Long-Term Effect of Lophodermium Needle Cast on The Growth of Scots Pine and Implications for Financial Outcomes



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Material
2.2. Statistical Analysis
2.3. Evaluation of the Financial Outcome
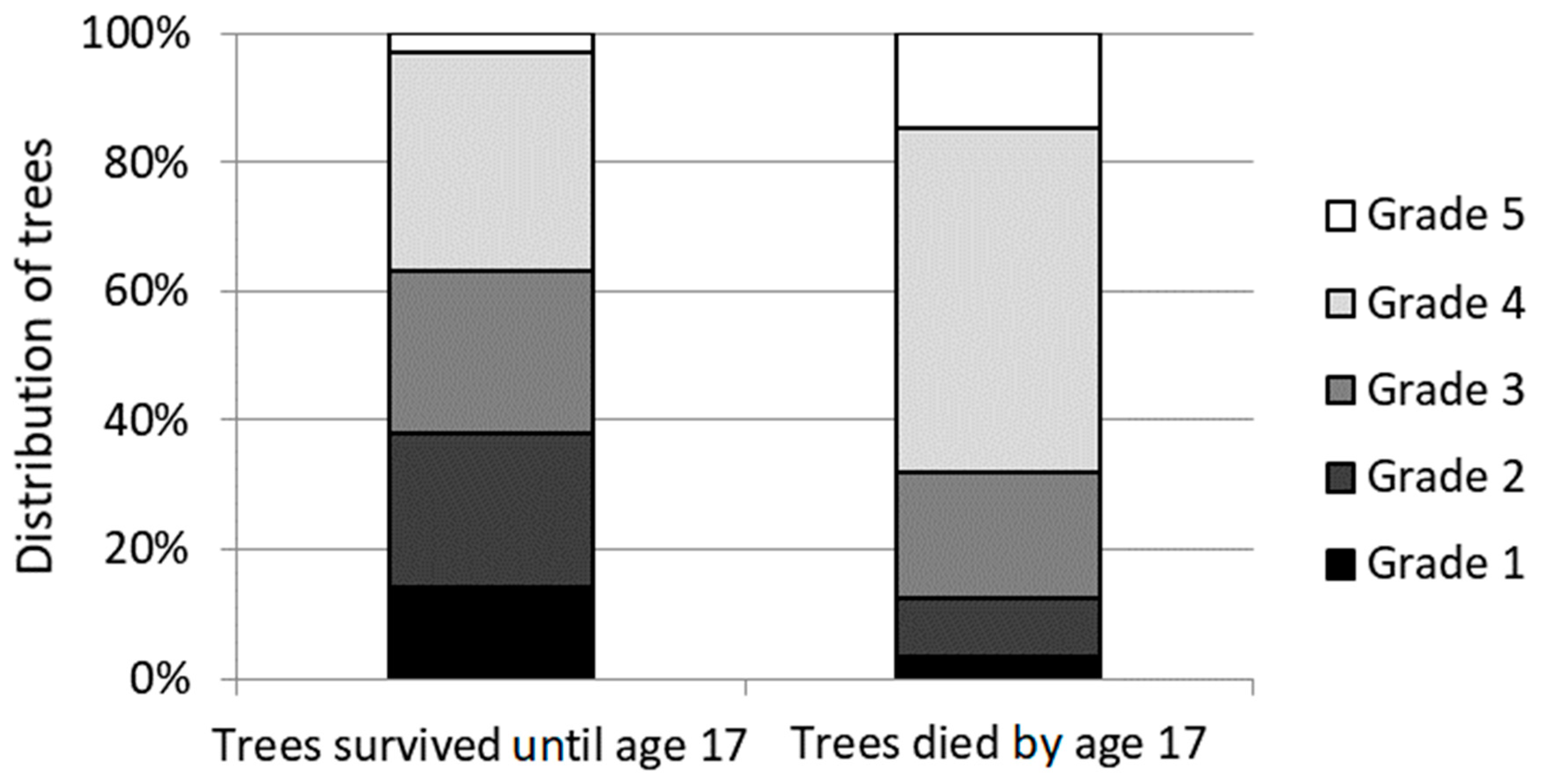
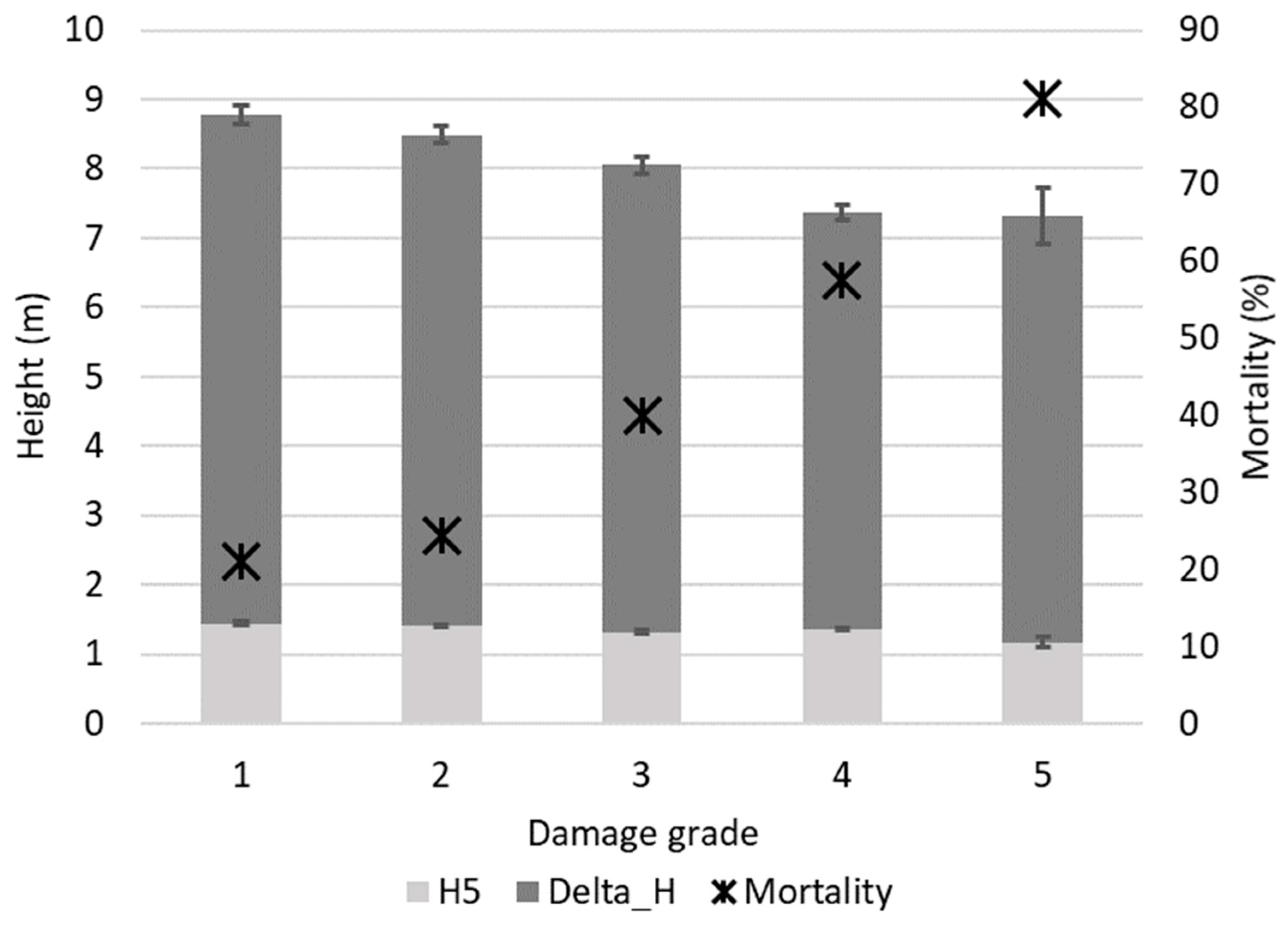
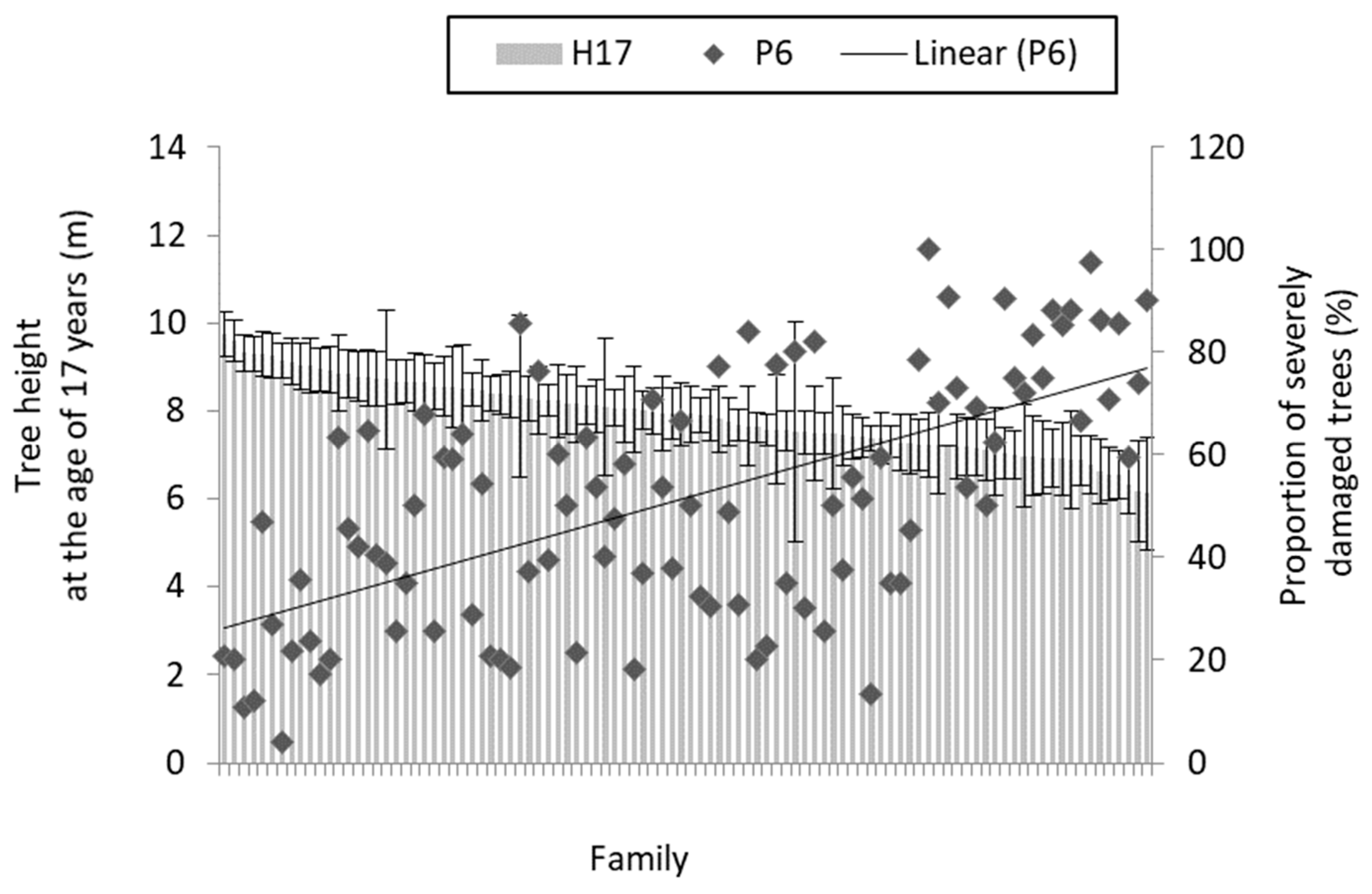
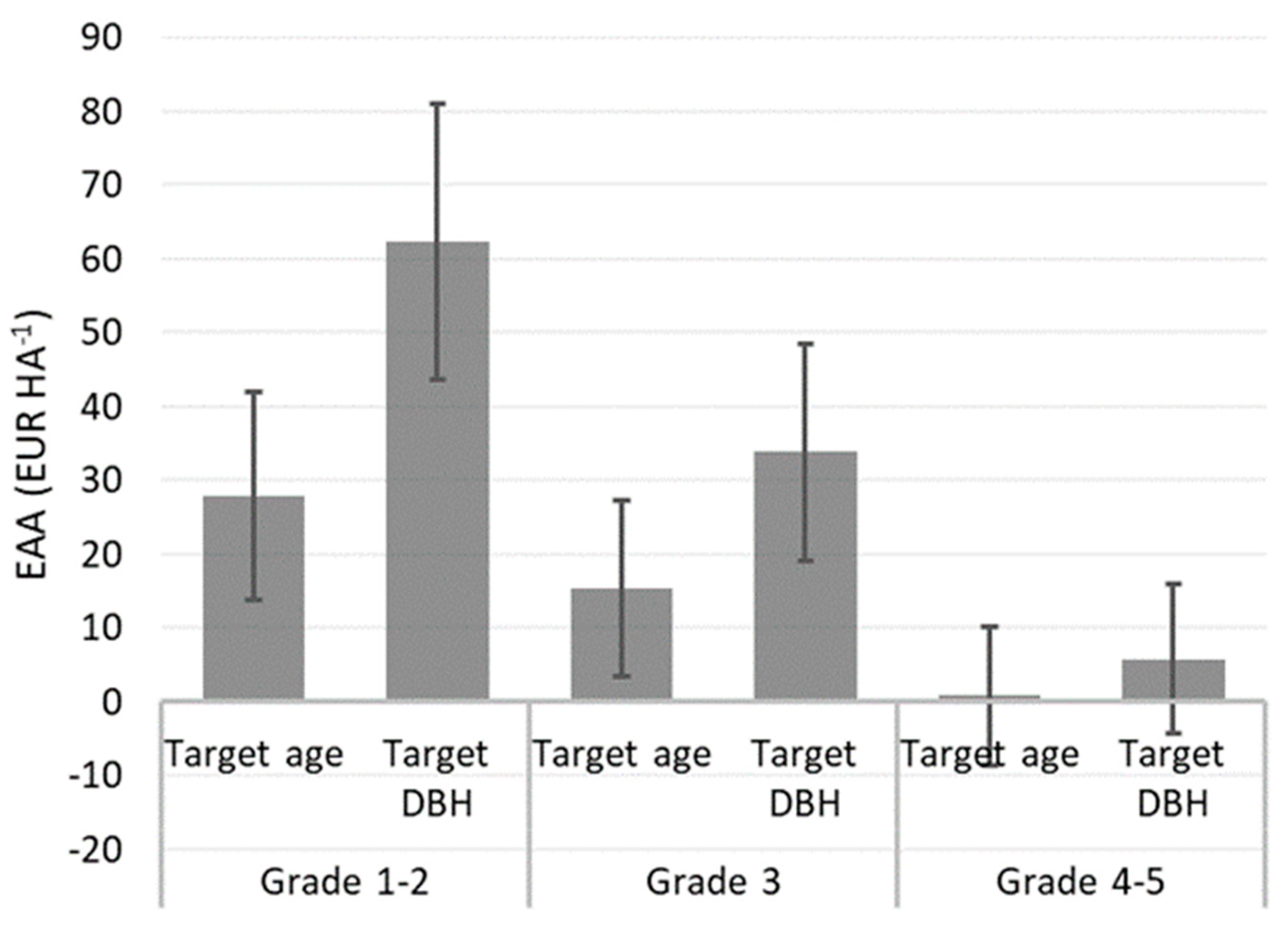
3. Results
3.1. Needle Cast Effect on Scots Pine Growth
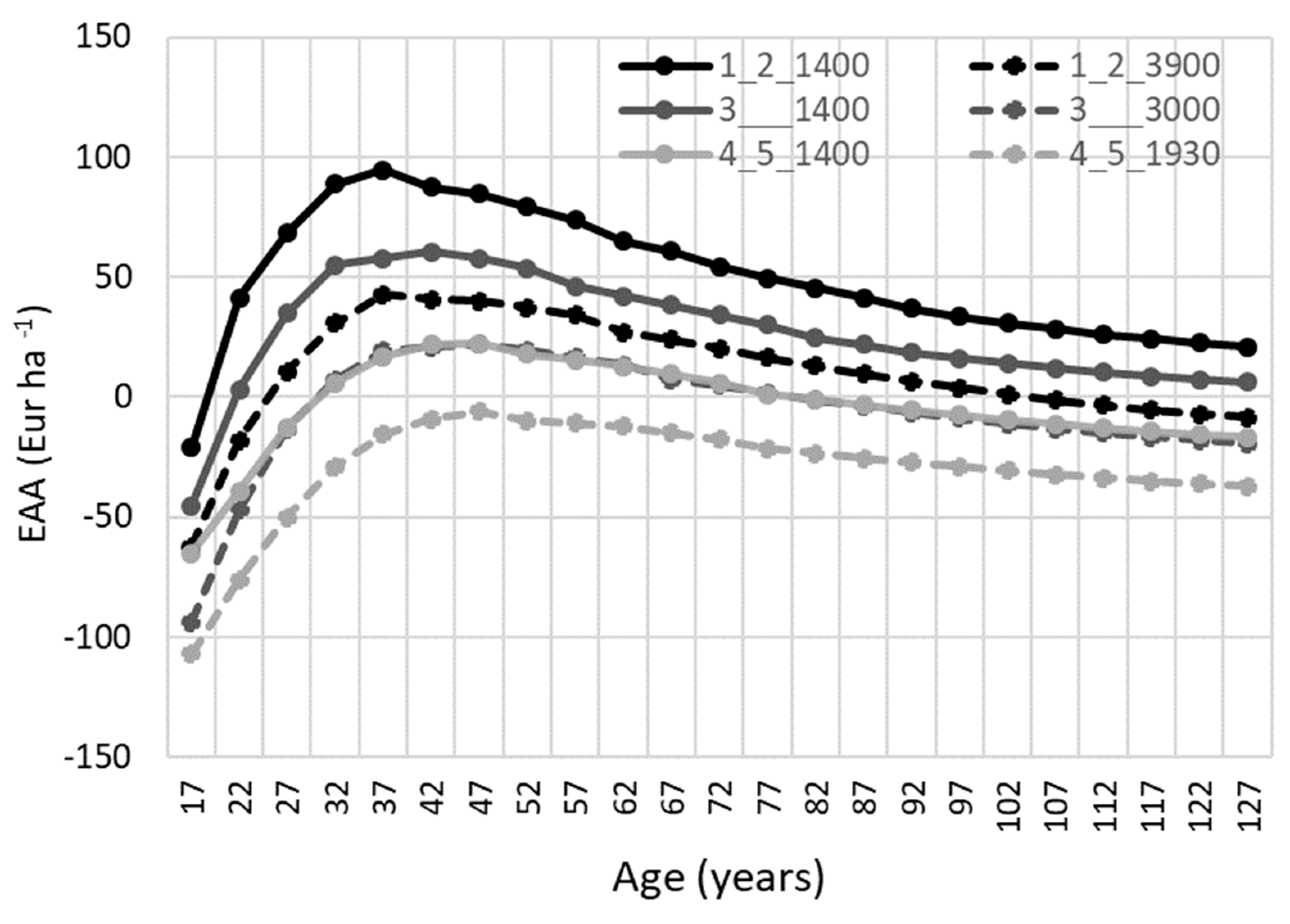
3.2. Long-Term Effects on Financial Value
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgements
Conflicts of Interest
References
- Kellomäki, S.; Peltola, H.; Nuutinen, T.; Korhonen, K.T.; Strandman, H. Sensitivity of managed boreal forests in Finland to climate change, with implications for adaptive management. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 2341–2351. [Google Scholar] [CrossRef]
- Mullett, M.S.; Brown, A.V. Effect of dothistroma needle blight on needle and shoot lengths. For. Pathol. 2018, 48, 1–7. [Google Scholar] [CrossRef]
- Bednářová, M.; Dvořák, M.; Janoušek, J.; Jankovský, L. Other foliar diseases of coniferous trees. In Infectious Forest Diseases; CABI Publishing: Wallingford, UK, 2013; pp. 458–487. ISBN 9781780640402. [Google Scholar]
- Desprez-Loustau, M.L.; Robin, C.; Reynaud, G.; Déqué, M.; Badeau, V.; Piou, D.; Husson, C.; Marçais, B. Simulating the effects of a climate-change scenario on the geographical range and activity of forest-pathogenic fungi. Can. J. Plant Pathol. 2007, 29, 101–120. [Google Scholar] [CrossRef]
