Diversity and Pathogenicity of Phytophthora Species Associated with Declining Alder Trees in Italy and Description of Phytophthora alpina sp. nov

 ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites, Field Surveys and Sampling Procedure
2.2. Phytophthora Isolation and Identification
2.3. DNA Extraction, Polymerase Chain Reaction (PCR) Amplification, and Sequencing
2.4. Phylogenetic Analysis
2.5. Pathogenicity Test
2.6. Data Analysis
3. Results
3.1. Field Survey
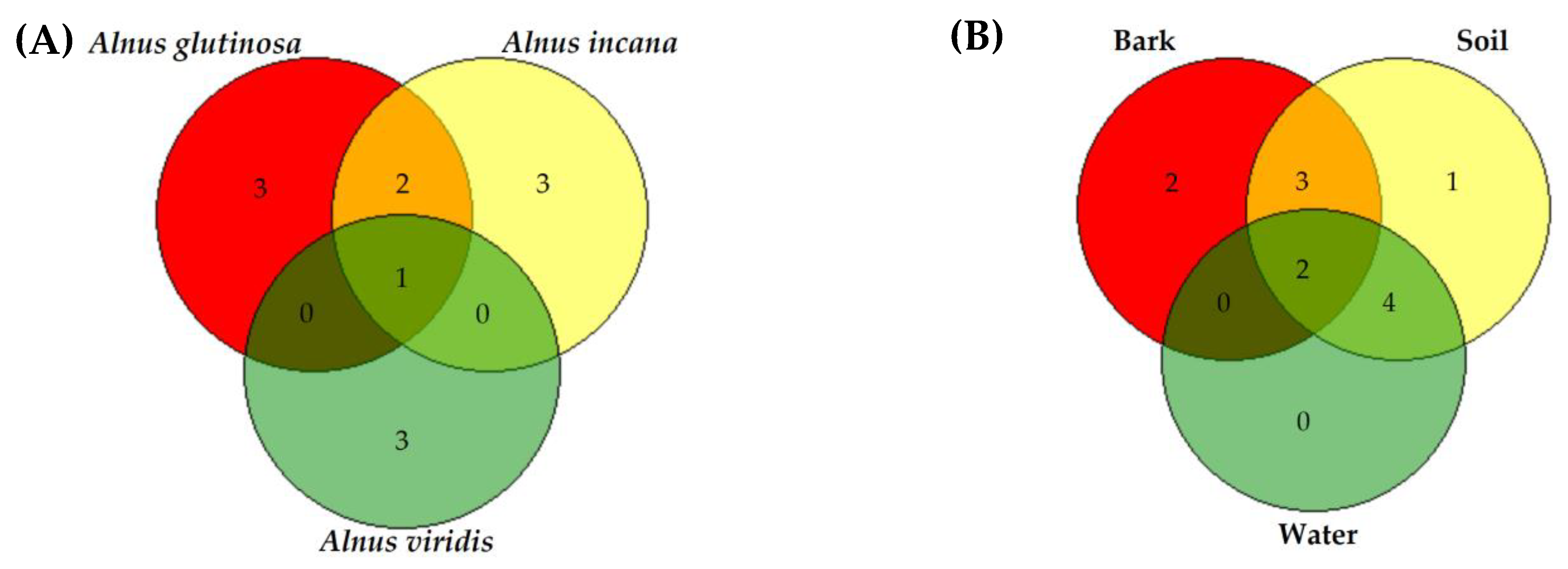
3.2. Phytophthora Diversity in Declining Alder Stands
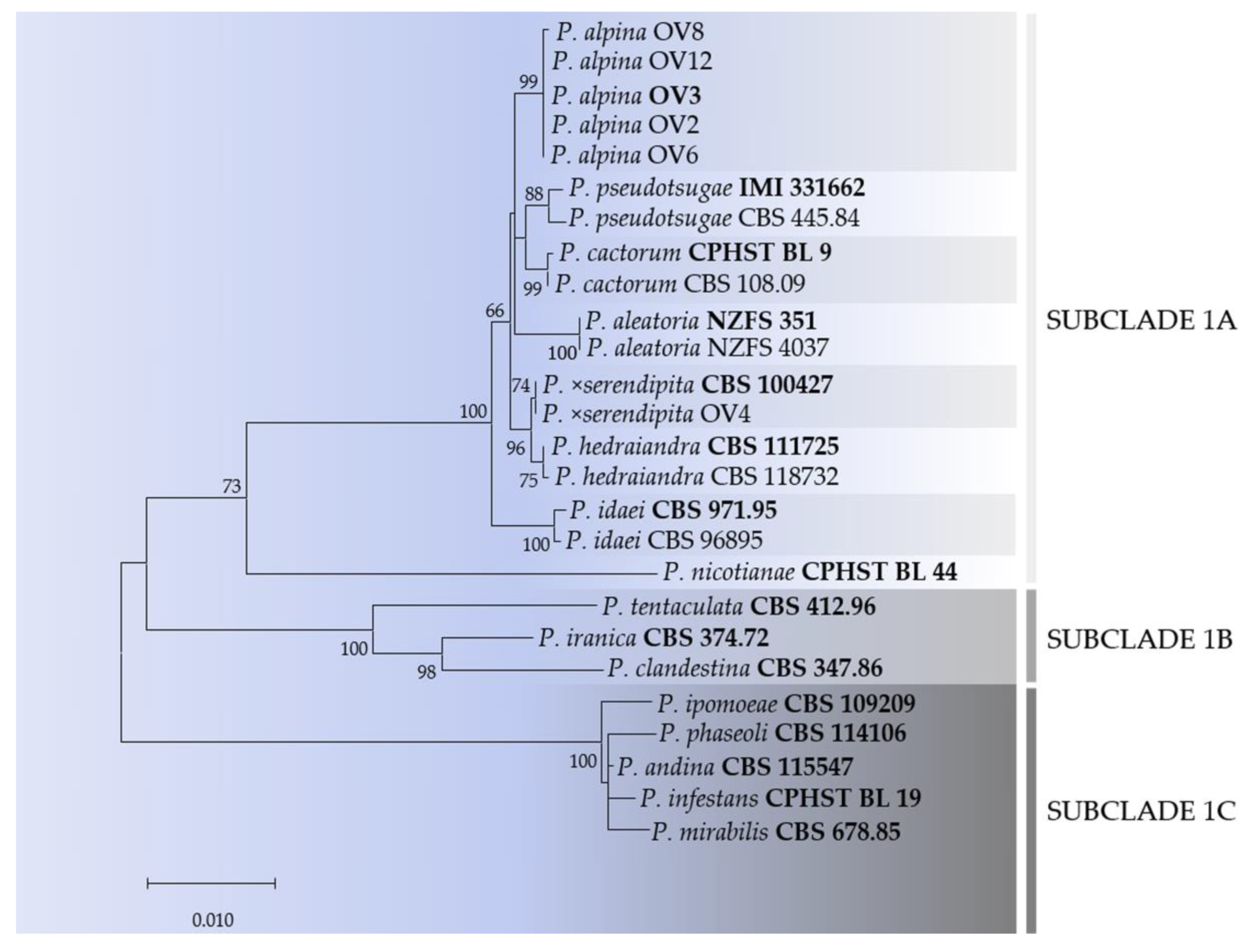
3.3. DNA Phylogeny of Phytophthora Clade 1
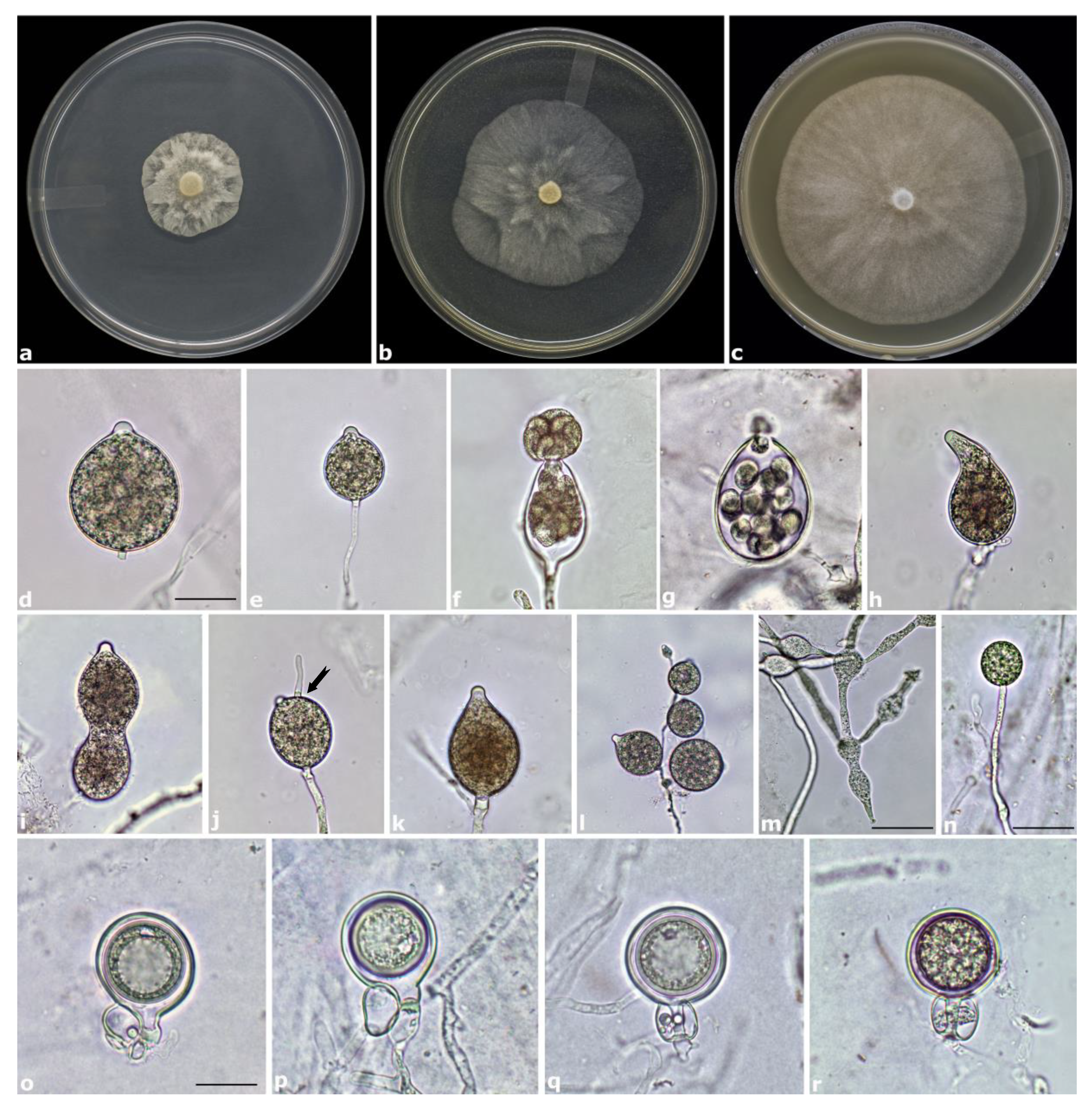
3.4. Taxonomy
3.5. Pathogenicity
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chen, Z.D.; Li, J.H. Phylogenetics and biogeography of Alnus (Betulaceae) inferred from sequences of nuclear ribosomal DNA its region. Int. J. Plant Sci. 2004, 165, 325–335. [Google Scholar] [CrossRef]
- Ren, B.Q.; Xiang, X.G.; Chen, Z.D. Species identification of Alnus (Betulaceae) using nrDNA and cpDNA genetic markers. Mol. Ecol. Resour. 2010, 10, 594–605. [Google Scholar] [CrossRef]
- Wipfli, M.; Musslewhite, J. Density of red alder (Alnus rubra) in headwaters influences invertebrate and detritus subsidies to downstream fish habitats in Alaska. Hydrobiologia 2004, 520, 153–163. [Google Scholar] [CrossRef]
- Claessens, H.; Oosterbaan, A.; Savill, P.; Rondeux, J. A review of the characteristics of black alder (Alnus glutinosa (L.) Gaertn.) and their implications for silvicultural practices. Forestry 2010, 83, 163–175. [Google Scholar] [CrossRef]
- Handa, I.T.; Aerts, R.; Berendse, F.; Berg, M.P.; Bruder, A.; Butenschoen, O.; Chauvet, E.; Gessner, M.O.; Jabiol, J.; Makkonen, M.; et al. Consequences of biodiversity loss for litter decomposition across biomes. Nature 2014, 509, 218–221. [Google Scholar] [CrossRef]
- Brasier, C.; Rose, J.; Gibbs, J. An unusual Phytophthora associated with widespread alder mortality in Britain. Plant Pathol. 1995, 44, 999–1007. [Google Scholar] [CrossRef]
