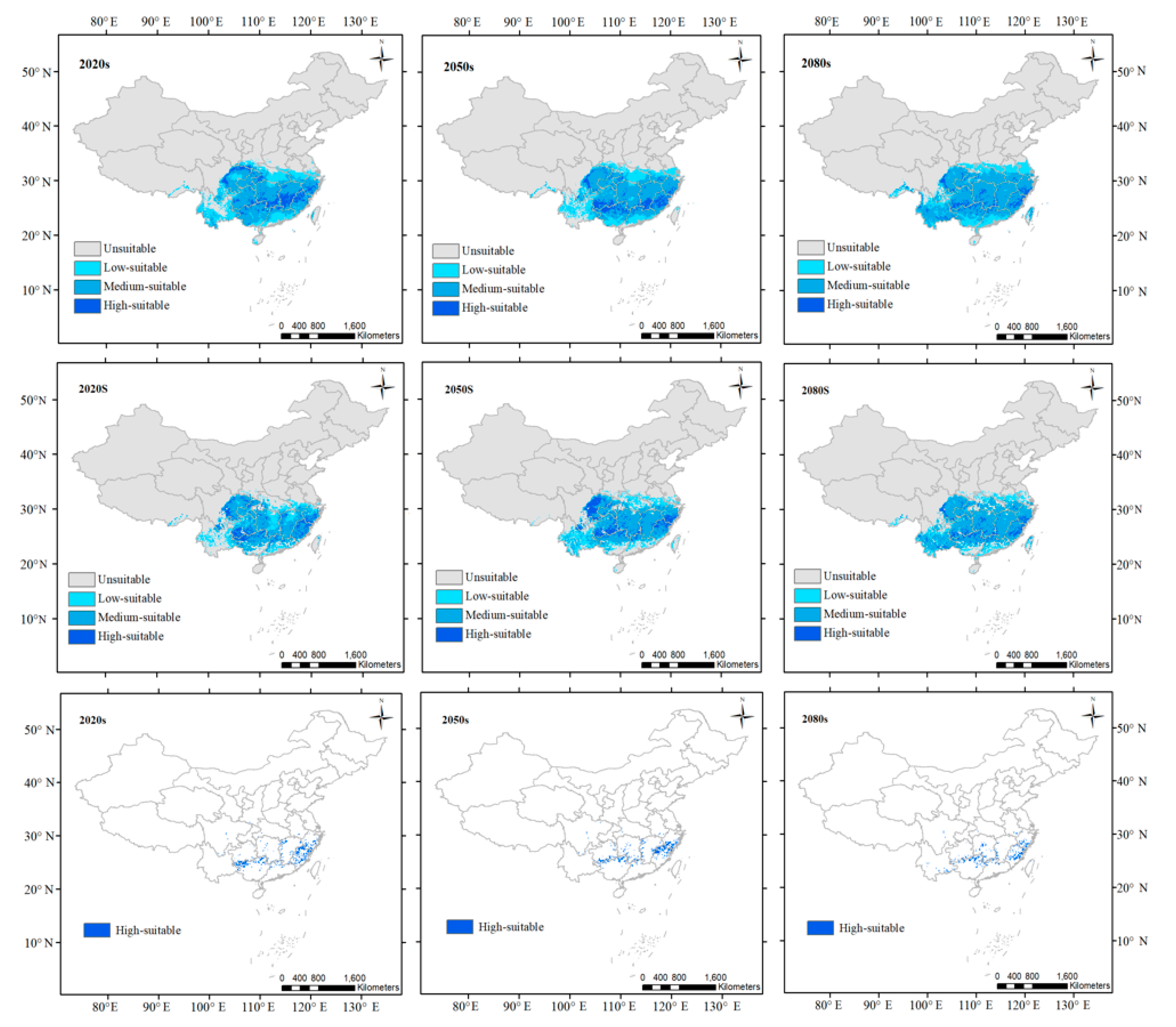
3. Results
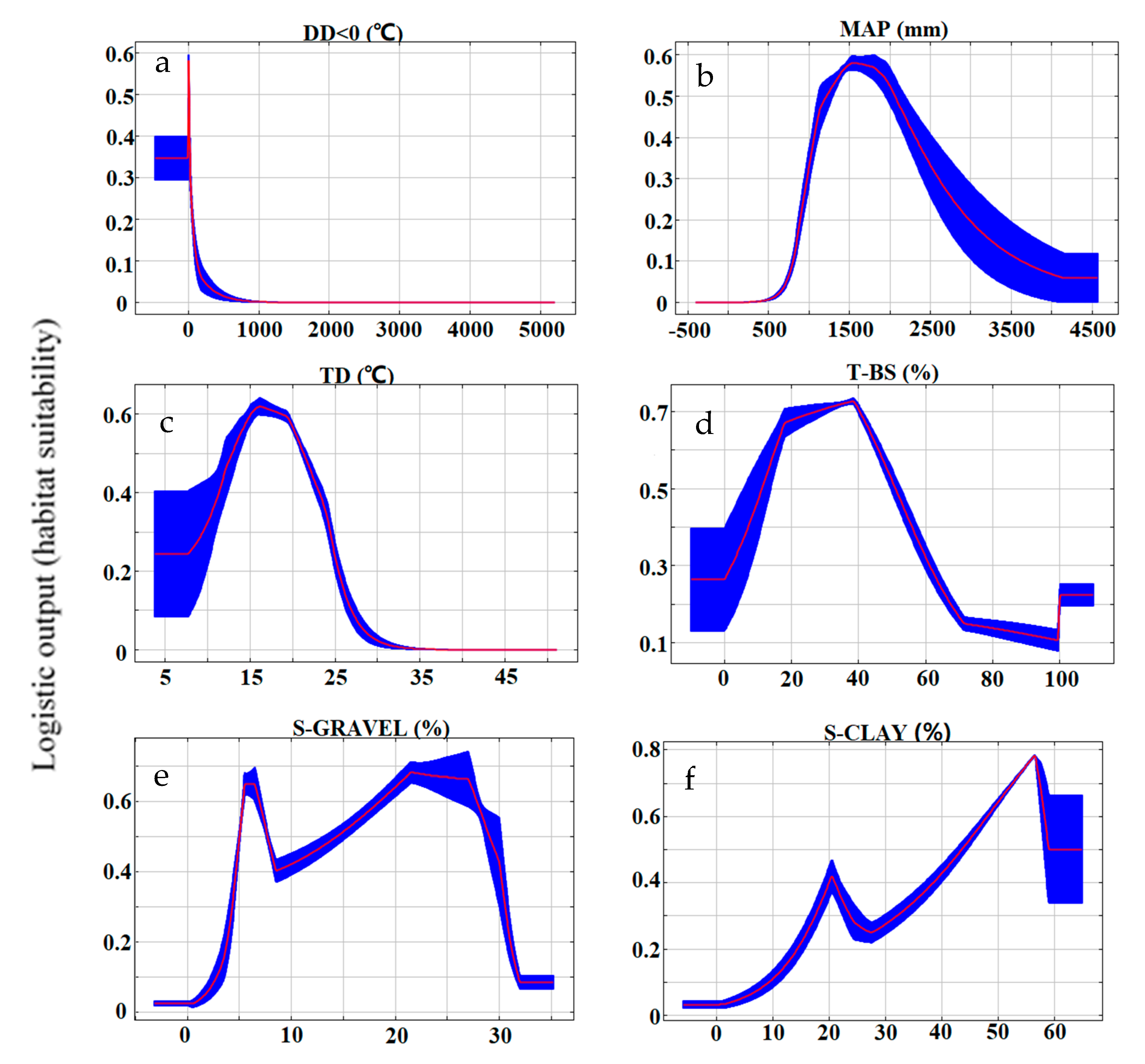
3.1. Model Performance and Contributing Variables
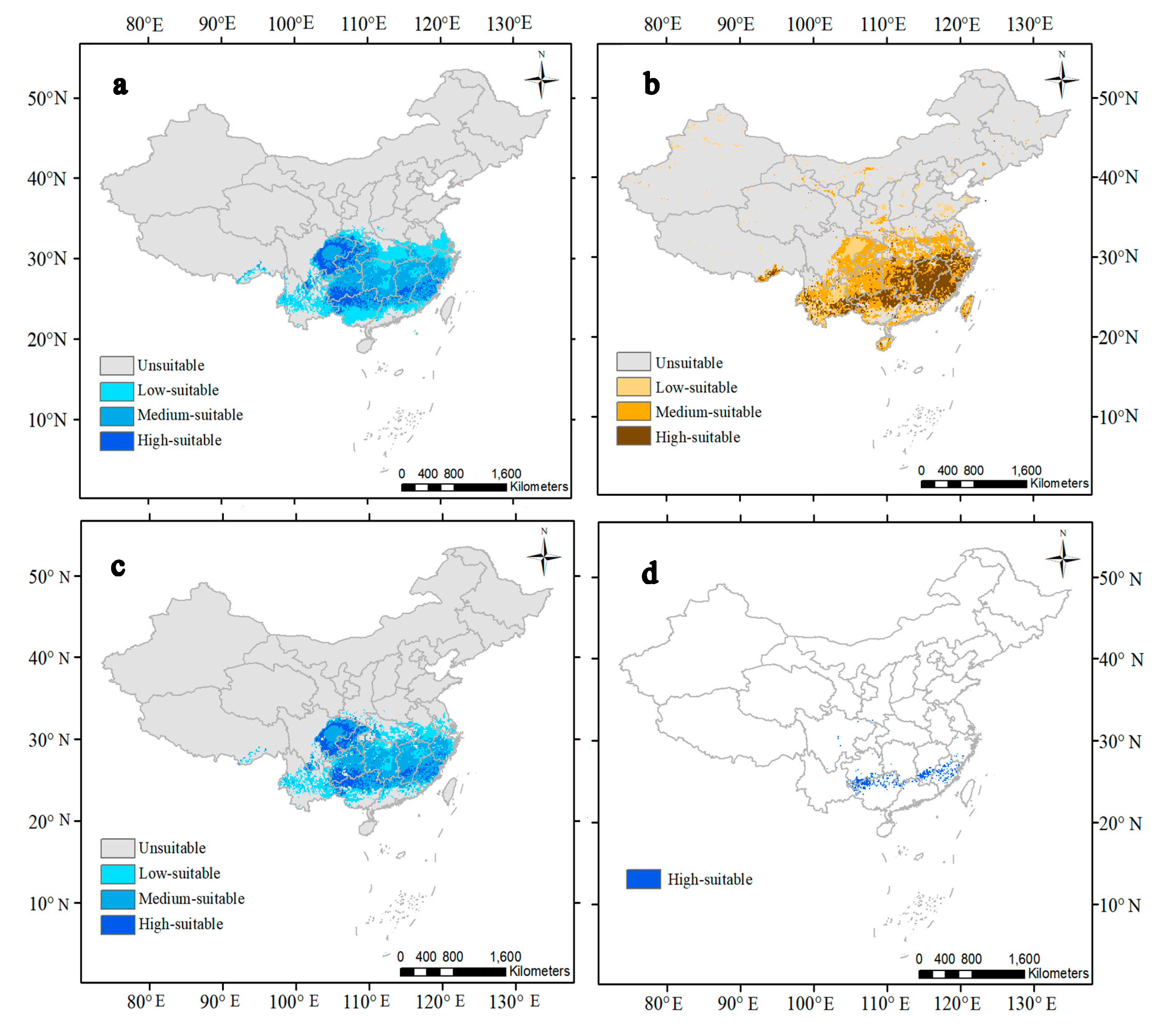
3.2. Predicted Suitable Habitats for the Current
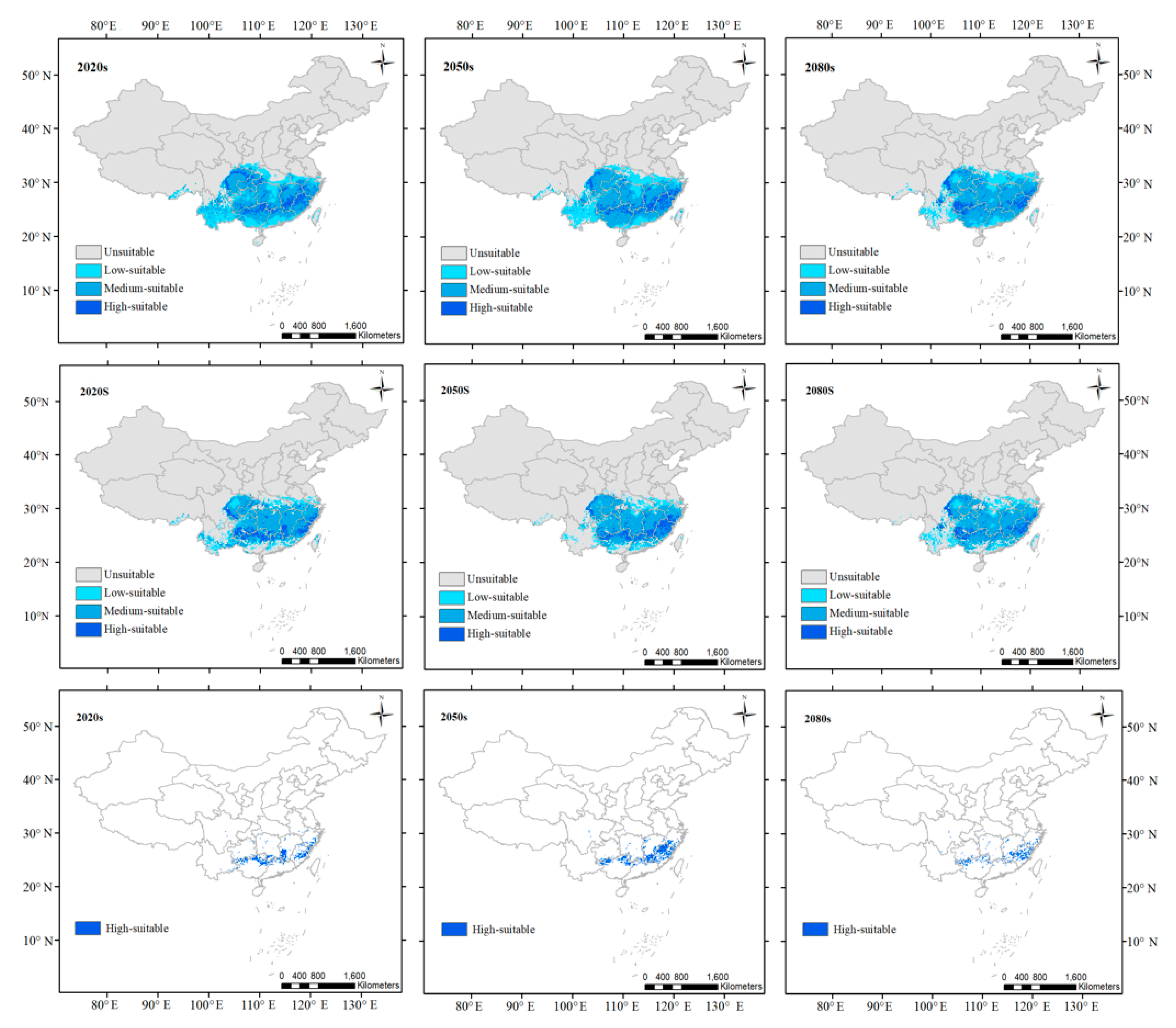
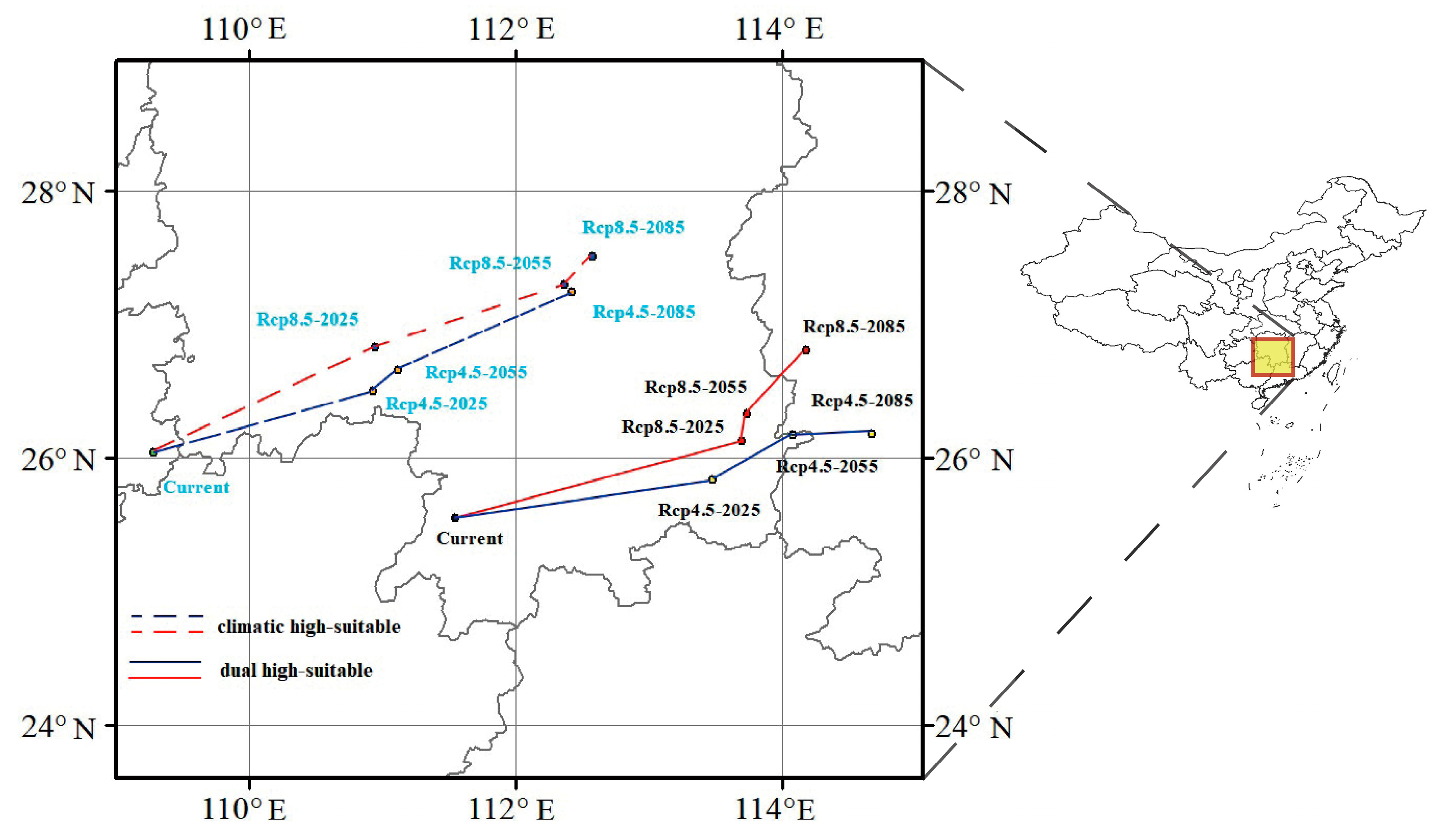
3.3. Projected Changes in Suitable Habitats for Future Periods
4. Discussion
4.1. Predicted Suitable Habitats Using Both Climatic and Soil Variables
4.2. Impact of Climate Change
4.3. Implications for Commercial Forest Management
4.4. Implications for Genetic Conservation
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hsiang, Y.H.; Hertzberg, R.; Hecht, S.; Liu, L.F. Camptothecin induces protein-linked DNA breaks via mammalian DNA topoisomerase I. J. Biol. Chem. 1985, 260, 14873–14878. [Google Scholar]
- Zhang, Y.; Jiang, K.; Qing, D.; Bing, H.; Jiang, J.; Wang, S.; Yan, C. Accumulation of camptothecin and 10-hydroxycamptothecin and the transcriptional expression of camptothecin biosynthetic genes in Camptotheca acuminata cambial meristematic and dedifferentiated cells. RSC Adv. 2017, 7, 12185–12193. [Google Scholar] [CrossRef]
- Lorence, A.; Nessler, C.L. Camptothecin, over four decades of surprising findings. Phytochemistry 2004, 65, 2735–2749. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Zhang, X.; Wang, R.; Liu, D.; Sun, J.; Tao, J. Herbaceous peony tryptophan decarboxylase confers drought and salt stresses tolerance. Environ. Exp. Bot. 2019, 162, 345–356. [Google Scholar] [CrossRef]
- Chen, I.C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid Range Shifts of Species Associated with High Levels of Climate Warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.; De Siqueira, M.F.; Grainger, A.; Hannah, L. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Peterson, M.L.; Doak, D.F.; Morris, W.F. Incorporating local adaptation into forecasts of species’ distribution and abundance under climate change. Glob. Chang. Biol. 2019, 25, 775–793. [Google Scholar] [CrossRef]
