Effectiveness of Restoration Treatments for Reducing Fuels and Increasing Understory Diversity in Shrubby Mixed-Conifer Forests of the Southern Rocky Mountains, USA



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design and Field Methods
2.3. Statistical Analysis
3. Results
3.1. Pretreatment Conditions
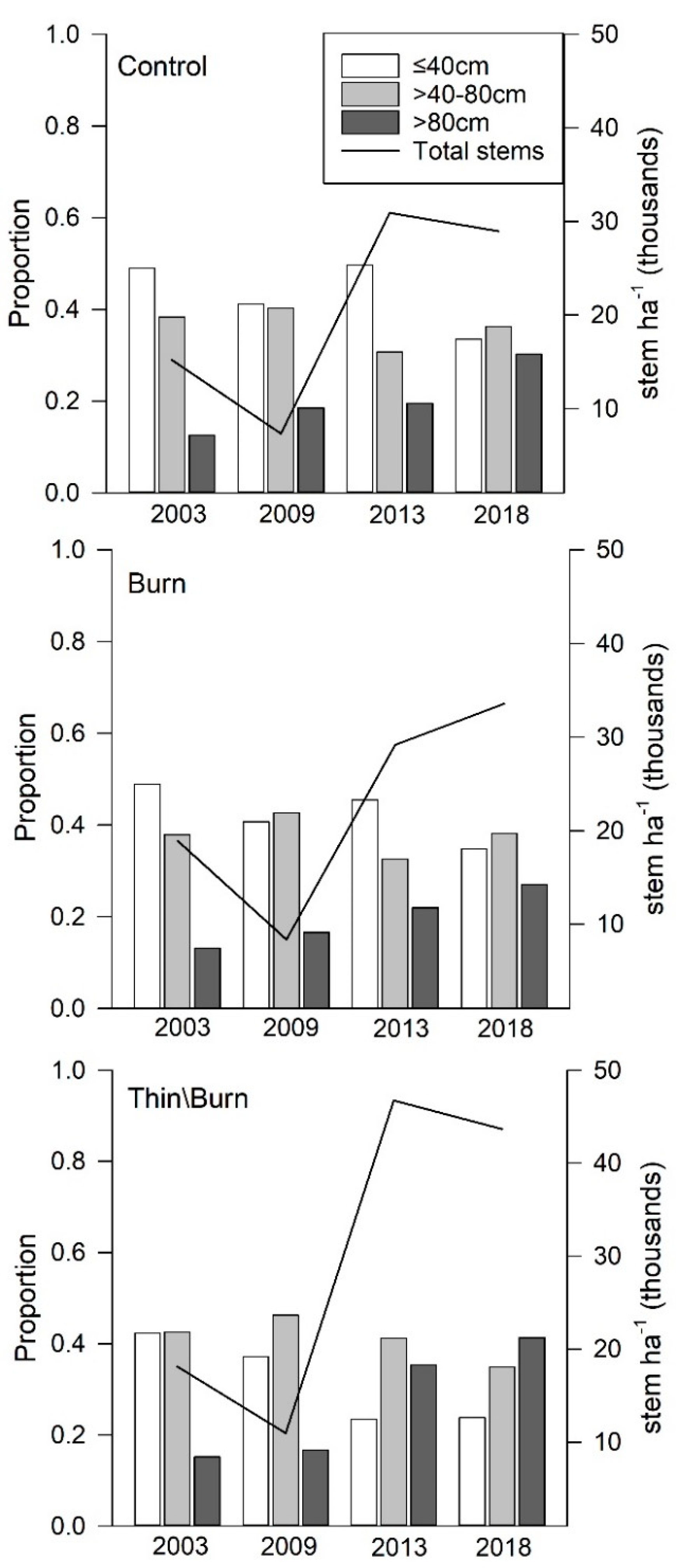
3.2. Overstory Structure
3.3. Surface and Canopy Fuels
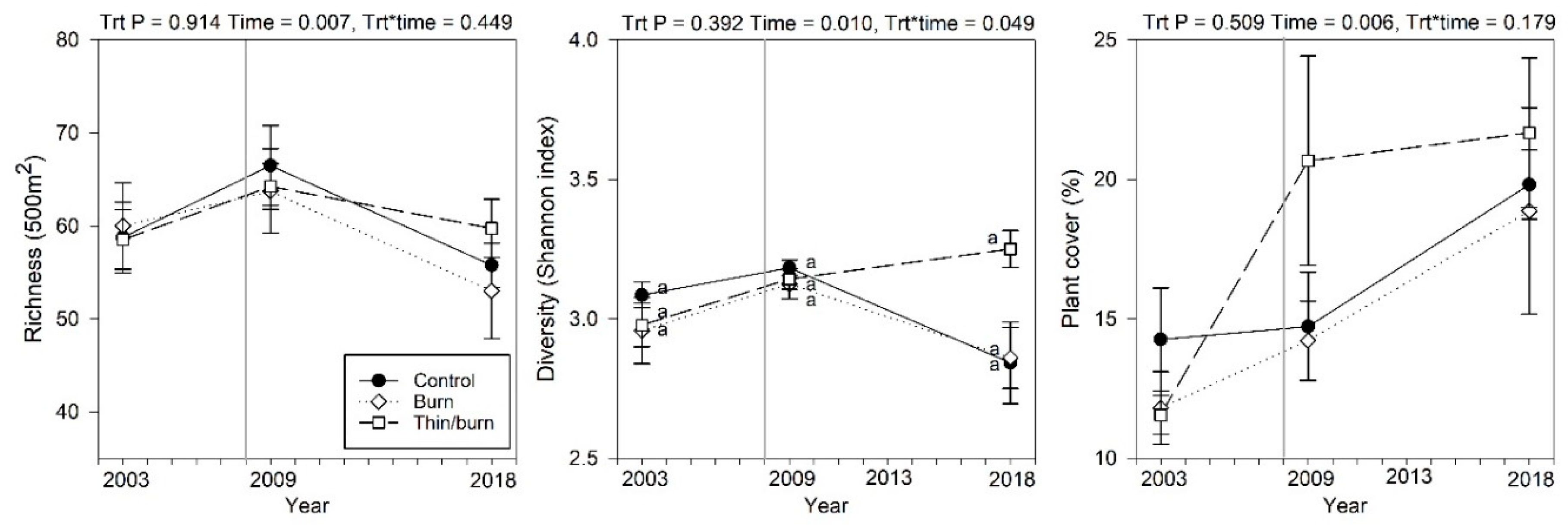
3.4. Understory Community Dynamics
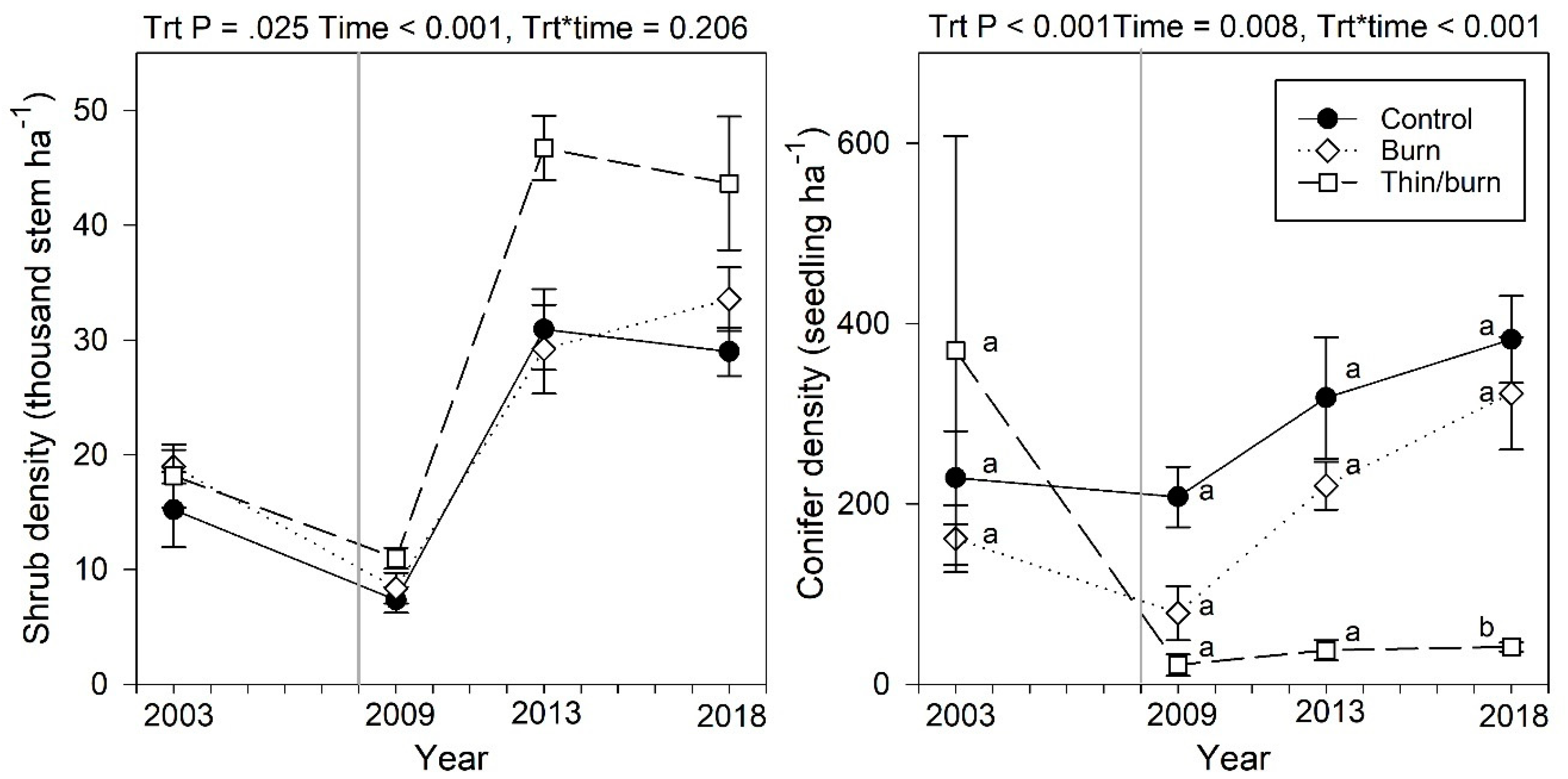
3.5. Midstory Shrubs and Conifer Regeneration
4. Discussion
4.1. Overstory Dynamics
4.2. Treatment Effects on Understory Vegetation
4.3. Future Resiliency
5. Conclusions and Management Implications
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Romme, W.H.; Floyd, M.L.; Hanna, D. Historical range of variability and current landscape condition analysis: South Central Highlands section, Southwestern Colorado and Northwestern New Mexico. Col. For. Rest. Inst. 2009, 2–256. [Google Scholar]