- La Porta, N.; Capretti, P.; Thomsen, I.M.; Kasanen, R.; Hietala, A.M.; Von Weissenberg, K. Forest pathogens with higher damage potential due to climate change in Europe. Can. J. Plant Pathol. 2008, 30, 177–195. [Google Scholar] [CrossRef]
- Sturrock, R.N.; Frankel, S.J.; Brown, A.V.; Hennon, P.E.; Kliejunas, J.T.; Lewis, K.J.; Worrall, J.J.; Woods, Ā.J. Climate change and forest diseases. Plant Pathol. 2011, 60, 133–149. [Google Scholar] [CrossRef]
- Drenkhan, R.; Tomešová-Haataja, V.; Fraser, S.; Bradshaw, R.E.; Vahalík, P.; Mullett, M.S.; Martín-García, J.; Bulman, L.S.; Wingfield, M.J.; Kirisits, T.; et al. Global geographic distribution and host range of Dothistroma species: a comprehensive review. For. Pathol. 2016, 46, 408–442. [Google Scholar] [CrossRef]
- Wyka, S.A.; McIntire, C.D.; Smith, C.; Munck, I.A.; Rock, B.N.; Asbjornsen, H.; Broders, K.D. Effect of climatic variables on abundance and dispersal of lecanosticta acicola spores and their impact on defoliation on eastern white pine. Phytopathology 2018, 108, 374–383. [Google Scholar] [CrossRef]
- McLaughlin, J.; Šica, L. Growing pine in Latvia: Pathological considerations. Proc. Latv. Acad. Sci. 1993, 7, 49–55. [Google Scholar]
- Lilja, A.; Lilja, S.; Kurkela, T.; Rikala, R. Nursery practices and management of fungal diseases in forest nurseries in Finland. A review. Silva Fenn. 1997, 31, 79–100. [Google Scholar] [CrossRef][Green Version]