- Murray, N.J.; Keith, D.A.; Bland, L.M.; Ferrari, R.; Lyons, M.B.; Lucas, R.; Pettorelli, N.; Nicholson, E. The role of satellite remote sensing in structured ecosystem risk assessments. Sci. Total Environ. 2018, 619, 249–257. [Google Scholar] [CrossRef]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Peterson, A.T. Original Article: Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Li, G.; Du, S.; Wen, Z. Mapping the climatic suitable habitat of oriental arborvitae (Platycladus orientalis) for introduction and cultivation at a global scale. Sci. Rep. 2016, 6, 30009. [Google Scholar] [CrossRef]
- Guevara, L.; Gerstner, B.E.; Kass, J.M.; Anderson, R.P. Toward ecologically realistic predictions of species distributions: A cross-time example from tropical montane cloud forests. Glob. Chang. Biol. 2017, 24, 1511–1522. [Google Scholar] [CrossRef] [PubMed]
- Esfanjani, J.; Ghorbani, A.; Chahouki, M.A.Z. MaxEnt Modeling for Predicting Impacts of Environmental Factors on the Potential Distribution of Artemisia aucheri and Bromus tomentellus-Festuca ovina in Iran. Pol. J. Environ. Stud. 2018, 27, 1041–1047. [Google Scholar] [CrossRef]
- Mungi, N.A.; Coops, N.C.; Ramesh, K.; Rawat, G.S. How global climate change and regional disturbance can expand the invasion risk? Case study of Lantana camara invasion in the Himalaya. Biol. Invasions 2018, 20, 1849–1863. [Google Scholar] [CrossRef]
- Linhoss, A.C.; Underwood, W.V. Modeling Salt Panne Land-Cover Suitability under Sea-Level Rise. J. Coast. Res. 2016, 32, 1116–1125. [Google Scholar] [CrossRef]
- Ashoori, A.; Kafash, A.; Varasteh Moradi, H.; Yousefi, M.; Kamyab, H.; Behdarvand, N.; Mohammadi, S. Habitat modeling of the common pheasant Phasianus colchicus (Galliformes: Phasianidae) in a highly modified landscape: Application of species distribution models in the study of a poorly documented bird in Iran. Eur. Zool. J. 2018, 85, 372–380. [Google Scholar] [CrossRef]