- Schoennagel, T.; Balch, J.K.; Brenkert-Smith, H.; Dennison, P.E.; Harvey, B.J.; Krawchuk, M.A.; Mietkiewicz, N.; Morgan, P.; Moritz, M.A.; Rasker, R.; et al. Adapt to more wildfire in western North American forests as climate changes. Proc. Natl. Acad. Sci. USA 2017, 114, 4582–4590. [Google Scholar] [CrossRef] [PubMed]
- Singleton, M.P.; Thode, A.E.; Sánchez Meador, A.J.; Iniguez, J.M. Increasing trends in high-severity fire in the southwestern USA from 1984 to 2015. For. Ecol. Manag. 2019, 433, 709–719. [Google Scholar] [CrossRef]
- Reynolds, R.T.; Sánchez Meador, A.J.; Youtz, J.A.; Nicolet, T.; Matonis, M.S.; Jackson, P.L.; DeLorenzo, D.G. Graves AD Restoring Composition and Structure in Southwestern Frequent-Fire Forests: A Science-Based Framework for Improving Ecosystem Resiliency; General Technical Report RMRS-GTR-310; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2013; p. 76.
- Heinlein, T.A.; Moore, M.M.; Fulé, P.Z.; Covington, W.W. Fire history of two ponderosa pine—Mixed conifer sites: San Francisco Peaks, Arizona, USA. Int. J. Wildland Fire 2005, 14, 307–320. [Google Scholar] [CrossRef]
- Fulé, P.Z.; Korb, J.E.; Wu, R. Changes in forest structure of a dry mixed conifer forest, southwestern Colorado USA. For. Ecol. Manag. 2009, 258, 1200–1210. [Google Scholar] [CrossRef]
- Huffman, D.W.; Zegler, T.J.; Fule, P.Z. Fire history of a mixed conifer forest on the Mogollon Rim, northern Arizona USA. Int. J. Wildland Fire 2015, 24, 680–689. [Google Scholar] [CrossRef]
- Korb, J.E.; Daniels, M.L.; Laughlin, D.C.; Fule, P.Z. Understory communities of warm-dry, mixed-conifer forests in southwestern Colorado. Southwest. Nat. 2007, 52, 493–503. [Google Scholar] [CrossRef]
- Evans, A.M.; Everett, R.G.; Stephens, S.L.; Youlz, J.A. Comprehensive Fuels Treatment Practices Guide for Mixed Conifer Forests: California, Central and Southern Rockies, and the Southwest; JFSP Synthesis Reports; Paper 12; 2011. Available online: http://digitalcommons.unl.edu/jfspsynthesis/12 (accessed on 10 December 2019).
