Western Larch Regeneration Responds More Strongly to Site and Indirect Climate Factors Than to Direct Climate Factors


Abstract
1. Introduction
2. Materials and Methods
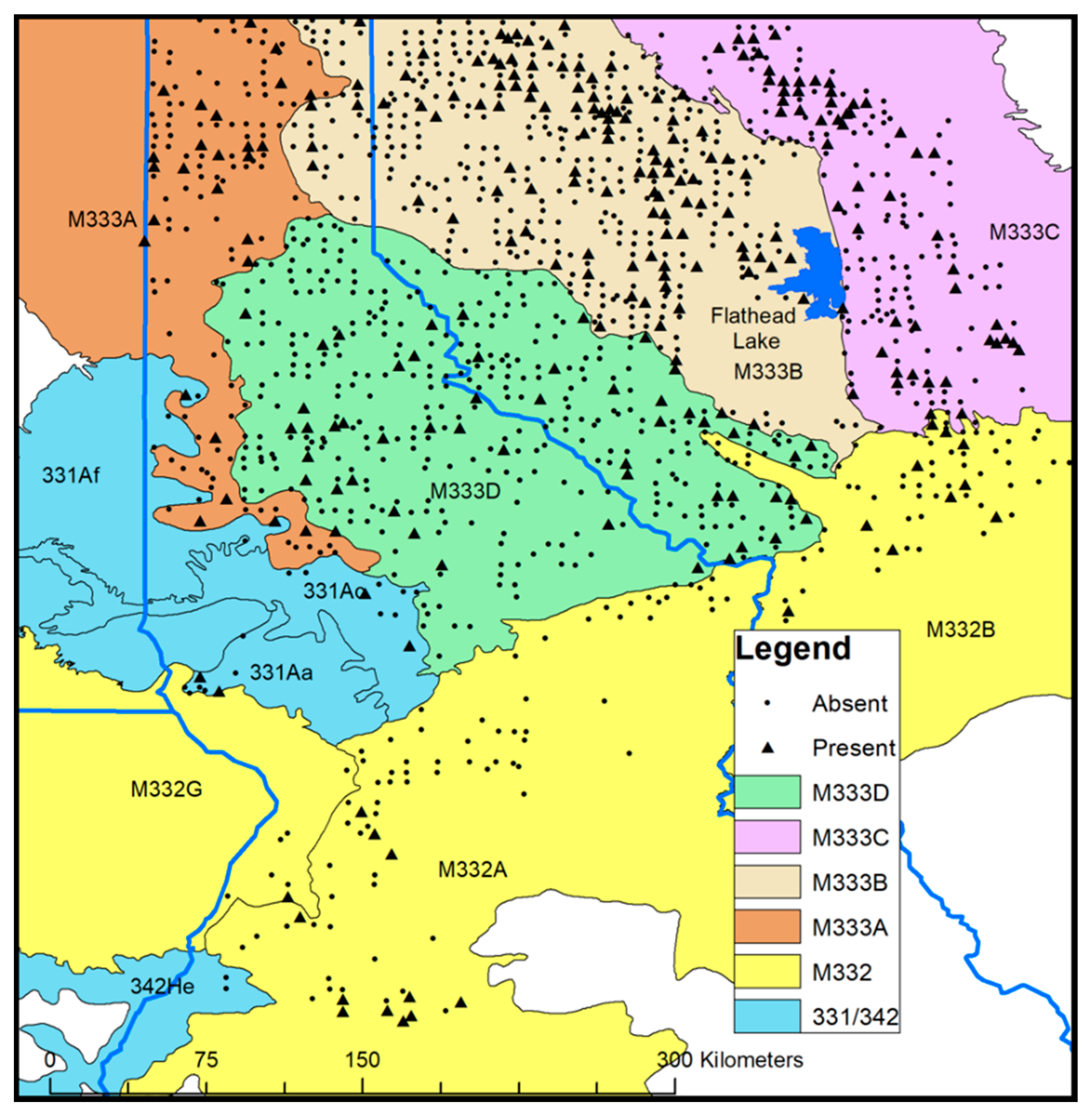
2.1. Study Design and Predictor Variables
2.2. Factors Related to Seedling Presence and Density
2.3. Trends in Seedling Presence and Density
2.4. Climate Niche Differentiation between Seedlings and Trees
3. Results
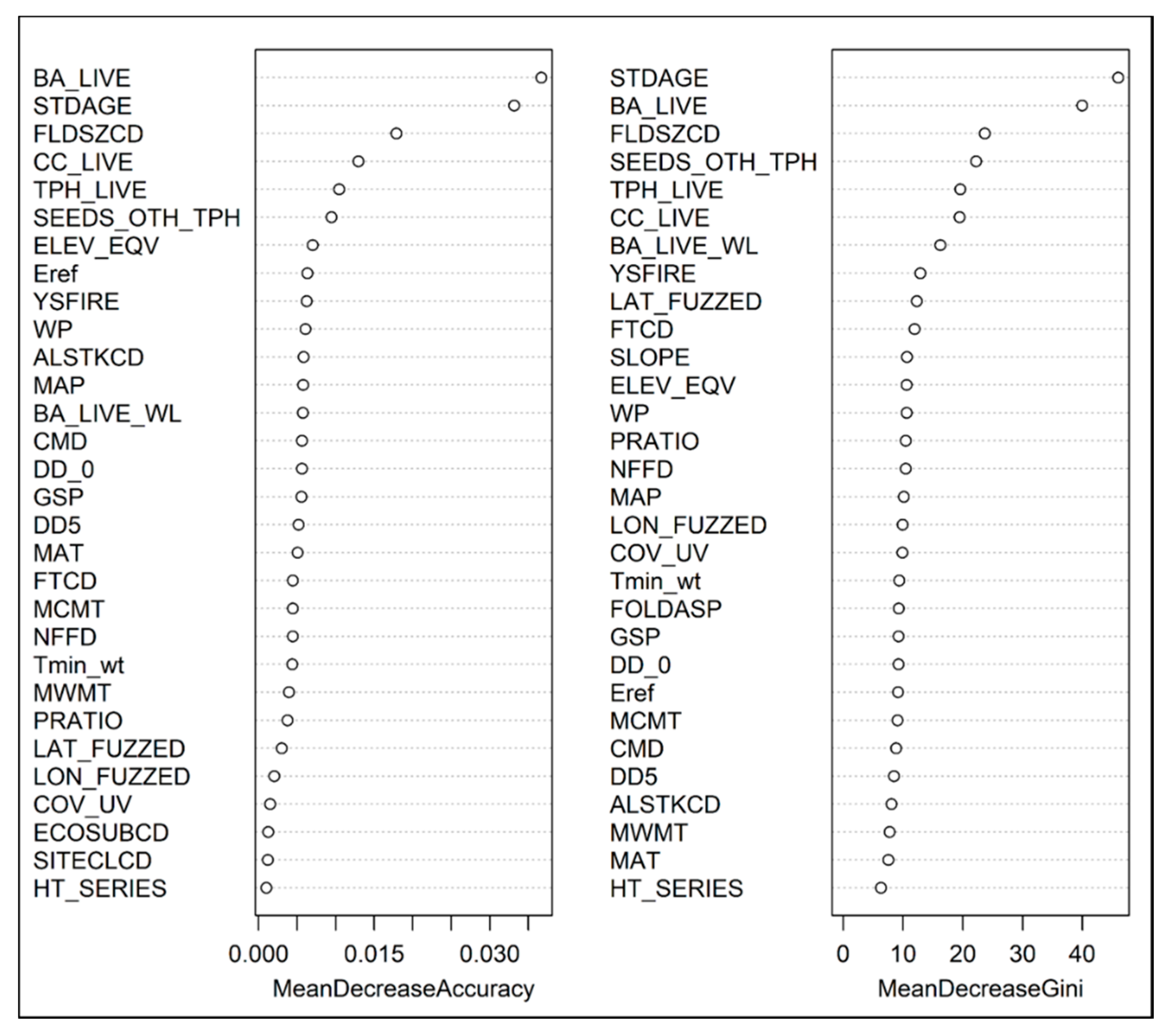
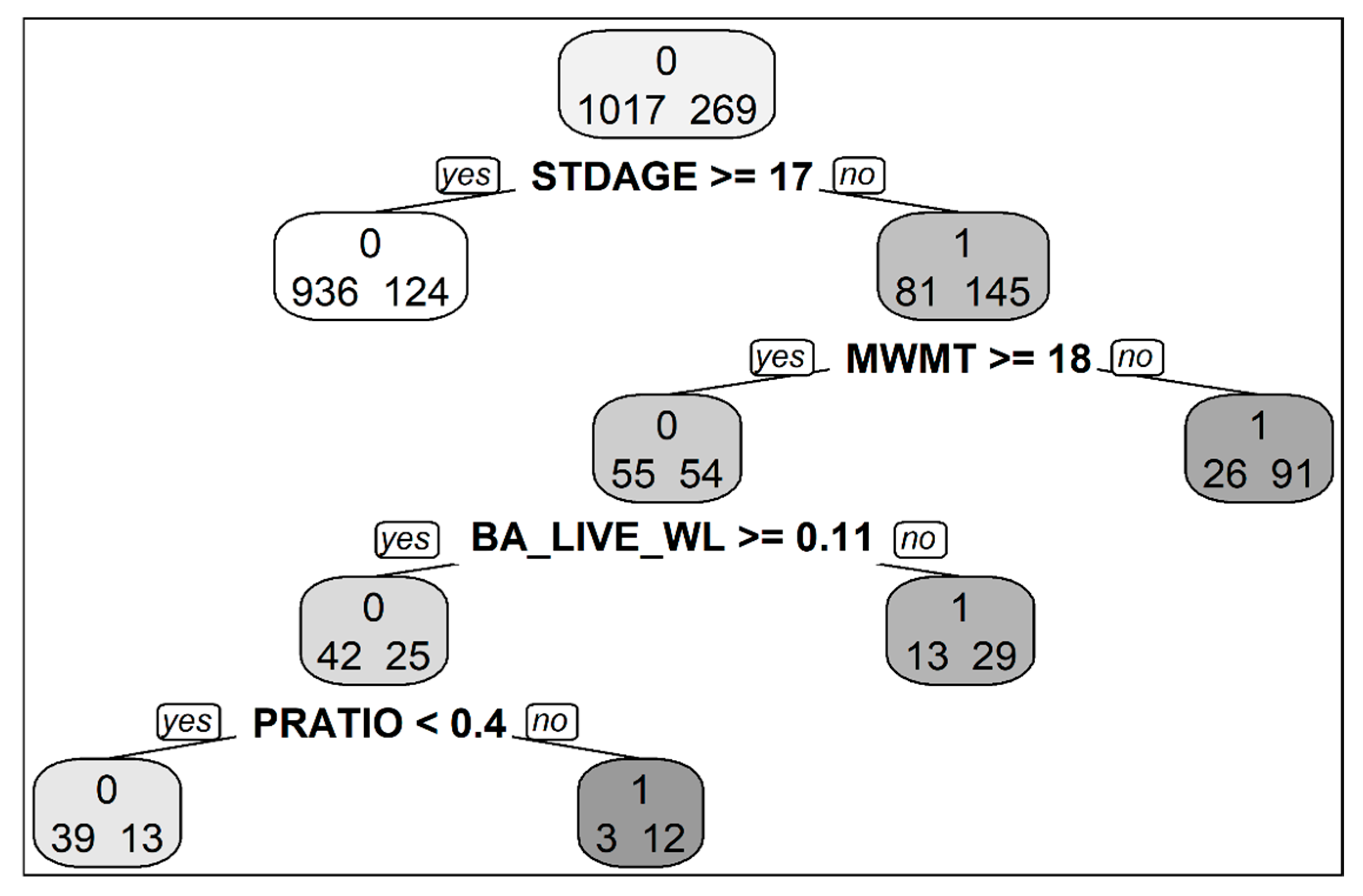
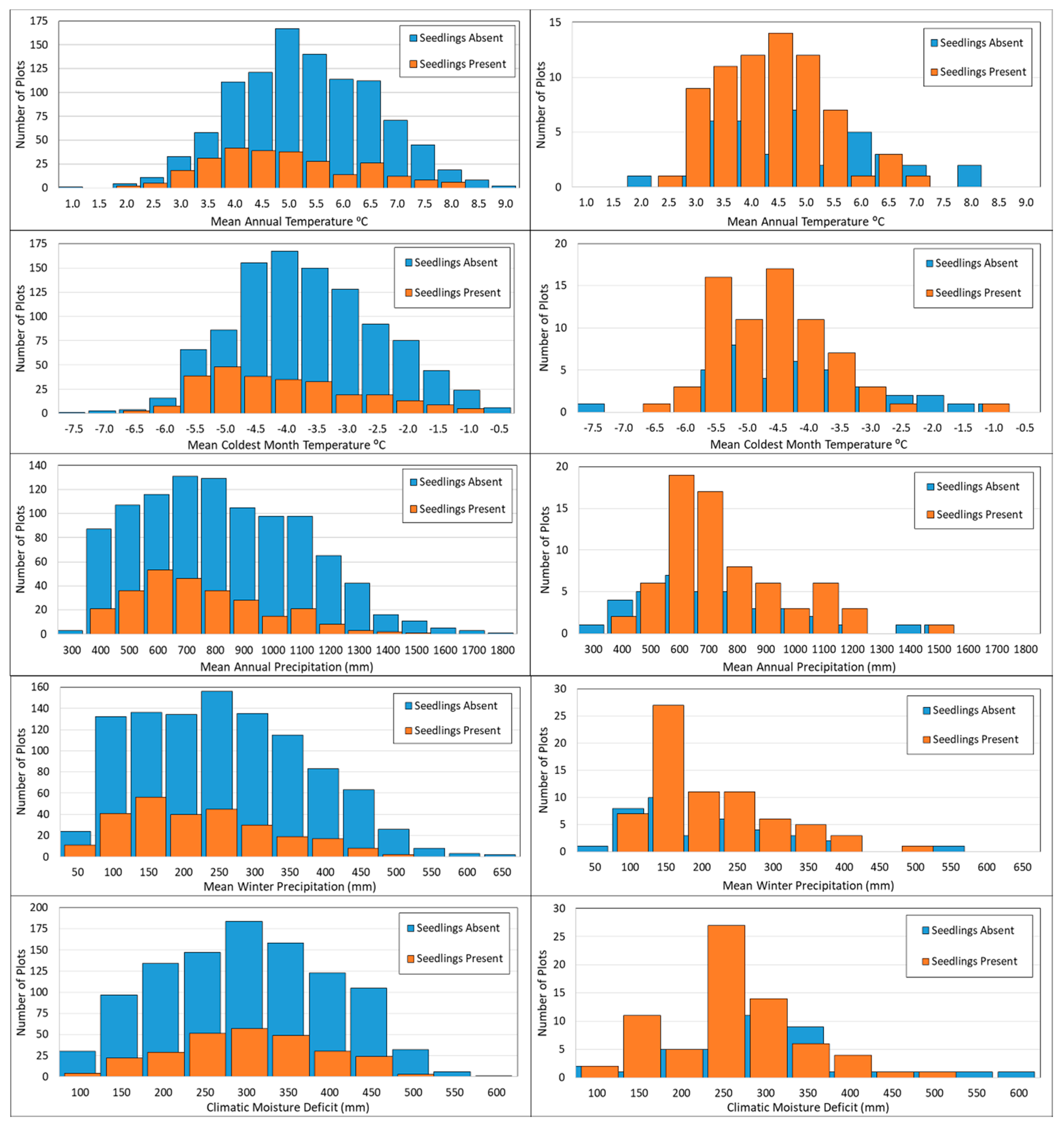
3.1. Factors Related to Seedling Presence and Density
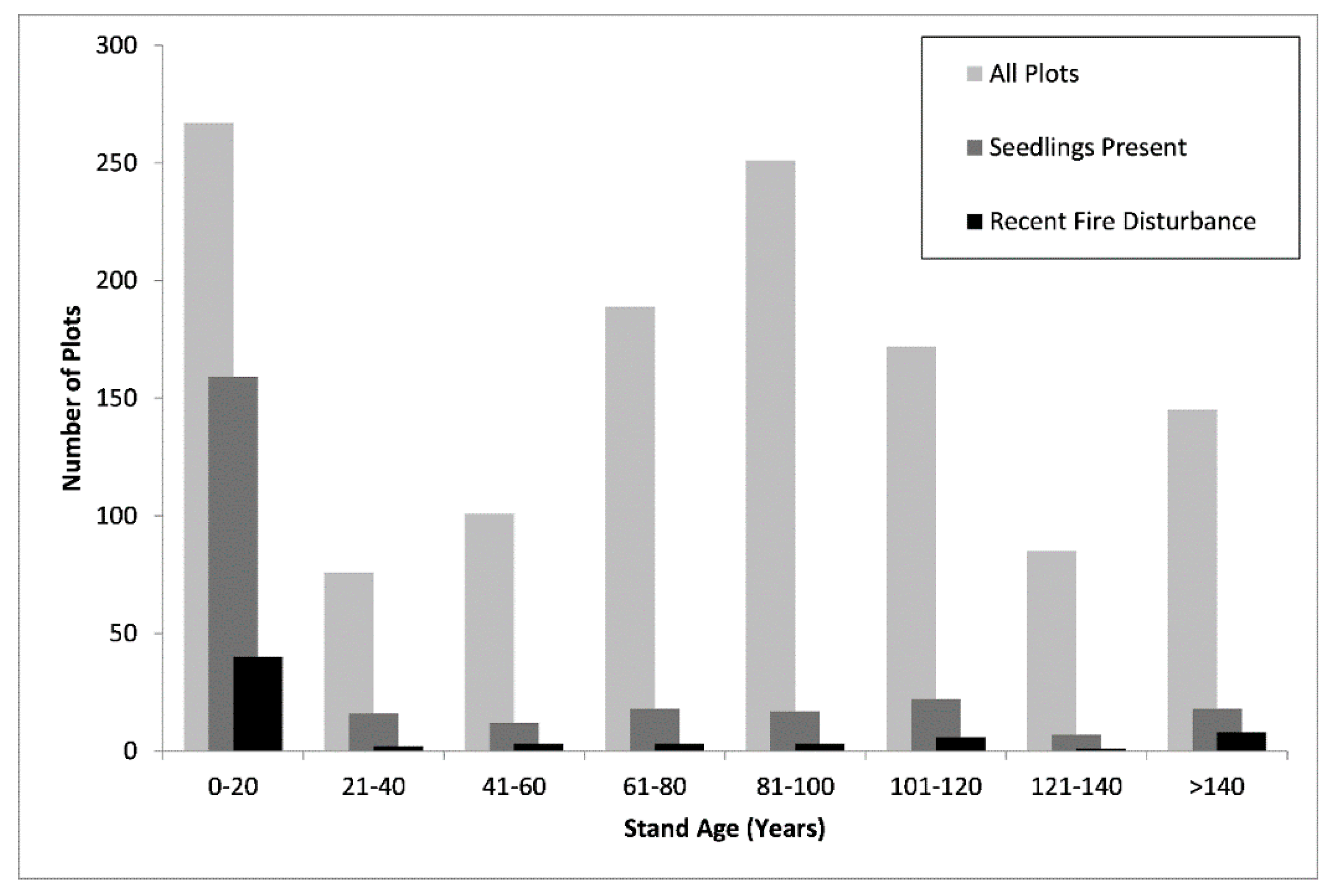
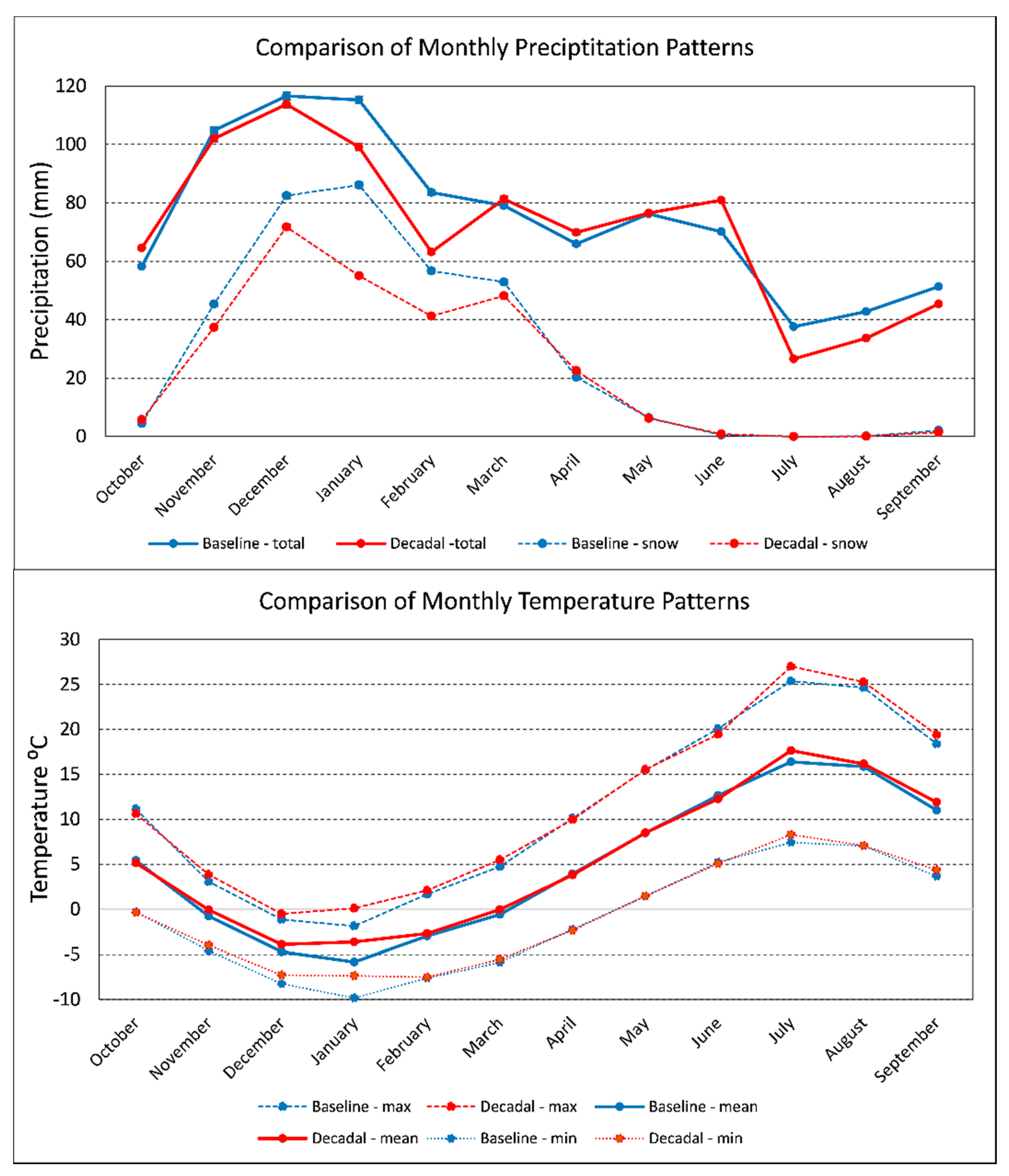
3.2. Trends in Seedling Presence and Density
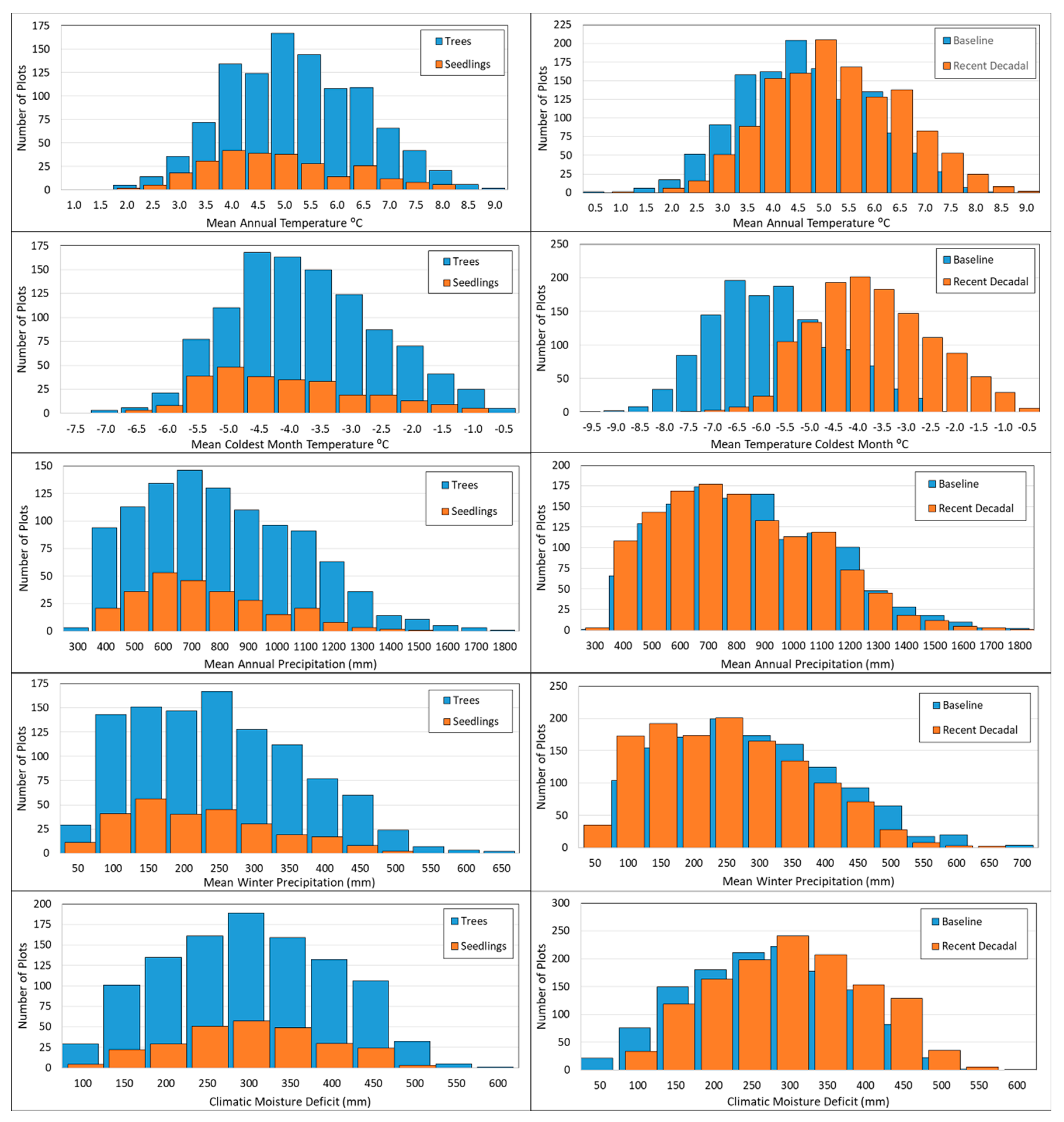
3.3. Climate Niche Differentiation between Seedlings and Trees
4. Discussion
4.1. Non-Climatic and Indirect Climatic Factors Affecting Seedling Recruitment
4.2. Climatic Factors Affecting Seedling Recruitment
4.3. Trends in Seedling Presence and Density
4.4. Climatic Niche Differentiation between Seedlings and Trees
4.5. Management Implications
4.6. Limitations of This Study
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time Period | Variable | MAP | GSP | WP | MAT | MWMT | MCMT | Tmin | DD_0 | DD5 | NFFD | Eref | CMD | PRATIO |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| baseline | Mean | 901 | 278 | 315 | 4.9 | 16.4 | −5.9 | −8.6 | 671 | 1372 | 165 | 678 | 301 | 0.322 |
| Median | 877 | 268 | 304 | 4.8 | 16.4 | −6.0 | −8.5 | 672 | 1359 | 166 | 669 | 302 | 0.303 | |
| 95th | 1394 | 393 | 528 | 7.2 | 18.5 | −3.5 | −6.1 | 923 | 1790 | 198 | 806 | 470 | 0.433 | |
| 5th | 497 | 199 | 136 | 2.9 | 14.1 | −7.8 | −11.0 | 416 | 996 | 132 | 552 | 136 | 0.243 | |
| decadal | Mean | 857 | 263 | 276 | 5.4 | 17.6 | −3.9 | −7.4 | 567 | 1438 | 169 | 696 | 327 | 0.324 |
| Median | 828 | 253 | 265 | 5.3 | 17.7 | −4.0 | −7.3 | 566 | 1425 | 170 | 695 | 327 | 0.309 | |
| 95th | 1342 | 371 | 483 | 7.6 | 19.8 | −1.8 | −5.0 | 798 | 1878 | 205 | 821 | 488 | 0.446 | |
| 5th | 465 | 187 | 109 | 3.4 | 15.2 | −5.8 | −9.9 | 342 | 1054 | 136 | 564 | 165 | 0.228 | |