- Adams, G.C.; Catal, M.; Trummer, L.; Hansen, E.M.; Reeser, P.; Worrall, J.J. Phytophthora alni subsp. uniformis found in Alaska beneath thinleaf alders. Plant Health Prog. 2008, 9, 38. [Google Scholar] [CrossRef]
- Cerny, K.; Strnadova, V. Phytophthora alni decline: Disease symptoms, causal agent and its distribution in the Czech Republic. Plant Prot. Sci. 2010, 46, 12–18. [Google Scholar] [CrossRef]
- Sims, L.L.; Sutton, W.; Reeser, P.; Hansen, E.M. The Phytophthora species assemblage and diversity in riparian alder ecosystems of western Oregon, USA. Mycologia 2015, 107, 889–902. [Google Scholar] [CrossRef]
- Bjelke, U.; Boberg, J.; Oliva, J.; Tattersdill, K.; McKie, B.G. Dieback of riparian alder caused by the Phytophthora alni complex: Projected consequences for stream ecosystems. Freshw. Biol. 2016, 61, 565–579. [Google Scholar] [CrossRef]
- Husson, C.; Aguayo, J.; Revellin, C.; Frey, P.; Ioos, R.; Marcais, B. Evidence for homoploid speciation in Phytophthora alni supports taxonomic reclassification in this species complex. Fungal Genet. Biol. 2015, 77, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Brasier, C.M.; Kirk, S.A. Comparative aggressiveness of standard and variant hybrid alder phytophthoras, Phytophthora cambivora and other Phytophthora species on bark of Alnus, Quercus and other woody hosts. Plant Pathol. 2001, 50, 218–229. [Google Scholar] [CrossRef]
- Trzewik, A.; Orlikowski, L.B.; Oszako, T.; Nowakowska, J.A.; Orlikowska, T. The characterization of Phytophthora isolates obtained from diseased Alnus glutinosa in Poland. Balt. For. 2015, 21, 44–50. [Google Scholar]
- Aday Kaya, A.G.; Lehtijärvi, A.; Şaşmaz, Y.; Nowakowska, J.A.; Oszako, T.; Doğmuş Lehtijärvi, H.T.; Woodward, S. Phytophthora species detected in the rhizosphere of Alnus glutinosa stands in the floodplain forests of Western Turkey. For. Pathol. 2018, 48, 11–14. [Google Scholar] [CrossRef]
- Seddaiu, S.; Linaldeddu, B.T. First Report of Phytophthora acerina, P. plurivora, and P. pseudocryptogea associated with declining common alder trees in Italy. Plant Dis. 2020, 104, 1874. [Google Scholar] [CrossRef]
- Zamora-Ballesteros, C.; Haque, M.M.U.; Diez, J.J.; Martín-García, J. Pathogenicity of Phytophthora alni complex and P. plurivora in Alnus glutinosa seedlings. For. Pathol. 2017, 47, e12299. [Google Scholar] [CrossRef]