- Strahan, R.T.; Sánchez Meador, A.J.; Huffman, D.W.; Laughlin, D.C. Shifts in community-level traits and functional diversity in a mixed conifer forest: A legacy of land-use change. J. Appl. Ecol. 2016, 53, 1755–1765. [Google Scholar] [CrossRef]
- Goodwin, M.J.; North, M.P.; Zald, H.S.; Hurteau, M.D. The 15-year post-treatment response of a mixed-conifer understory plant community to thinning and burning treatments. For. Ecol. Manag. 2018, 429, 617–624. [Google Scholar] [CrossRef]
- Churchill, D.J.; Larson, A.J.; Dahlgreen, M.C.; Franklin, J.F.; Hessburg, P.F.; Lutz, J.A. Restoring forest resilience: From reference spatial patterns to silvicultural prescriptions and monitoring. For. Ecol. Manag. 2013, 291, 442–457. [Google Scholar] [CrossRef]
- Larson, A.J.; Belote, A.T.; Cansler, C.A.; Parks, S.A.; Dietz, M.S. Latent resilience in ponderosa pine forest: Effects of resumed frequent fire. Ecol. Appl. 2013, 23, 1243–1249. [Google Scholar] [CrossRef] [PubMed]
- Hessburg, P.F.; Miller, C.L.; Parks, S.A.; Povak, N.A.; Taylor, A.H.; Higuera, P.E.; Prichard, S.J.; North, M.P.; Collins, B.M.; Hurteau, M.D.; et al. Climate, environment, and disturbance history govern resilience of western North American forests. Front. Ecol. Evol. 2019, 7, 1–27. [Google Scholar] [CrossRef]
- Rossman, A.K.; Halpern, C.B.; Harrod, R.J.; Urgenson, L.S.; Peterson, D.W.; Bakker, J.D. Benefits of thinning and burning for understory diversity vary with spatial scale and time since treatment. For. Ecol. Manag. 2018, 419–420, 58–78. [Google Scholar] [CrossRef]
- Dodson, E.K.; Metlen, K.L.; Fiedler, C.E. Common and uncommon understory species differentially respond to restoration treatments in ponderosa pine/Douglas-fir forests, Montana. Restor. Ecol. 2007, 15, 692–704. [Google Scholar] [CrossRef]
- Metlen, K.L.; Fiedler, C.E. Restoration treatment effects on the understory of ponderosa pine/Douglas-fir forests in western Montana, USA. For. Ecol. Manag. 2006, 222, 355–369. [Google Scholar] [CrossRef]
- Wayman, R.; North, M. Initial response of a mixed-conifer understory plant community to burning and thinning restoration treatments. For. Ecol. Manag. 2007, 239, 32–44. [Google Scholar] [CrossRef]
- Dodson, E.K.; Peterson, D.W.; Harrod, R. Understory vegetation response to thinning and burning restoration treatments in dry conifer forests of the eastern Cascades, USA. For. Ecol. Manag. 2008, 255, 3130–3140. [Google Scholar] [CrossRef]
- Abella, S.R.; Springer, J.D. Effects of tree cutting and fire on understory vegetation in mixed-conifer forests. For. Ecol. Manag. 2015, 335, 281–299. [Google Scholar] [CrossRef]
- Willms, J.; Bartuszevige, A.; Schwilk, D.W.; Kennedy, P.L. The effects of thinning and burning on understory vegetation in North America: A meta-analysis. For. Ecol. Manag. 2017, 392, 184–194. [Google Scholar] [CrossRef]
- Springer, S.D.; Huffman, D.W.; Stoddard, M.T.; Sánchez Meador, A.J.; Waltz, A.E.M. Plant community dynamics following hazardous fuel treatments and mega-wildfire in a warm-dry mixed-conifer forest of the USA. For. Ecol. Manag. 2018, 429, 278–286. [Google Scholar] [CrossRef]
- Schwilk, D.W.; Keeley, J.E.; Knapp, E.E.; McIver, J.; Bailey, J.D.; Fettig, C.J.; Fiedler, C.E.; Harrod, R.J.; Moghaddas, J.J.; Outcalt, K.W.; et al. The national Fire and Fire Surrogate study: Effects of fuel reduction methods on forest vegetation structure and fuels. Ecol. Appl. 2009, 19, 285–304. [Google Scholar] [CrossRef] [PubMed]
- Savage, M.; Mast, J.N. How resilient are southwestern ponderosa pine forests after crown fires? Can. J. For. Res. 2005, 35, 967–977. [Google Scholar] [CrossRef]
- Battaglia, M.A.; Smith, F.W.; Shepperd, W.D. Can prescriped fire be used to maintain fuel treatment effectiveness over time in Black Hills ponderosa pine forests? For. Ecol. Manag. 2008, 256, 2029–2038. [Google Scholar] [CrossRef]
- Stoddard, M.T.; Sánchez Meador, A.J.; Fulé, P.Z.; Korb, J.E. Five-year post restoration conditions and simulated climate-change trajectories in a warm/ dry mixed-conifer forest, southwestern Colorado, USA. For. Ecol. Manag. 2015, 356, 253–261. [Google Scholar] [CrossRef]
- Vaillant, N.M.; Noonan-Wright, E.K.; Reiner, A.L.; Ewel, C.M.; Rau, B.M.; Fites-Kaufman, J.A.; Dailey, S.N. Fuel accumulation and forest structure change following hazardous fuel reduction treatments throughout California. Int. J. Wildland Fire 2015, 24, 361–371. [Google Scholar] [CrossRef]
- Reinhardt, E.D.; Keane, R.E.; Calkin, D.E.; Cohen, J.D. Objectives and considerations for wildland fuel treatment in forested ecosystems of the interior western United States Forest. For. Ecol. Manag. 2008, 256, 1997–2006. [Google Scholar] [CrossRef]
- US Department of Agriculture, Forest Service. Lower Middle Mountain Research Project Environmental Assessment; US Department of Agriculture, Forest Service, San Juan National Forest: Durango, CO, USA, 2004.