| paired t-test (mean) | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | |
Appendix C
| Variable | Category | No. of Plots | Percent of Plots with Increase in Seedling Density | Cramer’s V | Effect | Mean Change in Seedling Density |
|---|---|---|---|---|---|---|
| cutting treatment | yes | 45 | 26.7 | 0.112 | higher likelihood w/cutting | 203 |
| no | 361 | 13.9 | 112 | |||
| ecoregion province/section 1 | 331/342 | 7 | 0.0 | 0.154 | −137 ab | |
| M332 | 49 | 10.2 | 78 ab | |||
| M333A | 26 | 15.4 | −80 ab | |||
| M333B | 140 | 11.4 | −37 a | |||
| M333C | 71 | 25.4 | 558 b | |||
| M333D | 113 | 16.8 | 126 ab | |||
| site productivity class | 2 | 5 | 0.0 | 0.2472 | higher likelihood in lowest site productivity class | −37 |
| 3 | 40 | 7.5 | −29 | |||
| 4 | 100 | 7.0 | −17 | |||
| 5 | 165 | 13.9 | 8 | |||
| 6 | 96 | 30.2 | 534 | |||
| stand-size class | 0 | 5 | 40.0 | 0.258 2,3 | Higher likelihood as stand size decreases | 74 |
| 1 | 44 | 34.1 | 530 | |||
| 2 | 97 | 10.8 | 51 | |||
| 3 | 247 | 17.5 | 65 | |||
| 4 | 13 | 38.5 | 375 | |||
| forest type 4 | DF | 136 | 8.8 | 0.213 | higher likelihood in LPP and WL | 0 |
| FSMH | 104 | 8.7 | 40 | |||
| HSS | 31 | 12.9 | 31 | |||
| LPP | 41 | 26.8 | 120 | |||
| OTHER | 17 | 29.4 | 393 | |||
| PP | NA 5 | NA | NA | |||
| WL | 77 | 24.7 | 426 | |||
| habitat type series 6 | ABGR | 75 | 16.0 | 0.082 | 118 | |
| ABLA | 126 | 18.3 | 81 | |||
| PICEA | NA | NA | NA | |||
| PSME | 65 | 15.4 | 14 | |||
| THPL | 67 | 14.9 | 347 | |||
| TSHE | 73 | 9.6 | 88 | |||
| stocking class | 1 (over) | 11 | 45.5 | 0.212 | higher likelihood in overstocked and poorly stocked stands | 2054 a |
| 2 (full) | 112 | 10.7 | −7 b | |||
| 3 (med) | 188 | 11.7 | 53 b | |||
| 4 (poor) | 93 | 24.7 | 192 ab | |||
| 5 (non) | 2 | 0.0 | 0 ab | |||
| ownership group | NF | 257 | 12.8 | 0.238 | higher likelihood with other federal ownership | 46 b |
| OTHFED | 14 | 57.1 | 1707 a | |||
| STATE | 40 | 7.5 | 27 b | |||
| PRIVATE/TRIBAL | 95 | 18.9 | 134 b |
Appendix D