- Rooney-Latham, S.; Blomquist, C.L.; Pastalka, T.; Costello, L. Collar rot on Italian alder trees in California caused by Phytophthora siskiyouensis. Plant Health Prog. 2009, 10, 20. [Google Scholar] [CrossRef]
- Navarro, S.; Sims, L.; Hansen, E. Pathogenicity to alder of Phytophthora species from riparian ecosystems in western Oregon. For. Pathol. 2015, 45, 358–366. [Google Scholar] [CrossRef]
- Kajba, D.; Gračan, J. EUFORGEN Technical Guidelines for Genetic Conservation and Use for Black Alder (Alnus glutinosa); International Plant Genetic Resources Institute: Rome, Italy, 2003; p. 4. [Google Scholar]
- Ducci, F.; Tani, A. EUFORGEN Technical Guidelines for Genetic Conservation and Use of Italian Alder (Alnus cordata); Bioversity International: Rome, Italy, 2009; p. 6. [Google Scholar]
- Mauri, A.; Caudullo, G. Alnus viridis in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the European Union: Luxembourg, 2016; p. e01f0e4+. [Google Scholar]
- Houston Durrant, T.; de Rigo, D.; Caudullo, G. Alnus incana in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the European Union: Luxembourg, 2016; p. e01ff87+. [Google Scholar]
- Pisetta, M.; Montecchio, L.; Longa, C.M.O.; Salvadori, C.; Zottele, F.; Maresi, G. Green alder decline in the Italian Alps. For. Ecol. Manag. 2012, 281, 75–83. [Google Scholar] [CrossRef]
- Huberli, D.; Hardy, G.E.S.J.; White, D.; Williams, N.; Burgess, T.I. Fishing for Phytophthora from Western Australia’s waterways: A distribution and diversity survey. Australas. Plant Pathol. 2013, 42, 251–260. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Mulas, A.A.; Bregant, C.; Piras, G.; Montecchio, L. First Report of Phytophthora pistaciae causing root and collar rot on nursery plants of Pistacia lentiscus in Italy. Plant Dis. 2020, 104, 1564. [Google Scholar] [CrossRef]
- Erwin, C.D.; Ribeiro, O.K. Phytophthora Diseases Worldwide; APS Press: Saint Paul, MN, USA, 1996. [Google Scholar]