- Araujo, M.B.; Peterson, A.T. Uses and misuses of bioclimatic envelope modeling. Ecology 2012, 93, 1527–1539. [Google Scholar] [CrossRef]
- Keith, D.A.; Akcakaya, H.R.; Thuiller, W.; Midgley, G.F.; Pearson, R.G.; Phillips, S.J.; Regan, H.M.; Araujo, M.B.; Rebelo, T.G. Predicting extinction risks under climate change: Coupling stochastic population models with dynamic bioclimatic habitat models. Biol. Lett. 2008, 4, 560–563. [Google Scholar] [CrossRef]
- Anderson, B.J.; Akcakaya, H.R.; Araujo, M.B.; Fordham, D.A.; Martinez-Meyer, E.; Thuiller, W.; Brook, B.W. Dynamics of range margins for metapopulations under climate change. Proc. R. Soc. B Biol. Sci. 2009, 276, 1415–1420. [Google Scholar] [CrossRef]
- Woodward, F.I.; Williams, B.G. Climate and plant distribution at global and local scales. Vegetation 1987, 69, 189–197. [Google Scholar] [CrossRef]
- Volis, S.; Zhang, Y.H.; Deng, T.; Dorman, M.; Blecher, M.; Abbott, R.J. Divergence and reproductive isolation between two closely related allopatric Iris species. Biol. J. Linn. Soc. 2019, 127, 377–389. [Google Scholar] [CrossRef]
- Zuquim, G.; Costa, F.R.C.; Tuomisto, H.; Moulatlet, G.M.; Figueiredo, F.O.G. The importance of soils in predicting the future of plant habitat suitability in a tropical forest. Plant Soil 2019, 450, 151–170. [Google Scholar] [CrossRef]
- Figueiredo, F.O.G.; Zuquim, G.; Tuomisto, H.; Moulatlet, G.M.; Balslev, H.; Costa, F.R.C. Beyond climate control on species range: The importance of soil data to predict distribution of Amazonian plant species. J. Biogeogr. 2018, 45, 190–200. [Google Scholar] [CrossRef]
- Syphard, A.D.; Franklin, J. Differences in spatial predictions among species distribution modeling methods vary with species traits and environmental predictors. Ecography 2009, 32, 907–918. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Varela, S.; Anderson, R.P.; Garciavaldes, R.; Fernandezgonzalez, F. Environmental filters reduce the effects of sampling bias and improve predictions of ecological niche models. Ecography 2014, 37, 1084–1091. [Google Scholar] [CrossRef]