| Variable | Mean for Plots with an Increase in Density | Mean for Plots without an Increase in Density | Correlation Coefficient with Change in Seedling Density |
|---|---|---|---|
| Years since fire disturbance | 51.5 | 71.8 | −0.293 |
| Equivalent elevation (m) | 1730.0 | 1694.7 | 0.010 |
| Aspect (folded) | 68.2 | 76.9 | −0.072 |
| Slope (percent) | 33.2 | 33.5 | 0.034 |
| Latitude | 47.7 | 47.7 | −0.012 |
| Longitude | −114.7 | −115.2 | 0.156 |
| Live basal area, all species (m2/ha) | 12.6 | 28.6 | −0.161 |
| Live basal area, all speciesdiff 1 | −5.4 | 1.9 | −0.292 |
| Live basal area, western larch (m2/ha) | 4.5 | 5.4 | −0.040 |
| Live basal area, western larchdiff | −0.7 | 0.4 | −0.240 |
| Live canopy cover (percent) | 39.7 | 59.7 | −0.100 |
| Live canopy coverdiff | 4.1 | 7.0 | −0.030 |
| Cover of understory vegetation (percent) | 48.6 | 48.2 | −0.089 |
| Stand age (years) | 69.3 | 84.8 | −0.005 |
| Live trees per hectare | 937.3 | 1414.5 | −0.108 |
| Live trees per hectarediff | 148.7 | 89.7 | −0.079 |
| Seedlings/hectare (other species) | 10828.1 | 2955.6 | 0.367 |
| Mean annual precipitation (mm) | 816.0 | 857.4 | −0.007 |
| Growing season precipitation (mm) | 265.8 | 265.3 | 0.062 |
| Winter precipitation (mm) | 252.7 | 274.1 | −0.039 |
| Mean annual temperature (°C) | 5.2 | 5.4 | −0.020 |
| Mean warmest month temp. (°C) | 17.5 | 17.6 | 0.004 |
| Mean coldest month temp. (°C) | −4.2 | −3.9 | −0.046 |
| Minimum winter temperature (°C) | −7.8 | −7.4 | −0.053 |
| Degree-days below 0 °C | 598.4 | 568.9 | 0.030 |
| Degree-days above 5 °C | 1405.6 | 1430.5 | −0.009 |
| Number of frost-free days | 164.1 | 167.5 | −0.035 |
| Reference evaporation (mm) | 682.6 | 697.2 | −0.015 |
| Climatic moisture deficit (mm) | 314.4 | 325.7 | −0.042 |
| Ratio of growing season to mean annual precipitation | 0.340 | 0.327 | 0.039 |
References
- McKenney, D.W.; Pedlar, J.H.; Lawrence, K.; Campbell, K.; Hitchinson, M.F. Potential impacts of climate change on the distribution of North American trees. BioScience 2007, 57, 939–948. [Google Scholar] [CrossRef]
- Coops, N.C.; Waring, R.H. Estimating the vulnerability of fifteen tree species under changing climate in Northwest North America. Ecol. Model. 2011, 222, 2119–2129. [Google Scholar] [CrossRef]