- Crous, P.W.; Gams, W.; Stalpers, J.A.; Robert, V.; Stegehuis, G. MycoBank: An online initiative to launch mycology into the 21st century. Stud. Mycol. 2004, 50, 19–22. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols, a Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Cooke, D.E.L.; Drenth, A.; Duncan, J.M.; Wagels, G.; Brasier, C.M. A molecular phylogeny of Phytophthora and related oomycetes. Fungal Genet. Biol. 2000, 30, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.N.; Tooley, P.W. Phylogenetic relationships among Phytophthora species inferred from sequence analysis of mitochondrially encoded cytochrome oxidase I and II genes. Mycologia 2003, 95, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Kroon, L.P.N.M.; Bakker, F.T.; Van Den Bosch, G.B.M.; Bonants, P.J.M.; Flier, W.G. Phylogenetic analysis of Phytophthora species based on mitochondrial and nuclear DNA sequences. Fungal Genet. Biol. 2004, 41, 766–782. [Google Scholar] [CrossRef] [PubMed]
- Linaldeddu, B.T.; Franceschini, A.; Alves, A.; Phillips, A.J.L. Diplodia quercivora sp. nov.: A new species of Diplodia found on declining Quercus canariensis trees in Tunisia. Mycologia 2013, 105, 1266–1274. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Nucleic Acids Symposium Series; Oxfrod University Press: Oxford, UK, 1999; Volume 41, pp. 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Schoebel, C.N.; Stewart, J.; Gruenwald, N.J.; Rigling, D.; Prospero, S. Population history and pathways of spread of the plant pathogen Phytophthora plurivora. PLoS ONE 2014, 9, e85368. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Bregant, C.; Montecchio, L.; Favaron, F.; Sella, L. First report of Phytophthora acerina, P. pini and P. plurivora causing root rot and sudden death on olive trees in Italy. Plant Dis. 2020, 104, 996. [Google Scholar] [CrossRef]
- Balci, Y.; Balci, S.; Eggers, J.; MacDonald, W.L.; Juzwik, J.; Long, R.P.; Gottschalk, K.W. Phytophthora spp. associated with forest soils in eastern and north-central US oak ecosystems. Plant Dis. 2007, 91, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Safaiefarahani, B.; Mostowfizadeh-Ghalamfarsa, R.; Hardy, G.E.S.J.; Burgess, T.I. Re-evaluation of the Phytophthora cryptogea species complex and the description of a new species, Phytophthora pseudocryptogea sp. nov. Mycol. Prog. 2015, 14, 108. [Google Scholar] [CrossRef]