- Stenström, E.; Arvidsson, B. Fungicidal control of Lophodermium seditiosum on Pinus sylvestris seedlings in Swedish forest nurseries. Scand. J. For. Res. 2001, 16, 147–154. [Google Scholar] [CrossRef]
- Drenkhan, R.; Hanso, M. Recent invasion of foliage fungi of pines (Pinus spp.) to the Northern Baltics. For. Stud./Metsanduslikud Uurim. 2009, 51, 49–64. [Google Scholar] [CrossRef]
- Raitelaitytė, K.; Rutkauskas, A.; Radzijevskaja, J.; Žukauskienė, J.; Markovskaja, S.; Paulauskas, A. The fungal pathogens causing diseases in pines. Biologija 2017, 62. [Google Scholar] [CrossRef]
- Sierota, Z.; Grodzki, W.; Szczepkowski, A. Abiotic and biotic disturbances affecting forest health in Poland over the past 30 years: Impacts of climate and forest management. Forests 2019, 10, 75. [Google Scholar] [CrossRef]
- Diwani, S.A.; Millar, C.S. Pathogenicity of three Lophodermium species on Pinus sylvestris L. Eur. J. For. Pathol. 1987, 17, 53–58. [Google Scholar] [CrossRef]
- Ostry, M.E. Effect of Lophodermium seditiosum on Growth of Pine Nursery Seedlings in Wisconsin. Plant Dis. 1989, 73, 798. [Google Scholar] [CrossRef]
- Kulman, H. Effects of Artificial Defoliation of Pine on Subsequent Shoot and Needle Growth. For. Sci. 1965, 11, 90–98. [Google Scholar] [CrossRef]
- Jansons, Ā.; Neimane, U.; Polmanis, K.; Zaļuma, A.; Gaitnieks, T.; Baumanis, I. Cumulative effect of needle cast on scots pine saplings. For. Stud. 2016, 65, 5–15. [Google Scholar] [CrossRef]