- Gray, L.K.; Hamann, A. Tracking suitable habitat for tree populations under climate change in western North America. Clim. Chang. 2013, 117, 289–303. [Google Scholar] [CrossRef]
- Bollenbacher, B.L.; Graham, R.T.; Reynolds, K.M. Regional forest landscape restoration priorities: Integrating historical conditions and an uncertain future in the northern Rocky Mountains. J. For. 2014, 112, 474–483. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Jaquish, B.C. Ecological impacts and management strategies for western larch in the face of climate-change. Mitig. Adapt. Strateg. Glob. Change 2010, 15, 283–306. [Google Scholar] [CrossRef]
- Hampe, A. Bioclimate envelope models: What they detect and what they hide. Global Ecol. Biogeogr. 2004, 13, 469–476. [Google Scholar] [CrossRef]
- Heikkinen, R.K.; Luoto, M.; Araújo, M.B.; Virkkala, R.; Thuiller, W.; Sykes, M.T. Methods and uncertainties in bioclimatic envelope modelling under climate change. Prog. Phys. Geogr. 2006, 30, 751–777. [Google Scholar] [CrossRef]
- Aiken, S.N.; Yeaman, S.; Holliday, J.A.; Wang, T.; Curtis-McLane, S. Adaptation, migration or extirpation: Climate change outcomes for tree populations. Evol. Appl. 2008, 1, 95–111. [Google Scholar] [CrossRef]
- Thuiller, W.; Albert, C.; Araujo, M.B.; Berry, P.M.; Cabeza, M.; Guisan, A.; Hickler, T.; Midgley, G.F.; Paterson, J.; Schurr, F.M.; et al. Predicting global change impacts on plant species’ distributions: Future challenges. Perspect. Plant. Ecol. 2007, 9, 137–152. [Google Scholar] [CrossRef]
- Haig, I.T.; Davis, K.P.; Weidman, R.H. Natural Regeneration in the Western White Pine Type; USDA Forest Service Technical Bulletin 767; USDA Forest Service: Washington, DC, USA, 1941.
- Ferguson, D.E.; Stage, A.R.; Boyd, R.J. Predicting regeneration in the grand fir-cedar-hemlock ecosystems of the Northern Rocky Mountains. For. Sci. 1986, 32, 1–41. [Google Scholar]
- Walck, J.L.; Hidayati, S.N.; Dixon, K.W.; Thompson, K.; Poschlod, P. Climate change and plant regeneration from seed. Glob. Chang. Biol. 2011, 17, 2145–2161. [Google Scholar] [CrossRef]