- Wang, T.; Wang, G.; Innes, J.L.; Seely, B.; Chen, B. ClimateAP: An application for dynamic local downscaling of historical and future climate data in Asia Pacific. Front. Agric. Sci. Eng. 2017, 4, 448–458. [Google Scholar] [CrossRef]
- Fischer, G.; Velthuizen, H.V.; Shah, M.; Nachtergaele, F. Global AgroEcological Assessment for Agriculture in the 21st Century; International Institute for Applied Systems Analysis: Laxenburg, Austria, 2002. [Google Scholar]
- Koeling, R. Chunking with maximum entropy models. In Proceedings of the Conference on Computational Natural Language Learning, Lisbon, Portugal, 13–14 September 2000; pp. 139–141. [Google Scholar]
- Yang, X.Q.; Kushwaha, S.P.S.; Saran, S.; Xu, J.; Roy, P.S. Maxent modeling for predicting the potential distribution of medicinal plant, Justicia adhatoda L. in Lesser Himalayan foothills. Ecol. Eng. 2013, 51, 83–87. [Google Scholar] [CrossRef]
- Guo, J.; Liu, X.P.; Zhang, Q.; Zhang, D.F.; Liu, X. Prediction for the potential distribution area of Codonopsis pilosula at global scale based on Maxent model. J. Appl. Ecol. 2017, 28, 992–1000. [Google Scholar]
- Lu, C.Y.; Gu, W.; Dai, A.H.; Wei, H.Y. Assessing habitat suitability based on geographic information system (GIS) and fuzzy: A case study of Schisandra sphenanthera Rehd. et Wils. in Qinling Mountains, China. Ecol. Model. 2012, 242, 105–115. [Google Scholar] [CrossRef]
- Hanley, J.A.; Mcneil, B.J. The meaning and use of the area under a receiver operating characteristic (ROC) curve. Radiology 1982, 143, 29–36. [Google Scholar] [CrossRef]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Yao, L.; Meng, J.; Tao, J. Maxent modeling for predicting the potential geographical distribution of two peony species under climate change. Sci. Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef] [PubMed]