- Montagné-Huck, C.; Brunette, M. Economic analysis of natural forest disturbances: A century of research. J. For. Econ. 2018, 32, 42–71. [Google Scholar] [CrossRef]
- Hanso, M.; Drenkhan, R. Lophodermium needle cast, insect defoliation and growth responses of young Scots pines in Estonia. For. Pathol. 2012, 42, 124–135. [Google Scholar] [CrossRef]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manage. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Macpherson, M.F.; Kleczkowski, A.; Healey, J.R.; Hanley, N. The effects of disease on optimal forest rotation: A generalisable analytical framework. Environ. Resour. Econ. 2016, 70, 565–588. [Google Scholar] [CrossRef]
- Hanso, M.; Drenkham, R. Retrospective Analysis of Lophodermium seditiosum Epidemics in Estonia. Acta Silv. LIgn. Hung. 2007, Spec. Edit, 31–45. [Google Scholar]
- Kanaskie, A. Lophodermium needle cast of Scotch pine. In Growing Healthy Seedlings: Identification and Management of Pests in Northwest Forest Nurseries; Hamm, P.B., Hansen, E.M., Campbell, S.J., Eds.; Forest Pest Management, U.S. Department of Agriculture, Forest Service, Pacific Northwest Region; and Forest Research Laboratory, College of Forestry, Oregon State University: Corvallis, OR, USA, 1990; p. 34. [Google Scholar]
- Berlin, M.; Lönnstedt, L.; Jansson, G.; Danell, Ö.; Ericsson, T. Developing a Scots pine breeding objective: A case study involving a Swedish sawmill. Silva Fenn. 2010, 44, 643–656. [Google Scholar] [CrossRef]