- Khaliq, I.; Hardy, G.E.S.J.; Mc Dougall, K.L.; Burgess, T.I. Phytophthora species isolated from alpine and sub-alpine regions of Australia, including the description of two new species; Phytophthora cacuminis sp. nov. and Phytophthora oreophila sp. nov. Fungal Biol. 2018, 123, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Bose, T.; Wingfield, M.J.; Roux, J.; Vivas, M.; Burgess, T.I. Community composition and distribution of Phytophthora species across adjacent native and non-native forests of South Africa. Fungal Ecol. 2018, 36, 17–25. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Bregant, C.; Ruzzon, B.; Montecchio, L. Coniella granati and Phytophthora palmivora the main pathogens involved in pomegranate dieback and mortality in north-eastern Italy. Ital. J. Mycol. 2020, 49, 92–100. [Google Scholar]
- Scanu, B.; Linaldeddu, B.T.; Franceschini, A. First report of Phytophthora pseudosyringae associated with ink disease of Castanea sativa in Italy. Plant Dis. 2010, 94, 1068. [Google Scholar] [CrossRef]
- Fajardo, S.N.A.; Valenzuela, S.B.; Dos Santos, A.F.C.; González, M.P.A.; Sanfuentes, E.A. Phytophthora pseudosyringae associated with the mortality of Nothofagus obliqua in a pure stand in central-southern Chile. For. Pathol. 2017, 47, e12361. [Google Scholar] [CrossRef]
- Wickland, A.C.; Jensen, C.E.; Rizzo, D.M. Geographic distribution, disease symptoms and pathogenicity of Phytophthora nemorosa and Phytophthora pseudosyringae in California, USA. For. Pathol. 2008, 38, 288–298. [Google Scholar] [CrossRef]
- Hansen, E.M.; Reeser, P.W.; Sutton, W. Ecology and pathology of Phytophthora ITS clade 3 species in forests in western Oregon, USA. Mycologia 2017, 109, 100–114. [Google Scholar] [CrossRef]
- Reeser, P.W.; Hansen, E.M.; Sutton, W.; Remigi, P.; Adams, G.C. Phytophthora species in forest streams in Oregon and Alaska. Mycologia 2011, 103, 22–35. [Google Scholar] [CrossRef]
- Aghighi, S.; Hardy, G.E.S.J.; Scott, J.K.; Burgess, T.I. Phytophthora bilorbang sp. nov., a new species associated with the decline of Rubus anglocandicans (European blackberry) in Western Australia. Eur. J. Plant Pathol. 2012, 133, 841–855. [Google Scholar] [CrossRef]