- Koralewski, T.E.; Wang, H.H.; Grant, W.E.; Byram, T.D. Plants on the move: Assisted migration of forest trees in the face of climate change. For. Ecol. Manag. 2015, 344, 30–37. [Google Scholar] [CrossRef]
- Keenan, R.J. Climate change impacts and adaptation in forest management: A review. Ann. For. Sci. 2015, 72, 145–167. [Google Scholar] [CrossRef]
- Pecchi, M.; Marchi, M.; Burton, V.; Giannetti, F.; Moriondo, M.; Bernetti, I.; Bindi, M.; Chirici, G. Species distribution modelling to support forest management. A literature review. Ecol. Model. 2019, 411, 108817. [Google Scholar] [CrossRef]
- Antúnez, P.; Suárez-Mota, E.M.; Valenzuela-Encinas, C.; Ruiz-Aquino, F. The Potential Distribution of Tree Species in Three Periods of Time under a Climate Change Scenario. Forests 2018, 9, 628. [Google Scholar] [CrossRef]
- Sehler, R.; Li, J.; Reager, J.; Ye, H. Investigating Relationship Between Soil Moisture and Precipitation Globally Using Remote Sensing Observations. J. Contemp. Water Res. Educ. 2019, 168, 106–118. [Google Scholar] [CrossRef]
- Wang, T.; Campbell, E.M.; O’Neill, G.A.; Aitken, S.N. Projecting future distributions of ecosystem climate niches: Uncertainties and management applications. For. Ecol. Manag. 2012, 279, 128–140. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Crookston, N.L.; Warwell, M.V.; Evans, J.S. Empirical analyses of plant-climate relationships for the western United States. Int. J. Plant Sci. 2006, 167, 1123–1150. [Google Scholar] [CrossRef]
- Wang, T.; Wang, G.; Innes, J.; Nitschke, C.; Kang, H. Climatic niche models and their consensus projections for future climates for four major forest tree species in the Asia—Pacific region. For. Ecol. Manag. 2016, 360, 357–366. [Google Scholar] [CrossRef]
- Mátyás, C.; Sun, G. Forests in a water limited world under climate change. Environ. Res. Lett. 2014, 9, 085001. [Google Scholar] [CrossRef]