- Krakau, U.-K.; Liesebach, M.; Aronen, T.; Lelu-Walter, M.-A.; Schneck, V. Scots Pine (Pinus sylvestris L.); Springer: Dordrecht, The Netherlands, 2013; pp. 267–323. [Google Scholar]
- Suontama, M.; Li, Y.; Low, C.B.; Dungey, H.S. Genetic improvement of resistance to cyclaneusma needle cast in pinus radiata. Can. J. For. Res. 2019, 49, 128–133. [Google Scholar] [CrossRef]
- Barzdajn, W.; Kowalkowski, W.; Chmura, D.J. Variation in growth and survival among European provenances of pinus sylvestris in a 30-year-old experiment. Dendrobiology 2016, 75, 67–77. [Google Scholar] [CrossRef][Green Version]
- Rieksts-Riekstiņš, R.; Zeltiņš, P.; Baliuckas, V.; Bruna, L.; Zaluma, A.; Kapostiņš, R. Pinus sylvestris breeding for resistance against natural infection of the fungus heterobasidion annosum. Forests 2020, 11, 23. [Google Scholar] [CrossRef]
- Squillace, A. Genetic Variation and Breeding of Scots Pine in the Netherlands. For. Sci. 1975, 21, 341–352. [Google Scholar] [CrossRef]
- Neimane, U.; Polmanis, K.; Zaluma, A.; Klavina, D.; Gaitnieks, T.; Jansons, A. Damage caused by Lophodermium needle cast in open-pollinated and control-crossed progeny trials of Scots pine (Pinus sylvestris L.). For. Chron. 2018, 94, 155–161. [Google Scholar] [CrossRef]
- Stephan, B.R.; Krusche, D. Genetic Variation of Resistance to Lophodermium Needle Cast in Scots Pine Progenies of Intraprovenance and Interprovenance Crossings1. In Proceedings of the Recent Research on Conifer Needle Diseases: Conference Proceedings, Gulfport, MS, USA, 14–18 October 1984; Petterson, G.W., Ed.; US Department of Agriculture, Forest Service: Washington, DC, USA, 1986. [Google Scholar]