- Petrie, M.; Wildeman, A.; Bradford, J.; Hubbard, R.; Lauenroth, W. A review of precipitation and temperature control on seedling emergence and establishment for ponderosa and lodgepole pine forest regeneration. For. Ecol. Manag. 2016, 361, 328–338. [Google Scholar] [CrossRef]
- Davis, K.T.; Higuera, P.E.; Sala, A. Anticipating fire-mediated impacts of climate change using a demographic framework. Funct. Ecol. 2018, 32, 1729–1745. [Google Scholar] [CrossRef]
- Stevens-Rumann, C.S.; Kemp, K.B.; Higuera, P.E.; Harvey, B.J.; Rother, M.T.; Donato, D.C.; Morgan, P.; Veblen, T.T. Evidence for declining forest resilience to wildfires under climate change. Ecol. Lett. 2018, 21, 243–252. [Google Scholar] [CrossRef]
- Shearer, R.C. Insolation Limits Establishment of Western Larch Seedlings; USDA Forest Service: Washington, DC, USA; Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1967.
- Davis, K.T.; Dobrowski, S.Z.; Higuera, P.E.; Holden, Z.A.; Veblen, T.T.; Rother, M.T.; Parks, S.A.; Sala, A.; Maneta, M.P. Wildfires and climate change push low-elevation forests across a critical climate threshold for tree regeneration. Proc. Natl. Acad. Sci. USA 2019, 116, 6193–6198. [Google Scholar] [CrossRef]
- Shive, K.L.; Preisler, H.K.; Welch, K.R.; Safford, H.D.; Butz, R.J.; O’Hara, K.L.; Stephens, S.L. From the stand scale to the landscape scale: Predicting the spatial patterns of forest regeneration after disturbance. Ecol. Appl. 2018, 28, 1626–1639. [Google Scholar] [CrossRef]
- Korb, J.E.; Fornwalt, P.J.; Stevens-Rumann, C.S. What drives ponderosa pine regeneration following wildfire in the western United States? For. Ecol. Manag. 2019, 454, 1–16. [Google Scholar] [CrossRef]
- Andrus, R.A.; Harvey, B.J.; Rodman, K.C.; Hart, S.J.; Veblen, T.T. Moisture availability limits subalpine tree establishment. Ecology 2018, 99, 567–575. [Google Scholar] [CrossRef]
- Schmidt, W.C.; Shearer, R.C.; Roe, A.L. Ecology and Silviculture of Western Larch Forests; Technical Bulletin No. 1520; USDA Forest Service: Washington, DC, USA, 1976.
