Distribution Pattern of Endangered Plant Semiliquidambar cathayensis (Hamamelidaceae) in Response to Climate Change after the Last Interglacial Period


Abstract
1. Introduction
2. Materials and Methods
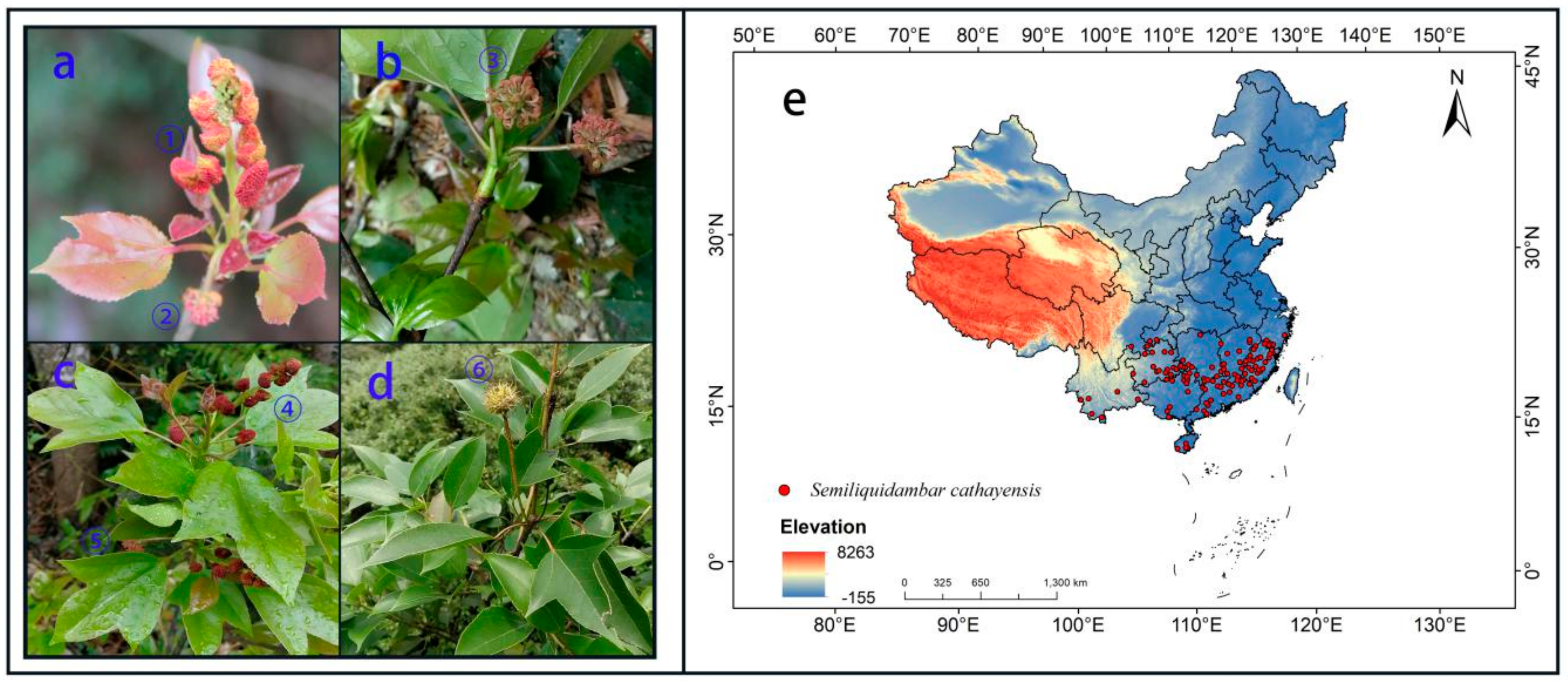
2.1. Study Area