- Ranta, H.; Pulkkinen, P.; Neuvonen, S. Susceptibility of six scots pine clones to the pathogenic fungus gremmeniella abietina. Scand. J. For. Res. 2000, 15, 7–12. [Google Scholar] [CrossRef]
- Schelhaas, M.J.; Nabuurs, G.J.; Schuck, A. Natural disturbances in the European forests in the 19th and 20th centuries. Glob. Chang. Biol. 2003, 9, 1620–1633. [Google Scholar] [CrossRef]
- Jactel, H.; Nicoll, B.C.; Branco, M.; Gonzalez-Olabarria, J.R.; Grodzki, W.; Långström, B.; Moreira, F.; Netherer, S.; Christophe Orazio, C.; Piou, D.; et al. The influences of forest stand management on biotic and abiotic risks of damage. Ann. For. Sci. 2009, 66. [Google Scholar] [CrossRef]
- Seidl, R.; Fernandes, P.M.; Fonseca, T.F.; Gillet, F.; Jönsson, A.M.; Merganičová, K.; Netherer, S.; Arpaci, A.; Bontemps, J.D.; Bugmann, H.; et al. Modelling natural disturbances in forest ecosystems: A review. Ecol. Modell. 2011, 222, 903–924. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.J.; Rammer, W.; Verkerk, P.J. Increasing forest disturbances in Europe and their impact on carbon storage. Nat. Clim. Chang. 2014, 4, 806–810. [Google Scholar] [CrossRef] [PubMed]
- Thürig, E.; Hagedorn, F.; Lindroth, A. Influence of storm damage on the forest carbon balance. In Living with the Storm Damage; Gardiner, E., Schuck, A., Schelhaas, M.-J., Orazio, C., Blennow, K., Nicoll, B., Eds.; European Forest institute: Joensuu, Finland, 2013; pp. 47–55. [Google Scholar]