- Nagel, J.H.; Slippers, B.; Wingfield, M.J.; Gryzenhout, M. Multiple Phytophthora species associated with a single riparian ecosystem in South Africa. Mycologia 2015, 107, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Matsiakh, I.; Oszako, T.; Kramarets, V.; Nowakowska, J.A. Phytophthora and Pythium species detected in rivers of the Polish-Ukrainian border areas. Balt. For. 2016, 22, 230–238. [Google Scholar]
- Hong, C.X.; Gallegly, M.E.; Richardson, P.A.; Kong, P.; Moorman, J.D.; Lea-Cox, J.D.; Ross, D.S. Phytophthora hydropathica, a new pathogen identified from irrigation water, Rhododendron catawbiense and Kalmia latifolia. Plant Pathol. 2010, 59, 913–921. [Google Scholar] [CrossRef]
- Hulvey, J.; Gobena, D.; Finley, L.; Lamour, K. Co-occurrence and genotypic distribution of Phytophthora species recovered from watersheds and plant nurseries of eastern Tennessee. Mycologia 2010, 102, 1127–1133. [Google Scholar] [CrossRef]
- Ginetti, B.; Moricca, S.; Squires, J.N.; Cooke, D.E.L.; Ragazzi, A.; Jung, T. Phytophthora acerina sp. nov., a new species causing bleeding cankers and dieback of Acer pseudoplatanus trees in planted forests in northern Italy. Plant Pathol. 2014, 63, 858–876. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Sites | Species | Elevation (m a. s. l.) | Geographic Coordinates | Number of Samples | |
|---|---|---|---|---|---|
| 1 | Black alder | 56 | 40°51′10″ N | 9°00′09″ E | 13 (S), 13 (R), 130 (L) |
| 2 | Grey alder | 1412 | 46°29′31″ N | 12°33′47″ E | 9 (S), 10 (R), 50 (L) |
| 3 | Green alder | 1803 | 46°28′38″ N | 12°35′36″ E | 18 (S), 18 (R) |
| Species | Collection No. | Host | GenBank Accession Number | ||
|---|---|---|---|---|---|
| ITS | Btub | Cox1 | |||
| P. aleatoria | NZFS 351 | Pinus radiata | MK282205 | MK294168 | MK294173 |
| P. aleatoria | NZFS 4037 | P. radiata | MK282209 | MK294172 | MK294177 |
| P. alpina | OV2 | Alnus viridis | MT707331 | MT729672 | MT729667 |
| P. alpina | OV3 | A. viridis | MT707332 | MT729673 | MT729668 |
| P. alpina | OV6 | A. viridis | MT707333 | MT729674 | MT729669 |
| P. alpina | OV8 | A. viridis | MT707334 | MT729675 | MT729670 |