- Western Regional Climate Center. Available online: https://wrcc.dri.edu/cgi-bin/cliMAIN.pl?co8582 (accessed on 10 November 2019).
- Korb, J.E.; Fulé, P.Z.; Stoddard, M.T. Forest restoration in a surface fire dependent ecosystem: An example from a mixed-conifer forest, southwestern Colorado, USA. For. Ecol. Manag. 2012, 269, 10–11. [Google Scholar] [CrossRef]
- Thomas, J.W.; Anderson, R.G.; Maser, C.; Bull, E.L. Snags. In Wildlife Habitats in Managed Forests—The Blue Mountains of Oregon and Washington; USDA Agricultural Handbook 553: Washington, DC, USA, 1979; pp. 60–77. [Google Scholar]
- Brown, J.K. Handbook for Inventorying Downed Woody Material; General Technical Report INT-16; US Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1974.
- Stohlgren, T.J.; Falkner, M.B.; Schell, L.D. A Modified-Whittaker nested vegetation sampling method. Vegetatio 1995, 117, 113–121. [Google Scholar] [CrossRef]
- Roccaforte, J.P.; Huffman, D.W.; Fulé, P.Z.; Covington, W.W.; Chancellor, W.W.; Stoddard, M.T.; Crouse, J.E. Forest structure and fuels dynamic following ponderosa pine restoration treatments, White Mountain, Arizona, USA. For. Ecol. Manag. 2015, 337, 174–185. [Google Scholar] [CrossRef]
- Ffolliot, P.F.; Clay, W.P.; Davis, J.R. Some Characteristics of the Forest Floor under Ponderosa Pine in Arizona; Research Note, RM-127; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ft Collins, CO, USA, 1968.
- Spellerberg, I.F.; Fedor, P.J. A tribute to Claude Shannon (1916–2001) and a plea for more rigorous use of species richness, species diversity and the “Shannon-Weiner” Index. Global Ecol. Biogeogr. 2003, 12, 177–179. [Google Scholar] [CrossRef]
- McCune, B.; Grace, J.B. Analysis of Ecological Communities; MjM Software Design: Gleneden, ON, Canada, 2002. [Google Scholar]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analysis of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- McArdle, B.H.; Anderson, M.J. Fitting multivariate models to community data: A comment on distance-based redundancy analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- McCune, B.; Mefford, M.J. Multivariate Analysis of Ecological Data, PCORD Version 5.10; MjM Software: Gleneden Beach, ON, Canada, 2006. [Google Scholar]
- Strom, B.A.; Fule’, P.Z. Pre-wildfire fuel treatments affect long-term ponderosa pine forest dynamics. Int. J. Wildland Fire 2007, 16, 128–138. [Google Scholar] [CrossRef]
- Fulé, P.Z.; Crouse, J.E.; Roccaforte, J.P.; Kalies, E.L. Do thinning and/or burning treatments in western USA ponderosa or Jeffrey pine dominated forests help restore natural fire behavior? For. Ecol. Manag. 2012, 269, 68–81. [Google Scholar] [CrossRef]
- Stephens, S.L.; Collins, B.M.; Roller, G. Fuel treatment longevity in a Sierra Nevada mixed conifer forest. For. Ecol. Manag. 2012, 285, 204–212. [Google Scholar] [CrossRef]
- Ziegler, J.P.; Hoffman, C.; Battaglia, M.; Mell, W. Spatially explicit measurements of forest structure and fire behavior following restoration treatments in dry forests. For. Ecol. Manag. 2017, 386, 1–12. [Google Scholar] [CrossRef]
- Cannon, J.B.; Barretta, K.J.; Gannona, B.M.; Addington, R.N.; Battaglia, M.A.; Fornwalt, P.J.; Aplet, G.H.; Chenga, A.C.; Underhill, J.L.; Briggs, J.S.; et al. Collaborative restoration effects on forest structure in ponderosa pine dominated forests of Colorado. For. Ecol. Manag. 2018, 424, 191–204. [Google Scholar] [CrossRef]
- Fulé, P.Z.; Waltz, A.E.M.; Covington, W.W.; Heinlein, T.A. Measuring forest restoration effectiveness in reducing hazardous fuels. J. Forest. 2001, 99, 24–29. [Google Scholar]
- Stephens, S.L.; Moghaddas, J.J. Experimental fuel treatment impacts on forest structure, potential fire behavior, and predicted tree mortality in a mixed conifer forest. For. Ecol. Manag. 2005, 215, 21–36. [Google Scholar] [CrossRef]
- Stephens, S.L.; Moghaddas, J.J.; Ediminster, C.; Fiedler, C.E.; Hasse, S.; Harrington, M.; Keeley, J.E.; Knapp, E.E.; McIver, J.D.; Metlen, K.; et al. Fire treatment effects on vegetation structure, fuels, and potential fire severity in western US forests. Ecol. Appl. 2009, 19, 305–320. [Google Scholar] [CrossRef] [PubMed]