- Zubizarreta-Gerendiain, A.; Pukkala, T.; Peltola, H. Effects of wind damage on the optimal management of boreal forests under current and changing climatic conditions. Can. J. For. Res. 2017, 47, 246–256. [Google Scholar] [CrossRef]
- Bušs, K. Meža ekoloģija un tipoloģija [Forest Ecology and Typology]; Zinātne: Rīga, Latvia, 1981. [Google Scholar]
- R Core Team R: A language and environment for statistical computing. 2019. Available online: https://www.gbif.org/tool/81287/r-a-language-and-environment-for-statistical-computing (accessed on 22 January 2020).
- Donis, J.; Šņepsts, G. Novēloti koptu vienvecuma egļu audžu apsaimniekošanas alternatīvas un to ekonomisks izvērtējums [Management alternatives and their financial analysis for even-age Norway spruce stands with delayed thinnings]. In Vienvecuma egļu meži Latvijā; Jansons, J., Ed.; LVMI Silava: Salaspils, Latvia, 2019; pp. 71–98. ISBN 978-9984-14-853-3. [Google Scholar]
- Ozolins, R. Forest stand assortment structure analysis using mathematical modelling. For. Stud. Uurim. 2002, 37, 33–42. [Google Scholar]
- Central Statistical Bureau of Latvia: Average purchase prices of wood. 2018. Available online: https://www.csb.gov.lv/en/statistics/statistics-by-theme/agriculture/forestry/tables/meg050/average-purchase-prices-wood-eur-m3-excluding-vat (accessed on 17 December 2019).