- Aitken, S.N.; Yeaman, S.; Holliday, J.A.; Wang, T.; Curtis-McLane, S. Adaptation, migration or extirpation: Climate change outcomes for tree populations. Evol. Appl. 2008, 1, 95–111. [Google Scholar] [CrossRef] [PubMed]
- Campbell, E.M.; Antos, J.A. Distribution and severity of white pine blister rust and mountain pine beetle on whitebark pine in British Columbia. Can. J. For. Res. 2000, 30, 1051–1059. [Google Scholar] [CrossRef]
- Allendorf, F.W.; Luikart, G.; Aitken, S.N. Conservation and the Genetics of Populations, 2nd ed.; Blackwell Publishing: Malden, MA, USA, 2012; p. 624. [Google Scholar]
- Wang, Y.; Clack, B.; Li, S. Genetic Diversity in Camptotheca Decaisne. Pharm. Crop. 2014, 5, 120–134. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Description | Code | Description |
|---|---|---|---|
| MAT | Mean annual temperature (℃) | DD > 5 | Degree-days above 5 ℃, growing degree-days |
| MWMT | Mean warmest month temperature (℃) | DD < 0 | Degree-days below 0 ℃, chilling degree-days |
| MCMT | Mean coldest month temperature (℃) | NFFD | Number of frost-free days |
| TD | Temperature difference between MWMT and MCMT, or continentality (℃) | PAS | Precipitation as snow (mm) |
| MAP | Mean annual precipitation (mm) | EMT | Extreme minimum temperature over 30 years |
| EXT | Extreme maximum temperature over 30 years | Eref | Hargreaves reference evaporation |
| AHM | Annual heat:moisture index (MAT+10)/(MAP/1000)) | CMD | Hargreaves climatic moisture deficit |
| DD < 18 | Degree-days below 18 °C | DD > 18 | Degree-days above 18 °C |
| Code | Description | Code | Description |
|---|---|---|---|
| T-GRAVEL | Topsoil Gravel Content | S-GRAVEL | Subsoil Gravel Content |
| T-SAND | Topsoil Sand Fraction | S-SAND | Subsoil Sand Fraction |
| T-SILT | Topsoil Silt Fraction | S_SILT | Subsoil Silt Fraction |
| T-CLAY | Topsoil Clay Fraction | S-CLAY | Subsoil Clay Fraction |