- Kemp, K.B.; Higuera, P.E.; Morgan, P. Fire legacies impact conifer regeneration across environmental gradients in the U.S. northern Rockies. Landsc. Ecol. 2016, 31, 619–636. [Google Scholar] [CrossRef]
- Urza, A.K.; Sibold, J.S. Climate and seed availability initiate alternate post-fire trajectories in a lower subalpine forest. J. Veg. Sci. 2017, 28, 43–56. [Google Scholar] [CrossRef]
- Hansen, W.D.; Braziunus, K.H.; Rammer, W.; Seidl, R.; Turner, M.G. It takes a few to tango: Changing climate and fire regimes can cause regeneration failure of two subalpine conifers. Ecology 2018, 99, 966–977. [Google Scholar] [CrossRef] [PubMed]
- Oswald, B.P.; Neuenschwander, L.F. Microsite variability and safe site description for western larch germination and establishment. Bull. Torrey Bot. Club 1993, 120, 148–156. [Google Scholar] [CrossRef]
- Dobrowski, S.Z.; Swanson, A.K.; Abatzoglou, J.T.; Holden, Z.A.; Safford, H.D.; Schwartz, M.K.; Gavin, D.G. Forest structure and species traits mediate projected recruitment declines in western US tree species. Glob. Ecol. Biogeogr. 2015, 24, 917–927. [Google Scholar] [CrossRef]
- Kroiss, S.J.; HilleRisLambers, J. Recruitment limitation of long-lived conifers: Implications for climate change responses. Ecology 2015, 96, 1286–1297. [Google Scholar] [CrossRef] [PubMed]
- Harvey, B.J.; Donato, D.C.; Turner, M.G. High and dry: Post-fire seedling establishment in subalpine forests decreases with post-fire drought and large stand-replacing burn patches. Global Ecol. Biogeogr. 2016, 25, 655–669. [Google Scholar] [CrossRef]
- Stevens-Rumann, C.; Morgan, P. Repeated wildfires alter forest recovery of mixed-conifer ecosystems. Ecol. Appl. 2016, 26, 1842–1853. [Google Scholar] [CrossRef]
- Kemp, K.B.; Higuera, P.E.; Morgan, P.; Abatzoglou, J.T. Climate will increasingly determine post-fire regeneration success of low-elevation forests, Northern Rockies, USA. Ecosphere 2019, 10, 1–17. [Google Scholar] [CrossRef]
- Gower, S.T.; Richards, J.H. Larches: Deciduous conifers in an evergreen world. BioScience 1990, 40, 818–826. [Google Scholar] [CrossRef]
- Clark, J.S.; Iverson, L.; Woodall, C.W.; Allen, C.D.; Bell, D.M.; Bragg, D.; D’Amato, A.W.; Davis, F.W.; Hersh, M.H.; Ibanez, I.; et al. The impacts of increasing drought on forest dynamics, structure, and biodiversity in the United States. Glob. Chang. Biol. 2016, 22, 2329–2352. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Allen, C.D.; Franklin, J.F.; Frelich, L.E.; Harvey, B.J.; Higuera, P.E.; Mack, M.C.; Meentemeyer, R.K.; Metz, M.R.; Perry, G.L.; et al. Changing disturbance regimes, ecological memory, and forest resilience. Front. Ecol. Environ. 2016, 14, 369–378. [Google Scholar] [CrossRef]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P. The interplay between climate change, forests, and disturbances. Sci. Total Environ. 2000, 262, 201–204. [Google Scholar] [CrossRef]
- Dennison, P.E.; Brewer, S.C.; Arnold, J.D.; Mortiz, M.A. Large wildfire trends in the western United States, 1984–2011. Geophys. Res. Lett. 2014, 41, 2928–2933. [Google Scholar] [CrossRef]
- Flannigan, M.D.; Stocks, B.J.; Wotton, B.M. Climate change and forest fires. Sci. Total Environ. 2000, 262, 221–229. [Google Scholar] [CrossRef]
- Brown, T.J.; Hall, B.L.; Westerling, A.L. The impact of twenty-first century climate change on wildland fire danger in the western United States: An applications perspective. Clim. Chang. 2004, 62, 365–388. [Google Scholar] [CrossRef]
- Abatzoglou, J.T.; Williams, A.P. Impact of anthropogenic climate change on wildfire across western US forests. Proc. Natl. Acad. Sci. USA 2016, 113, 11770–11775. [Google Scholar] [CrossRef]
- Jenkins, M.L.; Runyon, J.B.; Fettig, C.J.; Page, W.G.; Bentz, B.J. Interactions among the mountain pine beetle, fires, and fuels. For. Sci. 2014, 60, 489–501. [Google Scholar] [CrossRef]
- Ryan, K.C.; Reinhardt, E.D. Predicting postfire mortality of seven western conifers. Can. J. For. Res. 1988, 18, 1291–1297. [Google Scholar] [CrossRef]
- Schmidt, W.C.; Shearer, R.C. Larix occidentalis Nutt. western larch. In Silvics of North America; Burns, R.M., Honkala, B.H., Eds.; USDA Forest Service: Washington, DC, USA, 1990; pp. 160–172. [Google Scholar]
- Fiedler, C.; Harrington, M. Restoring vigor and reducing hazard in an old-growth western larch stand (Montana). Ecol. Restor. 2001, 22, 133–134. [Google Scholar]
- Pfister, R.D.; Kovalchik, B.L.; Arno, S.F.; Presby, R.C. Forest Habitat Types of Montana; General Technical Report INT-34; USDA Forest Service: Washington, DC, USA; Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1977; 174p.
- Westerling, A.L.; Hidalgo, H.G.; Cayan, D.R.; Swetnam, T.W. Warming and earlier spring increase western U.S. forest wildfire activity. Science 2006, 313, 940–943. [Google Scholar] [CrossRef]
- Zhu, K.; Woodall, C.W.; Clark, J.S. Failure to migrate: Lack of tree range expansion in response to climate change. Glob. Chang. Biol. 2012, 18, 1042–1052. [Google Scholar] [CrossRef]
- Bell, D.M.; Bradford, J.B.; Lauenroth, W.K. Early indicators of change: Divergent climate envelopes between tree life stages imply range shifts in the western United States. Glob. Ecol. Biogeogr. 2014, 23, 168–180. [Google Scholar] [CrossRef]
- Woodall, C.W.; Oswalt, C.M.; Westfall, J.A.; Perry, C.H.; Nelson, M.D.; Finley, A.O. An indicator of tree migration in forests of the eastern United States. For. Ecol. Manag. 2009, 257, 1434–1444. [Google Scholar] [CrossRef]
- Zhu, K.; Woodall, C.W.; Ghosh, S.; Gelfand, A.E.; Clark, J.S. Dual impacts of climate change: Forest migration and turnover through life history. Glob. Chang. Biol. 2014, 20, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Crookston, N.L.; Rehfeldt, G.E.; Dixon, G.E.; Weiskittel, A.R. Addressing climate change in the forest vegetation simulator to assess impacts on landscape forest dynamics. For. Ecol. Manag. 2010, 260, 1198–1211. [Google Scholar] [CrossRef]
- Bechtold, W.; Patterson, P. The Enhanced Forest Inventory and Analysis Program—National Sampling Design and Estimation Procedures; General Technical Report SRS-80; USDA Forest Service: Washington, DC, USA; Southern Research Station: Ashville, NC, USA, 2005; 85p.
- O’Connell, B.; Conkling, B.; Wilson, A.; Burrill, E.; Turner, J.; Pugh, S.; Christiansen, G.; Ridley, T.; Menlove, J. The Forest Inventory and Analysis Database: Database Description and User Guide Version 6.1.1 for Phase 2; USDA Forest Service: Washington, DC, USA, 2016; 870p.
- USDA. Interior West Forest Inventory and Analysis Forest Survey Field Procedures, Ver. 5.0; USDA Forest Service: Washington, DC, USA, 2013.
- McNab, W.; Cleland, D.T.; Freeouf, J.A.; Keys, J.E.; Nowacki, G.J.; Carpenter, C.A. Description of “Ecological Subregions: Sections of the Conterminous United States” First Approximation; General Technical Report WO-76B; USDA Forest Service: Washington, DC, USA, 2007; 80p.
- Goeking, S.A.; Izlar, D.K.; Edwards, T.C. A Landscape-Level Assessment of Whitebark Pine Regeneration in the Rocky Mountains, USA. For. Sci. 2019, 65, 87–99. [Google Scholar] [CrossRef]
- Eidenshink, J.; Schwind, B.; Brewer, K.; Zhu, Z.-L.; Quayle, B.; Howard, S. A project for monitoring trends in burn severity. Fire Ecol. 2009, 3, 3–21. [Google Scholar] [CrossRef]
- McCune, B.; Keon, D. Equations for potential annual direct incident radiation and heat load. J. Veg. Sci. 2002, 13, 603–606. [Google Scholar] [CrossRef]
- Windmuller-Campione, M.; Long, J. Limber pine (Pinus flexilis James), a flexible generalist of forest communities in the intermountain west. PLoS ONE 2016, 11, e0160324. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Hamann, A.; Spittlehouse, D.; Carroll, C. Locally downscaled and spatially customizable climate data for historical and future periods for North America. PLoS ONE 2016, 11, e0156720. [Google Scholar] [CrossRef]
- Daly, C.; Halbleib, M.; Smith, J.I.; Gibson, W.P.; Doggett, M.K.; Taylor, G.H.; Curtis, J.; Pasteris, P.P. Physiographically sensitive mapping of climatological temperature and precipitation across the conterminous United States. Int. J. Climatol. 2008, 28, 2031–2064. [Google Scholar] [CrossRef]
- Cutler, D.R.; Edwards, T.C.; Beard, K.H.; Cutler, A.; Hess, K.T.; Gibson, J.; Lawler, J.J. Random forests for classification in ecology. Ecology 2007, 88, 2783–2792. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- James, G.; Witten, D.; Hastie, T.; Tibshirani, R. An Introduction to Statistical Learning; Springer: New York, NY, USA, 2013; 426p. [Google Scholar]
- Therneau, T.; Atkinson, B.; Ripley, B. rpart: Recursive Partitioning and Regression Trees. R Package Version 4.1-10. 2015. Available online: https://CRAN.R-project.org/parkage=rpart (accessed on 30 March 2019).