- Fieldler, C.E.; Metlen, K.L.; Dodson, E.K. Restoration treatment effects on stand structure, tree growth, and fire hazard in a ponderosa pine/Douglas-fir forest in Montana. For. Sci. 2010, 56, 18–31. [Google Scholar]
- Dickinson, Y. Landscape restoration of a forest with a historically mixed-severity fire regime: What was the historical landscape pattern of forest and openings? For. Ecol. Manag. 2014, 331, 264–271. [Google Scholar] [CrossRef]
- Brown, P.M.; Battaglia, M.A.; Fornwalt, P.J.; Gannon, B.; Huckaby, L.S.; Julian, C.; Cheng, A.S. Historical (1860) forest structure in ponderosa pine forests of the northern Front Range, Colorado. Can. J. For. Res. 2015, 45, 1462–1473. [Google Scholar] [CrossRef]
- Allen, C.D.; Savage, M.; Falk, D.; Suckling, K.F.; Swetnam, T.W.; Schulke, T.; Stacey, P.B.; Morgan, P.; Hoffman, M.; Klingel, J.T. Ecological restoration of southwestern ponderosa pine ecosystems: A broad perspective. Ecol. Appl. 2002, 12, 1418–1433. [Google Scholar] [CrossRef]
- Battaglia, M.A.; Gannon, B.; Brown, P.M.; Fornwalt, P.J.; Cheng, A.S.; Huckaby, L.S. Changes in forest structure since 1860 in ponderosa pine dominated forests in the Colorado and Wyoming Front Range, USA. For. Ecol. Manag. 2018, 422, 147–160. [Google Scholar] [CrossRef]
- Korb, J.E.; Johnson, N.C.; Covington, W.W. Arbuscular mycorrhizal propagule densities respond rapidly to ponderosa pine restoration treatments. J. Appl. Ecol. 2003, 40, 101–110. [Google Scholar] [CrossRef]
- Nelson, C.R.; Halpern, C.B.; Agee, J.K. Thinning and burning result in low-level invasion by nonnative plants but neutral effects on natives. Ecol. Appl. 2008, 18, 762–770. [Google Scholar] [CrossRef]
- Bartuszevige, A.M.; Kennedy, P.L. Synthesis of Knowledge on the Effects of Fire and Thinning Treatments on Understory Vegetation in U.S. Dry Forests. Special Report 1095; Oregon State Agricultural Experiment Station, Oregon State University, Agricultural Experiment Station: Corvallis, OR, USA, 2009. [Google Scholar]
- Kerns, B.K.; Day, M.A. Prescribed fire regimes subtly alter ponderosa pine forest plant community structure. Ecosphere 2018, 9, 1–20. [Google Scholar] [CrossRef]
- Strahan, R.T.; Stoddard, M.T.; Springer, J.D.; Huffman, D.W. Increasing weight of evidence that thinning and burning treatments help restore understory plant communities in ponderosa pine forests. For. Ecol. Manag. 2015, 353, 208–220. [Google Scholar] [CrossRef]
- Moore, M.M.; Casey, C.A.; Bakker, J.D.; Springer, J.D. Herbaceous vegetation responses (1992–2004) to restoration treatments in a ponderosa pine forest. Rangel. Ecol. Manag. 2006, 59, 135–144. [Google Scholar] [CrossRef]
- Korb, J.E.; Covington, W.W.; Fulé, P.Z. Sampling techniques influence understory plant trajectories after restoration: An example from ponderosa pine restoration. Restor. Ecol. 2003, 11, 504–515. [Google Scholar] [CrossRef]
- Abella, S.R.; Covington, W.W. Monitoring an Arizona ponderosa pine restoration: Sampling efficiency and multivariate analysis of understory vegetation. Restor. Ecol. 2004, 12, 359–367. [Google Scholar] [CrossRef]
- Munson, S.M. Plant responses, climate pivot points, and trade-offs in water limited ecosystems. Ecosphere 2013, 4, 109. [Google Scholar] [CrossRef]
- MacDonald, G.M. Water, climate change, and sustainability in the southwest. Proc. Natl. Acad. Sci. USA. 2010, 107, 21256–21262. [Google Scholar] [CrossRef] [PubMed]
- Stoddard, M.T.; McGlone, C.M. Effects of ecological restoration alternative treatments on nonnative plant species establishment. In Fort Valley Experimental Forest—A Century of Research 1908–2008, Proceedings of the RMRS-P-55, Fort Collins, CO, USA, 7–9 August 2008; Olberding, S.D., Moore, M.M., Eds.; Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2008; tech cords; p. 282. [Google Scholar]
- Korb, J.E.; Springer, J.D. Herbaceous understory response to ponderosa pine restoration thinning and prescribed burning. In Ecological Restoration of Southwestern Ponderosa Pine Forests; Island Press: Washington, DC, USA, 2003; pp. 233–250. [Google Scholar]
- Graham, R.T.; McCaffrey, S.; Jain, T.B. Science Basis for Changing Forest Structure to Modify Wildfire Behavior and Severity; General Technical Report, RMRS-120; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2004; p. 43.