| P. alpina | OV12 | A. viridis | MT707335 | MT729676 | MT729671 |
| P. andina | CBS 115547 | Solanum brevifolium | MK496515 | EU080183 | MH136846 |
| P. cactorum | CPHST BL9 | Syringa vulgaris | MG783385 | MH493911 | MH136858 |
| P. cactorum | CBS 108.09 | Cactus sp. | Q-bank | MH493911 | Q-bank |
| P. clandestina | CBS 347.86 | Trifolium subterraneum | MG865477 | EU079867 | MH136873 |
| P. hedraiandra | CBS 111725 | Viburnum sp. | MG865504 | KX250398 | MH136898 |
| P. hedraiandra | CBS 118732 | Unknown | Q-bank | Q-bank | Q-bank |
| P. idaei | CBS 971.95 | Rubus idaeus | HQ261579 | EU080130 | HQ261326 |
| P. idaei | CBS 968.95 | R. idaeus | HQ643246 | Q-bank | HQ708308 |
| P. infestans | CPHSTBL19 | Solanum tuberosum | HQ261589 | EU079626 | HQ261336 |
| P. ipomoeae | CBS 109209 | Ipomoea longipedunculata | MG865518 | EU080831 | MH136912 |
| P. iranica | CBS 374.72 | Solanum melongena | MG865519 | EU080112 | MH136913 |
| P. mirabilis | CBS 678.85 | Mirabilis jalapa | MG865541 | MH493979 | MH136984 |
| P. nicotianae | CPHSTBL44 | Nicotiana tabacum | MG865550 | MH493985 | MH136943 |
| P. phaseoli | CBS 114106 | Phaseolus lunatus | MG865564 | EU080762 | MH136956 |
| P. pseudotsugae | IMI 331662 | Pseudotsuga menziesii | HQ261654 | EU080427 | HQ261401 |
| P. pseudotsugae | CBS 445.84 | P. menziesii | HQ643331 | Q-bank | HQ708380 |
| P. tentaculata | CBS 412.96 | Argyranthemum frutescens | MG865591 | EU080154 | MH136983 |
| P. ×serendipita | CBS 100427 | Idesia policarpa | MG865599 | MH494027 | MH477762 |
| P. ×serendipita | OV4 | Alnus viridis | MT707344 | MT729666 | MT729665 |
| Species | Accession Number | ITS Clade | Black alder | Grey Alder | Green Alder | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| (S) | (R) | (W) | (S) | (R) | (W) | (S) | (R) | |||
| P. acerina | MT707330 | 2 | 2 | 3 | - | - | 2 | - | - | - |
| P. alpina | MT707332 | 1 | - | - | - | - | - | - | 4 | 12 |
| P. bilorbang | MT707336 | 6 | - | 4 | 9 | - | - | - | - | - |
| P. cactorum | MT707337 | 1 | - | - | - | 1 | - | - | - | - |
| P. gonapodyides | MT707338 | 6 | - | - | - | - | 1 | 13 | - | - |
| P. hydropathica | MT707339 | 9 | - | 7 | 11 | - | - | - | - | - |