- Breiman, L.; Friedman, J.; Olshen, R.; Stone, C.; Olshen, R. Classification and Regression Trees; Chapman and Hall: New York, NY, USA, 1984; 368p. [Google Scholar]
- Liaw, A.; Wiener, M. Classification and regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]
- Zar, J. Biostatistical Analysis, 3rd ed.; Prentice Hall: Upper Saddle River, NJ, USA, 1996; 662p. [Google Scholar]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Lawrence Erlbaum: Hillsdale, NJ, USA, 1988; 400p. [Google Scholar]
- Cooper, S.V.; Neiman, K.E.; Roberts, D.W. Forest Habitat Types of Northern Idaho: A Second Approximation; General Technical Report INT-236; USDA Forest Service: Washington, DC, USA; Intermountain Research Station: Ogden, UT, USA, 1991; 143p.
- Overpeck, J.; Rind, D.; Goldberg, R. Climate-induced changes in forest disturbance and vegetation. Nature 1990, 343, 51–53. [Google Scholar] [CrossRef]
- Arno, S.F.; Smith, H.Y.; Krebs, M.A. Old Growth Ponderosa Pine and Western Larch Stand Structures: Influences of Pre-1900 Fires and Fire Exclusion; Research Paper INT-RP-495; USDA Forest Service: Washington, DC, USA; Intermountain Research Station: Ogden, UT, USA, 1997; 20p.
- Hopkins, T.; Larson, A.J.; Belote, R.T. Contrasting effects of wildfire and ecological restoration in old-growth western larch forests. For. Sci. 2014, 60, 1005–1013. [Google Scholar] [CrossRef]
- Schmidt, W.C.; Shearer, R.C.; Naumann, J.R. Western larch. In Silvicultural Systems for the Major Forest Types of the United States; Burns, R.M., Ed.; USDA Forest Service: Washington, DC, USA, 1983; pp. 56–58. [Google Scholar]
- Sheehan, T.; Bachelet, D.; Ferschweiler, K. Projected major fire and vegetation changes in the Pacific Northwest of the conterminous United States under selected CMIP5 climate futures. Ecol. Model. 2015, 317, 16–29. [Google Scholar] [CrossRef]
- Chmura, D.J.; Anderson, P.D.; Howe, G.T.; Harrington, C.A.; Halofsky, J.E.; Peterson, D.L.; Shaw, D.C.; St.Clair, J.B. Forest responses to climate change in the northwestern United States: Ecophysiological foundations for adaptive management. For. Ecol. Manag. 2011, 261, 1121–1142. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? Glob. Ecol. Biogeogr. 2003, 12, 361–371. [Google Scholar] [CrossRef]
- Lenoir, J.; Gégout, J.C.; Pierrat, J.C.; Bontemps, J.D.; Dhôte, J.F. Differences between tree species seedling and adult altitudinal distribution in mountain forests during the recent warm period (1986–2006). Ecography 2009, 32, 765–777. [Google Scholar] [CrossRef]
- McDaniel, P.A.; Wilson, M.A. Physical and chemical characteristics of ash-influenced soils of Inland Northwest forests. In Volcanic-Ash-Influenced Forest Soils of the Inland Northwest: Properties and Implications for Management and Restoration; Page-Dumroese, D., Miller, R., Mital, J., McDaniel, P., Miller, D., Eds.; USDA Forest Service: Washington, DC, USA; Rocky Mountain Research Station: Fort Collins, CO, USA, 2007; pp. 31–45. [Google Scholar]






| Predictor Group | Description | Code | Source | Type |
|---|---|---|---|---|
| abiotic site variable | cutting treatment | CUTTING | Derived from FIA data | factor |
| ecoregion province/section | ECOSUBCD | FIA data (ECOPROV) | factor | |
| site productivity class | SITECLCD | FIA data (SITECLCD) | factor | |
| years since fire disturbance 1 | YSFIRE | FIA and MTBS data | numeric | |
| equivalent elevation adjusted for latitude | ELEV_EQV | Derived from DEM 3 | numeric | |
| aspect (folded about a north-south axis) | FOLDASP | Derived from DEM 3 | numeric | |
| latitude | LAT_FUZZED | FIA data (LAT_FUZZED) | numeric | |
| longitude | LON_FUZZED | FIA data (LON_FUZZED) | numeric | |
| slope (percent) | SLOPE | FIA data (SLOPE) | numeric | |
| biotic stand variable | stand-size class (field designation) | FLDSZCD | FIA data (FLDSZCD) | factor |
| forest type (field designation) 2 | FLDTYPCD | FIA data (FLDTYPCD) | factor | |
| habitat type series | HT_SERIES | FIA data (HABTYPCD1) | factor | |
| all live stocking code | ALSTKCD | FIA data (ALSTKCD) | factor | |
| live basal area, all species (m2/ha) | BA_LIVE | Derived from FIA data | numeric | |
| live basal area, wl (m2/ha) | BA_LIVE_WL | Derived from FIA data | numeric | |
| live canopy cover (percent) | CC_LIVE | Derived from FIA data | numeric | |
| cover of shrubs, forbs, and graminoids (%) | COV_UV | Derived from FIA data | numeric | |
| stand age (years) | STDAGE | FIA data (STDAGE) | numeric | |
| live trees per hectare | TPH_LIVE | FIA data | numeric | |
| seedlings/hectare of all other tree species | SEEDS_TPH_OTH | FIA data | numeric | |
| climatic variable | mean annual precipitation (mm) | MAP | Derived from PRISM data | numeric |
| growing-season prec. (May-Sept.) (mm) | GSP | Derived from PRISM data | numeric | |
| winter precipitation (mm) | WP | Derived from PRISM data | numeric | |
| mean annual temperature (°C) | MAT | Derived from PRISM data | numeric | |
| mean warmest month temperature (°C) | MWMT | Derived from PRISM data | numeric | |
| mean coldest month temperature (°C) | MCMT | Derived from PRISM data | numeric | |
| minimum winter temperature (°C) | Tmin_wt | Derived from PRISM data | numeric | |
| degree-days below 0 °C | DD_0 | Derived from PRISM data | numeric | |
| degree-days above 5 °C | DD5 | Derived from PRISM data | numeric | |
| number of frost-free days | NFFD | Derived from PRISM data | numeric | |
| Hargreaves ref. evaporation (mm) | Eref | Derived from PRISM data | numeric | |
| Hargreaves climatic moisture deficit (mm) | CMD | Derived from PRISM data | numeric | |
| GSP/MAP | PRATIO | Derived from PRISM data | numeric | |
| management | artificial regeneration of western larch | PLANT | FIA data (STDORGCD) | factor |
| ownership group | OWNGRPCD | FIA data (OWNGRPCD) | factor |
| Statistical Model | Confusion Matrices | Performance Metrics | |||
|---|---|---|---|---|---|
| Logistic regression | Predicted | PCC | 0.850 | ||
| A | P | Kappa | 0.495 | ||
| Observed | A | 958 | 59 | AUC | 0.865 |
| P | 134 | 135 | Sensitivity | 0.502 | |
| Specificity | 0.942 | ||||
| FP | 0.304 | ||||
| TSS | 0.444 | ||||
| Classification tree | Predicted | PCC | 0.836 | ||
| A | P | Kappa | 0.442 | ||
| Observed | A | 952 | 65 | AUC | 0.732 |
| P | 146 | 123 | Sensitivity | 0.457 | |
| Specificity | 0.936 | ||||
| FP | 0.346 | ||||
| TSS | 0.393 | ||||
| Random forests | Predicted | PCC | 0.855 | ||
| A | P | Kappa | 0.509 | ||
| Observed | A | 965 | 50 | AUC | 0.891 |
| P | 133 | 136 | Sensitivity | 0.506 | |