- Agee, J.K.; Skinner, C.N. Basic principles of forest fuel reduction treatments. For. Ecol. Manag. 2005, 211, 83–96. [Google Scholar] [CrossRef]
- Tinkham, W.D.; Hoffman, C.M.; Ex, S.A.; Battaglia, M.A.; Saralecos, J.D. Ponderosa Pine Forest Restoration Treatment Longevity: Implications of Regeneration on Fire Hazard. Forests 2016, 7, 137. [Google Scholar] [CrossRef]
- Kiefer, M.; van Wagtendonk, J.W.; Buhler, N. Long-term surface fuel accumulation in burned and unburned mixed conifer forests of the central and southern Sierra Nevada, CA (USA). Fire Ecol. 2006, 2, 53–72. [Google Scholar] [CrossRef]
- Vaillant, N.M.; Fites-Kaufman, J.; Stephens, S.L. Effectiveness of prescribed fire as a fuel treatment. Int. J. Wildland Fire 2009, 18, 165–175. [Google Scholar] [CrossRef]
- Willard, E.E. Some Factors Involved in Activation of Sprouting in Little Rabbitbrush and Snowberry on Summer Range. Master’s Thesis, Utah State University, Logan, UT, USA, 1971. [Google Scholar]
- Bradley, A.F. Rhizome Morphology, Soil Distribution, and the Potential Fire Survival of Eight Woody Understory Species in Western Montana. Master’s Thesis, University of Montana, Missoula, MT, USA, 1984. [Google Scholar]
- Engle, D.M.; Bonham, C.D.; Bartel, L.E. Ecological characteristics and control of Gambel oak. J. Range Manag. 1983, 36, 363–365. [Google Scholar] [CrossRef]
- Tiedemann, A.R.; Clary, W.P.; Barbour, R.J. Underground systems of Gambel oak (Quercus gambelii) in central Utah. Am. J. Bot. 1987, 74, 1065–1071. [Google Scholar] [CrossRef]
- Abrams, M.D. Adaptations and responses to drought in Quercus species of North America. Tree Physiol. 1990, 7, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Collins, B.M.; Moghaddas, J.J.; Stephens, S.L. Initial changes in forest structure and understory plant communities following fuel reduction activities in a Sierra Nevada mixed conifer forest. For. Ecol. Manag. 2007, 239, 102–111. [Google Scholar] [CrossRef]
- Converse, S.J.; White, G.C.; Farris, K.L.; Zack, S. Small mammals and forest fuel reduction: National-scale responses to fire and fire surrogates. Ecol. Appl. 2006, 16, 1717–1729. [Google Scholar] [CrossRef]
- Brown, H.E. Gambel oak in west-central Colorado. Ecology 1958, 39, 317–327. [Google Scholar] [CrossRef]
- Clary, W.P.; Tiedemann, A.R. Distribution of biomass within small tree and shrub from Quercus gambelli stands. For. Sci. 1986, 32, 234–242. [Google Scholar]
- Waltz, A.E.M.; Fulé, P.Z.; Covington, W.W.; Moore, M.M. Diversity in ponderosa pine forest structure following ecological restoration treatments. For. Sci. 2003, 49, 885–900. [Google Scholar]
- Savage, M.; Mast, J.N.; Feddema, J.J. Double whammy: High-severity fire and drought in ponderosa pine forests of the southwest. Can. J. For. Res. 2013, 43, 570–583. [Google Scholar] [CrossRef]
- Coop, J.D.; Parks, S.A.; McClernan, S.R.; Holsinger, L.M. Influences of prior wildfires on vegetation response to subsequent fire in a reburned Southwestern landscape. Ecol. Appl. 2016, 26, 346–354. [Google Scholar] [CrossRef]
- Walker, R.B.; Coop, J.D.; Parks, S.A.; Trader, L. Fire regimes approaching historic norms reduce wildfire-facilitated conversion from forest to non-forest. Ecosphere 2018, 9, e02182. [Google Scholar] [CrossRef]
- Larson, A.J.; Churchill, D.J. Tree spatial patterns in fire-frequent forests of western North America, including mechanisms of pattern formation and implications for designing fuel reduction and restoration treatments. For. Ecol. Manag. 2012, 267, 74–92. [Google Scholar] [CrossRef]