| T_REF_BULK_DENSITY | Topsoil bulk density | S_REF_BULK_DENSITY | Subsoil bulk density |
| T-OC | Topsoil Organic Carbon | S-OC | Subsoil Organic Carbon |
| T-PH-H2O | Topsoil pH (H 2 O) | S-PH-H2O | Subsoil pH (H 2 O) |
| T-CEC-CLAY | Topsoil CEC (clay) | S-CEC-CLAY | Subsoil CEC (clay) |
| T-CEC-SOIL | Topsoil CEC (soil) | S-CEC-SOIL | Subsoil CEC (soil) |
| T-BS | Topsoil Base Saturation | S-BS | Subsoil Base Saturation |
| T-TEB | Topsoil TEB | S-TEB | Subsoil TEB |
| T-CACO3 | Topsoil Calcium Carbonate | S-CACO3 | Subsoil Calcium Carbonate |
| T-CASO4 | Topsoil Gypsum | S-CASO4 | Subsoil Gypsum |
| T-ESP | Topsoil Sodicity (ESP) | S-ESP | Subsoil Sodicity (ESP) |
| T-ECE | Topsoil Salinity (Elco) | S-ECE | Subsoil Salinity (Elco) |
| Model | Variable | Unit | Contribution (%) |
|---|---|---|---|
| DD < 0 | ℃ | 80.7 | |
| MAP | mm | 6.3 | |
| Climatic | TD | ℃ | 4.3 |
| CMD | mm | 3.8 | |
| PAS | mm | 3.1 | |
| DD > 18 | ℃ | 1.9 | |
| T-BS | % | 31.8 | |
| S-GRAVEL | % | 22.7 | |
| S-CLAY | % | 18.4 | |
| S-CASO4 | % | 7.1 | |
| T-CEC-CLAY | 5.8 | ||
| Soil | S-ESP | 2.9 | |
| S-CEC-CLAY | 2.9 | ||
| T-TEB | 2.7 | ||
| T-GRAVEL | % | 1.9 | |
| T-REF-BULK | 1.7 | ||
| T-SILT | % | 1.2 | |
| S-CACO3 | % | 0.9 | |
| DD < 0 | ℃ | 79.6 | |
| MAP | mm | 4.4 | |
| T-BS | % | 3.2 | |
| TD | ℃ | 2.8 | |
| CMD | mm | 2.7 | |
| DD > 18 | ℃ | 1.5 | |
| Climatic + Soil | S-CASO4 | % | 1.5 |
| T-GRAVEL | % | 1.3 | |
| PAS | mm | 0.6 | |
| T-CEC-CLAY | 0.5 | ||
| S-ESP | 0.4 | ||
| S-CLAY | % | 0.4 | |
| T-REF-BULK | 0.4 | ||
| S-GRAVEL | % | 0.2 | |
| T-SILT | % | 0.2 | |
| S-CEC-CLAY | 0.1 | ||
| S-CACO3 | % | 0.1 | |
| T-TEB | 0.1 |
| Classes | High-Suitable | Medium-Suitable | Low-Suitable | Unsuitable | ||||
|---|---|---|---|---|---|---|---|---|
| Area | Proportion | Area | Proportion | Area | Proportion | Area | Proportion | |
| km2 | % | km2 | % | km2 | % | km2 | % | |