- Klempered, W.D. Forest Resource Economics and Finance; McGraw-Hill Series in Forest Resources: Blacksburg, VA, USA, 1996. [Google Scholar]
- Stenström, E.; Ihrmark, K. Identification of Lophodermium seditiosum and L. pinastri in Swedish forest nurseries using species-specific PCR primers from the ribosomal ITS region. For. Pathol. 2005, 35, 163–172. [Google Scholar] [CrossRef]
- Martinsson, O. Testing Scots Pine for Resistance to Lophodermium Needle Cast. Stud. For. Suec. 1979, 63. [Google Scholar]
- Garrett, K.A.; Nita, M.; De Wolf, E.D.; Esker, P.D.; Gomez-Montano, L.; Sparks, A.H. Plant Pathogens as Indicators of Climate Change. In Climate Change: Observed Impacts on Planet Earth: Second Edition; Elsevier Inc.: Amsterdam, The Netherlands, 2016; pp. 325–338. ISBN 9780444635242. [Google Scholar]
- Kurkela, T.; Drenkhan, R.; Vuorinen, M.; Hanso, M. Growth response of young Scots pines to needle loss assessed from productive foliage. For. Stud./Metsanduslikud Uurim. 2009, 50, 5–22. [Google Scholar] [CrossRef]
- Donis, J.; Saleniece, R.; Krisans, O.; Dubrovskis, E.; Kitenberga, M.; Jansons, A. A Financial Assessment of Windstorm Risks for Scots Pine Stands in Hemiboreal Forests. Forests 2020, 11, 566. [Google Scholar] [CrossRef]
- Samariks, V.; Krisans, O.; Donis, J.; Silamikele, I.; Katrevics, J.; Jansons, A. A cost-benefit analysis of measures to reduce windstorm impact in Norway spruce (Picea abies L. Karst.) stands: A case study in Latvia. Forests 2020, 11, 576. [Google Scholar] [CrossRef]
- Kowalski, T. Interrelationships among Lophodermium seditiosum, L. pinastri and Cyclaneusma minus in pine plantations (Pinus sylvestris L.) in Poland. In Proceedings of the Recent Research on Foliage Diseases, Carlisle, PA, USA, 29 May–2 June 1989; Merrill, W., Ostry, M.E., Eds.; USDA: Washington, DC, USA, 1990; pp. 13–18. [Google Scholar]
- Kimberley, M.O.; Hood, I.A.; Knowles, R.L. Impact of Swiss needle-cast on growth of Douglas-fir. Phytopathology 2011, 101, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Brawner, J.T.; Carter, D.R.; Huber, D.A.; White, T.L. Projected gains in rotation-age volume and value from fusiform rust resistant slash and loblolly pines. Can. J. For. Res. 1999, 29, 737–742. [Google Scholar] [CrossRef]
- Ivković, M.; Baltunis, B.; Gapare, W.; Sasse, J.; Dutkowski, G.; Elms, S.; Wu, H. Breeding against dothistroma needle blight of radiata pine in Australia. Can. J. For. Res. 2010, 40, 1653–1660. [Google Scholar] [CrossRef]
- Petucco, C.; Andrés-Domenech, P. Land expectation value and optimal rotation age of maritime pine plantations under multiple risks. J. For. Econ. 2018, 30, 58–70. [Google Scholar] [CrossRef]
- Mäkinen, H.; Hynynen, J.; Isomäki, A. Intensive management of Scots pine stands in southern Finland: First empirical results and simulated further development. For. Ecol. Manag. 2005, 215, 37–50. [Google Scholar] [CrossRef]
- Mäkinen, H.; Isomäki, A. Thinning intensity and growth of Scots pine stands in Finland. For. Ecol. Manag. 2004, 201, 311–325. [Google Scholar] [CrossRef]
- Randolph, K.C. Status of fusiform rust incidence in slash and loblolly pine plantations in the southeastern United States. Gen. Tech. Rep. South. Res. Station. USDA For. Serv. 2016, 212, 53–59. [Google Scholar]
- Susaeta, A. Implications of future risk of fusiform rust on optimal forest management of even-aged slash pine plantations. For. Pol. Econ. 2020, 116, 102183. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Correlation Coefficient with the Proportion of Severely Damaged Trees 1 | Range of Family Mean Values (± 95% Confidence Interval) | |
|---|---|---|---|
| Minimum | Maximum | ||
| Survival at the age of 5 years (%) | −0.07 | 43 | 100 |
| Survival at the age of 17 years relative to that at age 5 (%) | −0.70 *** | 6 | 96 |
| Height at the age of 5 years (cm) | −0.25 * | 103 ± 10.1 | 157 ± 13.4 |
| Diameter of the root collar at the age of 5 years (mm) | −0.37 *** | 20 ± 1.8 | 32 ± 2.2 |
| Height at the age of 17 years (m) | −0.62 *** | 6 ± 1.3 | 10 ± 0.5 |
| Stem diameter at the breast height at the age of 17 years (cm) | −0.40 *** | 7 ± 1.3 | 12 ± 2.9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jansons, Ā.; Zeltiņš, P.; Donis, J.; Neimane, U. Long-Term Effect of Lophodermium Needle Cast on The Growth of Scots Pine and Implications for Financial Outcomes. Forests 2020, 11, 718. https://doi.org/10.3390/f11070718
Jansons Ā, Zeltiņš P, Donis J, Neimane U. Long-Term Effect of Lophodermium Needle Cast on The Growth of Scots Pine and Implications for Financial Outcomes. Forests. 2020; 11(7):718. https://doi.org/10.3390/f11070718
Chicago/Turabian StyleJansons, Āris, Pauls Zeltiņš, Jānis Donis, and Una Neimane. 2020. "Long-Term Effect of Lophodermium Needle Cast on The Growth of Scots Pine and Implications for Financial Outcomes" Forests 11, no. 7: 718. https://doi.org/10.3390/f11070718
APA StyleJansons, Ā., Zeltiņš, P., Donis, J., & Neimane, U. (2020). Long-Term Effect of Lophodermium Needle Cast on The Growth of Scots Pine and Implications for Financial Outcomes. Forests, 11(7), 718. https://doi.org/10.3390/f11070718