| Specificity | 0.948 | ||||
| FP | 0.277 | ||||
| TSS | 0.454 | ||||
| Variable | Category | No. of Plots | Percent of Plots with Seedlings Present | Cramer’s V | Effect | Median Seedling Density |
|---|---|---|---|---|---|---|
| cutting treatment | yes | 95 | 42.1 | 0.147 | higher likelihood w/cutting treatment | 370 |
| no | 1191 | 19.2 | 556 | |||
| ecoregion province/section 1 | 331/342 | 25 | 24.0 | 0.147 | higher likelihood in M333B and M333C and lower in M333D | 463 ab |
| M332 | 160 | 16.2 | 370 ab | |||
| M333A | 123 | 19.5 | 185 a | |||
| M333B | 423 | 25.8 | 741 b | |||
| M333C | 182 | 29.1 | 765 ab | |||
| M333D | 373 | 13.7 | 370 ab | |||
| site productivity class | 2 | NA | NA | 0.2142 | higher likelihood as site productivity decreases | NA |
| 3 | 144 | 11.1 | 741 | |||
| 4 | 324 | 16.4 | 556 | |||
| 5 | 532 | 17.8 | 556 | |||
| 6 | 286 | 36.7 | 494 | |||
| stand-size class | 0 | 12 | 50.0 | 0.4632 | higher likelihood as stand size decreases | 309 |
| 1 | 195 | 63.8 | 740 | |||
| 2 | 283 | 19.1 | 403 | |||
| 3 | 756 | 10.5 | 450 | |||
| 4 | 40 | 12.5 | 370 | |||
| forest type 3 | DF | 427 | 15.0 | 0.264 | higher likelihood in WL and LPP and lower in FSMH, DF, and HSS | 370 a |
| FSMH | 297 | 11.7 | 247 a | |||
| HSS | 110 | 10.9 | 185 a | |||
| LPP | 148 | 33.8 | 556 ab | |||
| OTHER | 9 | 33.3 | 926 ab | |||
| PP | 37 | 21.6 | 335 ab | |||
| WL | 258 | 37.6 | 1297 b | |||
| habitat type series 4 | ABGR | 235 | 19.6 | 0.141 | higher likelihood in ABLA series and lower in THPL series | 494 |
| ABLA | 352 | 29.8 | 556 | |||
| PICEA | 18 | 16.7 | 556 | |||
| PSME | 214 | 19.3 | 219 | |||
| THPL | 203 | 14.4 | 556 | |||
| TSHE | 264 | 16.7 | 741 | |||
| stocking class | 1 (over) | 50 | 56.0 | 0.202 | likelihood is highest at low and high stocking levels | 2532 c |
| 2 (full) | 388 | 19.1 | 753 bc | |||
| 3 (med) | 548 | 15.9 | 370 ab | |||
| 4 (poor) | 283 | 26.8 | 370 a | |||
| 5 (non) | 17 | 23.5 | 216 abc | |||
| artificial regeneration | yes | 18 | 55.6 | 0.1025 | 727 | |
| no | 1268 | 20.4 | 556 | |||
| ownership group | NF | 804 | 17.7 | 0.114 | lower likelihood on National Forest and higher on other federal and private | 590 |
| OTHFED | 50 | 34.0 | 2223 | |||
| STATE | 109 | 22.0 | 463 | |||
| PRIVATE/TRIBAL | 323 | 26.5 | 370 |
| Variable | Mean for Plots with Seedlings Present | Mean for Plots with Seedlings Absent | Correlation Coefficient with Seedling Density |
|---|---|---|---|
| Years since fire disturbance | 58.0 | 72.5 | −0.253 |
| Equivalent elevation (m) | 1766.0 | 1667.0 | 0.130 |
| Aspect (folded) | 72.1 | 72.3 | −0.026 |
| Slope (percent) | 28.1 | 35.3 | −0.068 |
| Latitude | 47.8 | 47.7 | 0.114 |
| Longitude | −115.1 | −115.3 | 0.083 |
| Live basal area, all species (m2/ha) | 12.1 | 30.6 | −0.204 |
| Live basal area, western larch (m2/ha) | 3.6 | 5.2 | 0.015 |
| Live canopy cover (percent) | 41.1 | 61.7 | −0.094 |
| Cover of understory veg. (percent) | 41.8 | 43.1 | −0.066 |
| Stand age (years) | 47.9 | 88.0 | −0.131 |
| Live trees per hectare | 1314.9 | 1340.5 | 0.007 |
| Seedlings/hectare (other species) | 8246.4 | 3527.7 | 0.268 |
| Mean annual precipitation (mm) | 787.9 | 875.0 | −0.062 |
| Growing season precipitation (mm) | 254.2 | 265.3 | −0.006 |
| Winter precipitation (mm) | 246.3 | 283.8 | −0.066 |
| Mean annual temperature (°C) | 5.1 | 5.5 | −0.134 |
| Mean warmest month temp. (°C) | 17.3 | 17.7 | −0.132 |
| Mean coldest month temperature (°C) | −4.3 | −3.8 | −0.138 |
| Minimum winter temperature (°C) | −7.9 | −7.3 | −0.139 |
| Degree-days below 0 °C | 610.3 | 555.8 | 0.137 |
| Degree-days above 5 °C | 1377.7 | 1454.5 | −0.128 |
| Number of frost-free days | 162.7 | 170.5 | −0.131 |
| Reference evaporation (mm) | 679.7 | 700.1 | −0.110 |
| Climatic moisture deficit (mm) | 324.7 | 327.3 | −0.062 |
| Ratio of growing season to mean annual precipitation | 0.338 | 0.320 | 0.084 |
| Age Class | Measure | MAP | GSP | WP | MAT | MWMT | MCMT | Tmin | DD_0 | DD5 | NFFD | Eref | CMD | PRATIO |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| seedlings | Mean | 788 | 254 | 246 | 5.1 | 17.3 | −4.3 | −7.9 | 610 | 1378 | 163 | 680 | 325 | 0.338 |
| Median | 742 | 246 | 232 | 4.9 | 17.3 | −4.5 | −8.1 | 621 | 1354 | 160 | 672 | 325 | 0.338 | |
| 95th | 1209 | 339 | 439 | 7.5 | 19.7 | −1.9 | −5.2 | 840 | 1837 | 205 | 806 | 478 | 0.455 | |
| 5th | 475 | 190 | 100 | 3.3 | 15.0 | −5.9 | −10.3 | 354 | 1017 | 130 | 560 | 173 | 0.231 | |
| Tolerance | 731 | 150 | 339 | 4.3 | 4.7 | 4.0 | 5.2 | 487 | 821 | 75 | 247 | 305 | 0.224 | |
| trees | Mean | 856 | 263 | 293 | 5.4 | 17.6 | −3.9 | −7.4 | 566 | 1439 | 169 | 696 | 327 | 0.324 |
| Median | 828 | 252 | 265 | 5.4 | 17.7 | −4.0 | −7.3 | 565 | 1427 | 169 | 696 | 324 | 0.310 | |
| 95th | 1346 | 371 | 486 | 7.6 | 19.8 | −1.8 | −5.0 | 796 | 1885 | 205 | 819 | 490 | 0.447 | |
| 5th | 460 | 187 | 109 | 3.4 | 15.3 | −5.8 | −9.9 | 343 | 1057 | 136 | 565 | 164 | 0.229 | |
| Tolerance | 887 | 184 | 378 | 4.2 | 4.5 | 4.0 | 4.9 | 453 | 828 | 69 | 254 | 326 | 0.218 | |
| t-test (mean) | <0.01 | 0.02 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | 0.75 | <0.01 | |
| difference (median) | 0.097 | 0.033 | 0.086 | 0.119 | 0.089 | 0.125 | 0.163 | −0.124 | 0.088 | 0.130 | 0.094 | −0.003 | −0.129 | |
| difference (95th) | 0.155 | 0.176 | 0.125 | 0.024 | 0.033 | 0.025 | 0.031 | −0.096 | 0.058 | 0.000 | 0.053 | 0.040 | −0.036 | |
| difference (5th) | −0.021 | −0.014 | 0.023 | 0.048 | 0.067 | 0.025 | 0.082 | −0.023 | 0.049 | 0.087 | 0.022 | −0.028 | −0.009 | |
| response type 1 | C | C | ↓ | ↓ | ↓ | ↓ | ↓ | ↑ | ↓ | ↓ | ↓ | C | ↑ | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steed, J.E.; Goeking, S.A. Western Larch Regeneration Responds More Strongly to Site and Indirect Climate Factors Than to Direct Climate Factors. Forests 2020, 11, 482. https://doi.org/10.3390/f11040482
Steed JE, Goeking SA. Western Larch Regeneration Responds More Strongly to Site and Indirect Climate Factors Than to Direct Climate Factors. Forests. 2020; 11(4):482. https://doi.org/10.3390/f11040482
Chicago/Turabian StyleSteed, James E., and Sara A. Goeking. 2020. "Western Larch Regeneration Responds More Strongly to Site and Indirect Climate Factors Than to Direct Climate Factors" Forests 11, no. 4: 482. https://doi.org/10.3390/f11040482
APA StyleSteed, J. E., & Goeking, S. A. (2020). Western Larch Regeneration Responds More Strongly to Site and Indirect Climate Factors Than to Direct Climate Factors. Forests, 11(4), 482. https://doi.org/10.3390/f11040482