| P. lacustris | MT707340 | 6 | - | 2 | 9 | - | - | - | - | - |
| P. plurivora | MT707341 | 2 | 12 | 13 | 36 | 3 | 9 | 11 | - | - |
| P. pseudocryptogea | MT707342 | 8 | 5 | 13 | 18 | 2 | 4 | - | 2 | 6 |
| P. pseudosyringae | MT707343 | 3 | - | - | - | - | - | - | 7 | 5 |
| P. ×serendipita | MT707344 | 1 | - | - | - | - | - | - | - | 1 |
| Phytophthora sp. | MT707345 | 6 | - | - | 5 | - | - | - | - | |
| Species | Isolate | Mean Lesion Length (cm) * | Exudates | Wilted Foliage | Re-Isolation (%) |
|---|---|---|---|---|---|
| Phytophthora alpina | OV2 | 0.9 ± 0.2c | no | no | 100 |
| Phytophthora alpina | OV3 | 1.1 ± 0.2c | no | no | 100 |
| Phytophthora pseudocryptogea | OV5 | 2.8 ± 1.1b | yes | no | 100 |
| Phytophthora pseudosyringae | OV13 | 4.3 ± 1.4a | yes | yes | 100 |
| Phytophthora ×serendipita | OV4 | 3.4 ± 0.5ab | yes | no | 100 |
| Control | - | 0.7 ± 0.1c | no | no | - |
| LSD critical value | 2.01 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bregant, C.; Sanna, G.P.; Bottos, A.; Maddau, L.; Montecchio, L.; Linaldeddu, B.T. Diversity and Pathogenicity of Phytophthora Species Associated with Declining Alder Trees in Italy and Description of Phytophthora alpina sp. nov. Forests 2020, 11, 848. https://doi.org/10.3390/f11080848
Bregant C, Sanna GP, Bottos A, Maddau L, Montecchio L, Linaldeddu BT. Diversity and Pathogenicity of Phytophthora Species Associated with Declining Alder Trees in Italy and Description of Phytophthora alpina sp. nov. Forests. 2020; 11(8):848. https://doi.org/10.3390/f11080848
Chicago/Turabian StyleBregant, Carlo, Gian Paolo Sanna, Adriano Bottos, Lucia Maddau, Lucio Montecchio, and Benedetto T. Linaldeddu. 2020. "Diversity and Pathogenicity of Phytophthora Species Associated with Declining Alder Trees in Italy and Description of Phytophthora alpina sp. nov" Forests 11, no. 8: 848. https://doi.org/10.3390/f11080848
APA StyleBregant, C., Sanna, G. P., Bottos, A., Maddau, L., Montecchio, L., & Linaldeddu, B. T. (2020). Diversity and Pathogenicity of Phytophthora Species Associated with Declining Alder Trees in Italy and Description of Phytophthora alpina sp. nov. Forests, 11(8), 848. https://doi.org/10.3390/f11080848