- Laughlin, D.C.; Bakker, J.D.; Daniels, M.L.; Moore, M.M.; Casey, C.A.; Springer, J.D. Restoring plant species diversity and community composition in a ponderosa pine–bunchgrass ecosystem. Plant Ecol. 2008, 197, 139–151. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment/Time | Species | Indicator Value | p-Value |
|---|---|---|---|
| TREATMENT | |||
| Control | Pedicularis racemosa | 75 | 0.05 |
| Control | Erigeron divergens | 74.6 | 0.017 |
| Burn | Paxistima myrsinites | 74.5 | 0.035 |
| Thin/Burn | Chamerion angustifolium | 83.3 | 0.02 |
| Thin/Burn | Cirsium arvense | 80.6 | 0.02 |
| Thin/Burn | Vicia americana | 69.5 | 0.007 |
| Thin/Burn | Galium boreale | 58.4 | 0.007 |
| Thin/Burn | Gentianella amarella | 46.8 | 0.017 |
| TIME | |||
| 2018 | Poa pratensis | 83.1 | 0.0002 |
| 2018 | Elymus elymoides | 78.6 | 0.0008 |
| 2018 | Poa fendleriana | 75.1 | 0.0004 |
| 2018 | Carex geophila | 68.0 | 0.0002 |
| 2018 | Bromus spp.* | 56.4 | 0.005 |
| Density (Stems/Trees ha−1) | Frequency (Proportion of Plots) | ||||||
|---|---|---|---|---|---|---|---|
| Species | Year | Control | Burn | Thin/Burn | Control | Burn | Thin/Burn |
| Density (stems ha−1) | |||||||
| AMAL | 2003 | 3516.3 | 4317.5 | 4712.5 | 0.93 | 0.91 | 0.93 |
| 2018 | 6565.0 | 8190.0 | 10,205.0 | 0.84 | 0.89 | 0.86 | |
| PRVI | 2003 | 251.3 | 433.8 | 885.0 | 0.20 | 0.28 | 0.50 |
| 2018 | 398.8 | 496.3 | 1501.3 | 0.15 | 0.28 | 0.55 | |
| QUGA | 2003 | 3666.3 | 4598.8 | 4426.3 | 0.75 | 0.71 | 0.85 |
| 2018 | 6222.5 | 7670.0 | 10,080.0 | 0.68 | 0.70 | 0.83 | |
| ROWO | 2003 | 1952.5 | 1613.8 | 1563.8 | 0.74 | 0.63 | 0.80 |
| 2018 | 3495.0 x,y | 2325.0 y | 4725.0 x | 0.65 | 0.59 | 0.78 | |
| SYRO | 2003 | 5785.0 | 7638.8 | 6413.8 | 0.99 | 1.00 | 0.95 |
| 2018 | 12,170.0 | 11,630 | 15,805 | 0.99 | 0.93 | 0.99 | |
| ABCO | 2003 | 166.3 | 107.5 | 347.5 | 0.51 | 0.43 | 0.60 |
| 2018 | 108.8 x | 63.8 x | 11.3 y | 0.36 | 0.26 | 0.06 | |
| PIPO | 2003 | 3.8 | 8.8 | 6.3 | 0.04 | 0.05 | 0.04 |
| 2018 | 163.8 x | 223.8 x | 25.0 y | 0.40 | 0.39 | 0.11 | |
| POTR | 2003 | 856.3 | 493.8 | 308.8 | 0.63 | 0.35 | 0.39 |
| 2018 | 837.5 | 682.5 | 1411.3 | 0.68 | 0.40 | 0.43 | |
| PSME | 2003 | 58.8 | 45.0 | 16.3 | 0.28 | 0.21 | 0.13 |
| 2018 | 108.8 x | 31.3 y | 5.0 z | 0.43 | 0.18 | 0.03 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korb, J.E.; Stoddard, M.T.; Huffman, D.W. Effectiveness of Restoration Treatments for Reducing Fuels and Increasing Understory Diversity in Shrubby Mixed-Conifer Forests of the Southern Rocky Mountains, USA. Forests 2020, 11, 508. https://doi.org/10.3390/f11050508
Korb JE, Stoddard MT, Huffman DW. Effectiveness of Restoration Treatments for Reducing Fuels and Increasing Understory Diversity in Shrubby Mixed-Conifer Forests of the Southern Rocky Mountains, USA. Forests. 2020; 11(5):508. https://doi.org/10.3390/f11050508
Chicago/Turabian StyleKorb, Julie E., Michael T. Stoddard, and David W. Huffman. 2020. "Effectiveness of Restoration Treatments for Reducing Fuels and Increasing Understory Diversity in Shrubby Mixed-Conifer Forests of the Southern Rocky Mountains, USA" Forests 11, no. 5: 508. https://doi.org/10.3390/f11050508
APA StyleKorb, J. E., Stoddard, M. T., & Huffman, D. W. (2020). Effectiveness of Restoration Treatments for Reducing Fuels and Increasing Understory Diversity in Shrubby Mixed-Conifer Forests of the Southern Rocky Mountains, USA. Forests, 11(5), 508. https://doi.org/10.3390/f11050508