| Climatic habitats | 260,119 | 2.7 | 799,626 | 8.3 | 693,652 | 7.2 | 7,880,658 | 81.8 |
| Soil habitats | 568,409 | 5.9 | 973,039 | 10.1 | 404,630 | 4.2 | 7,687,977 | 79.8 |
| Climatic habitats filtered by soil habitats | 246,400 | 2.6 | 733,200 | 7.6 | 463,000 | 4.8 | 8,179,400 | 85 |
| Dual high-suitable habitats | 83,600 | 0.87 | ||||||
| Time | Current Area (km2) | Change by 2020s (%) | Change by 2050s (%) | Change by 2080s (%) | |||
|---|---|---|---|---|---|---|---|
| Emissions Scenarios | RCP4.5 | RCP8.5 | RCP4.5 | RCP8.5 | RCP4.5 | RCP8.5 | |
| Climatic habitats | 1,758,000 | +7.2 | +11.3 | +7.7 | +10.1 | +9.5 | +25.3 |
| High-suitable habitats | 246,400 | −2.8 | −3.2 | −4.2 | −4.9 | −28.7 | −24.6 |
| Medium-suitable habitats | 733,200 | +14.8 | +18.6 | +17.4 | +23.1 | 35.2 | +59.8 |
| Low-suitable habitats | 463,000 | −4.1 | −10 | −8.6 | −19.9 | −20.6 | −29.6 |
| Total suitable habitats | 1,442,600 | +5.7 | +5.7 | +5.4 | +4.5 | +6.4 | +16.7 |
| Dual high-suitable habitats | 83,600 | +98 | +105.3 | +100.5 | +66.1 | −12.7 | −20.5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, L.; Sun, J.; Shi, Y.; Wang, G.; Wang, T. Predicting Suitable Habitats of Camptotheca acuminata Considering Both Climatic and Soil Variables. Forests 2020, 11, 891. https://doi.org/10.3390/f11080891
Feng L, Sun J, Shi Y, Wang G, Wang T. Predicting Suitable Habitats of Camptotheca acuminata Considering Both Climatic and Soil Variables. Forests. 2020; 11(8):891. https://doi.org/10.3390/f11080891
Chicago/Turabian StyleFeng, Lei, Jiejie Sun, Yuanbao Shi, Guibin Wang, and Tongli Wang. 2020. "Predicting Suitable Habitats of Camptotheca acuminata Considering Both Climatic and Soil Variables" Forests 11, no. 8: 891. https://doi.org/10.3390/f11080891
APA StyleFeng, L., Sun, J., Shi, Y., Wang, G., & Wang, T. (2020). Predicting Suitable Habitats of Camptotheca acuminata Considering Both Climatic and Soil Variables. Forests, 11(8), 891. https://doi.org/10.3390/f11080891