2.2. Collection and Screening of Sample Data
2.3. Environmental Variable Screening and Data Processing
2.4. Model Establishment, Optimization, and Evaluation
2.5. Multi-Environment Similarity Surface (MESS) and the Most Dissimilar (MoD) Variable Analysis
2.6. Analysis of Spatial Pattern Changes
2.7. The core Distributional Shifts
3. Results
3.1. Analysis of the Accuracy of the Model
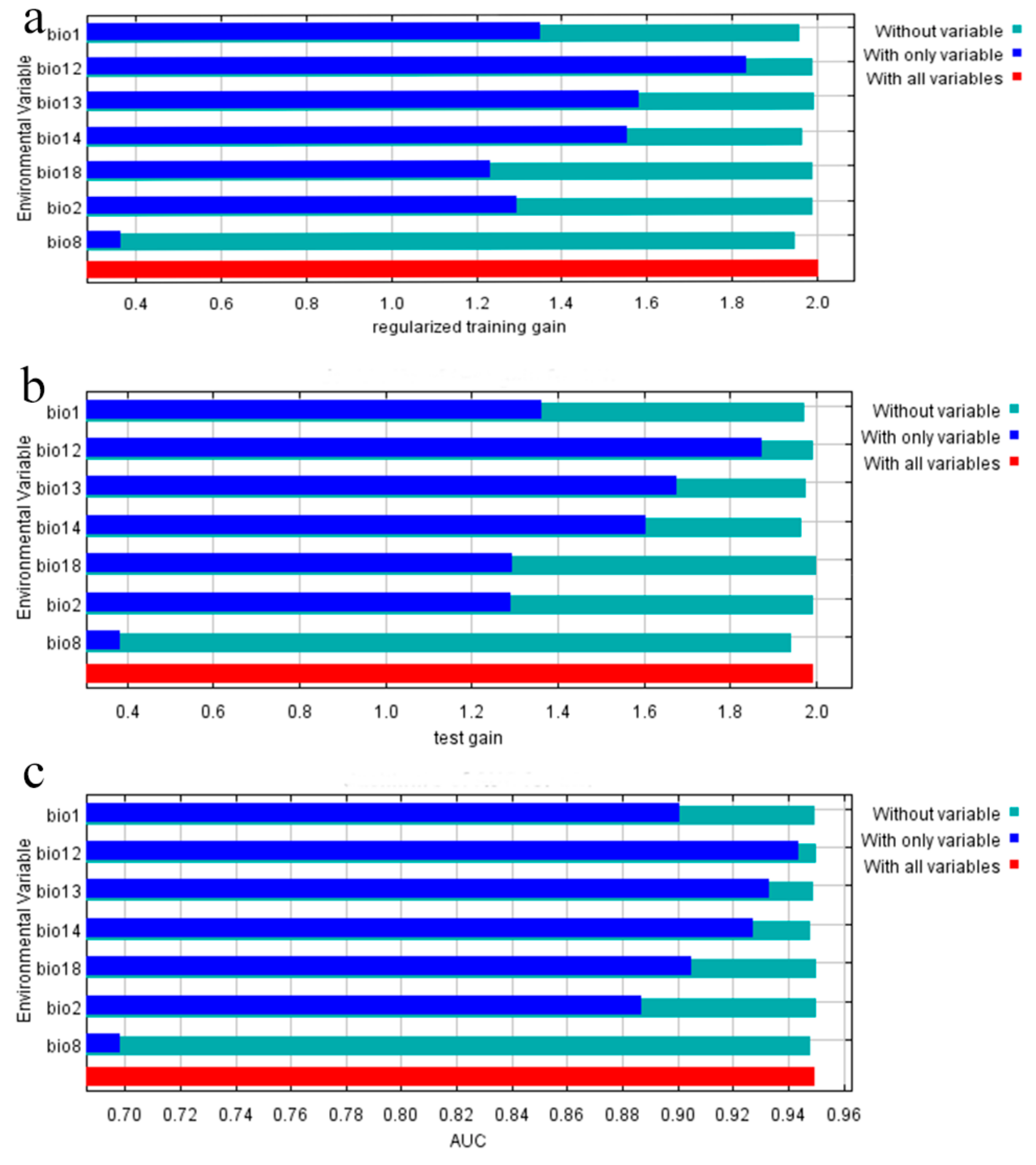
3.2. Contribution Analysis of Environmental Variables
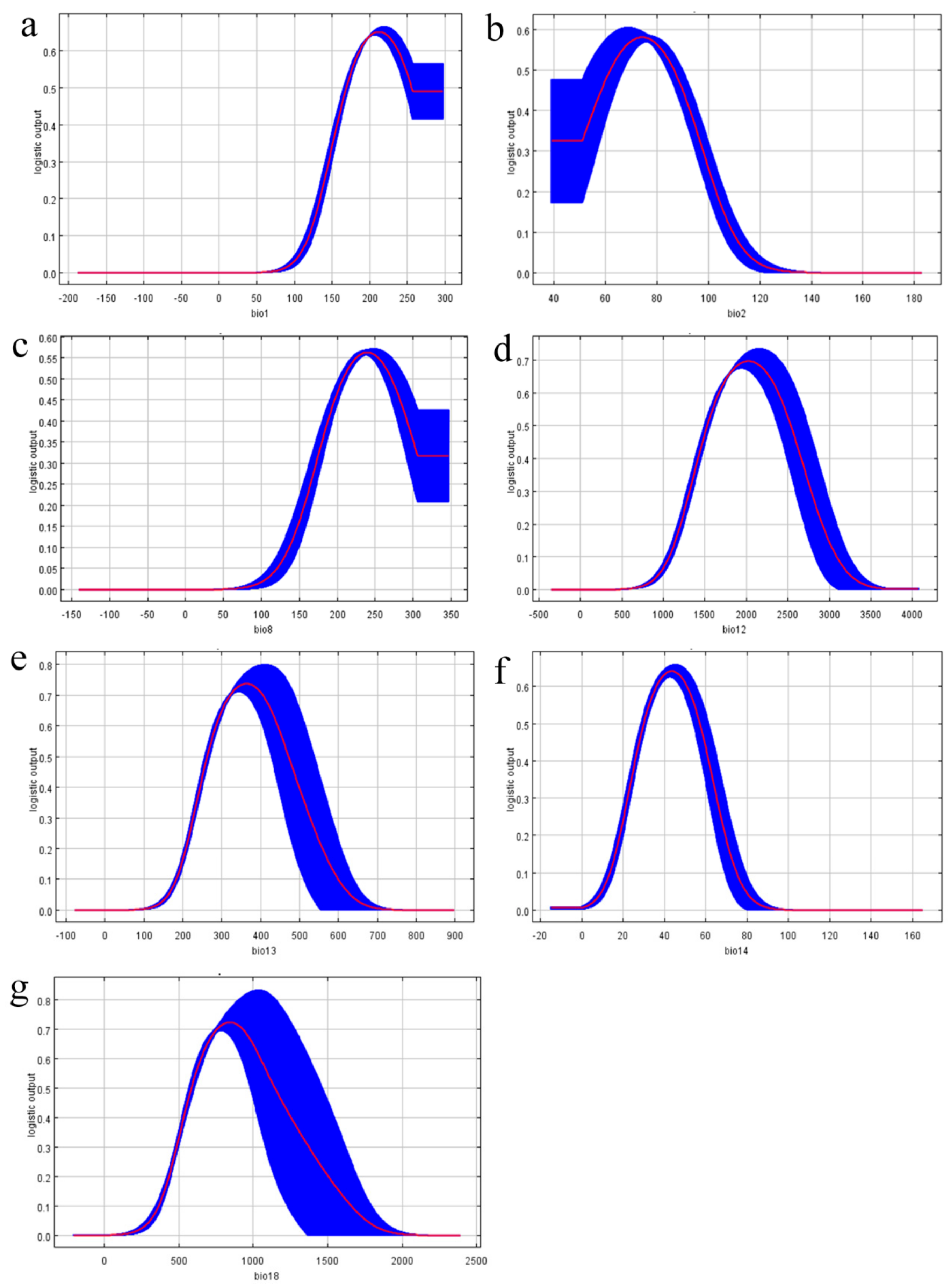
3.3. Environmental Variables Analysis
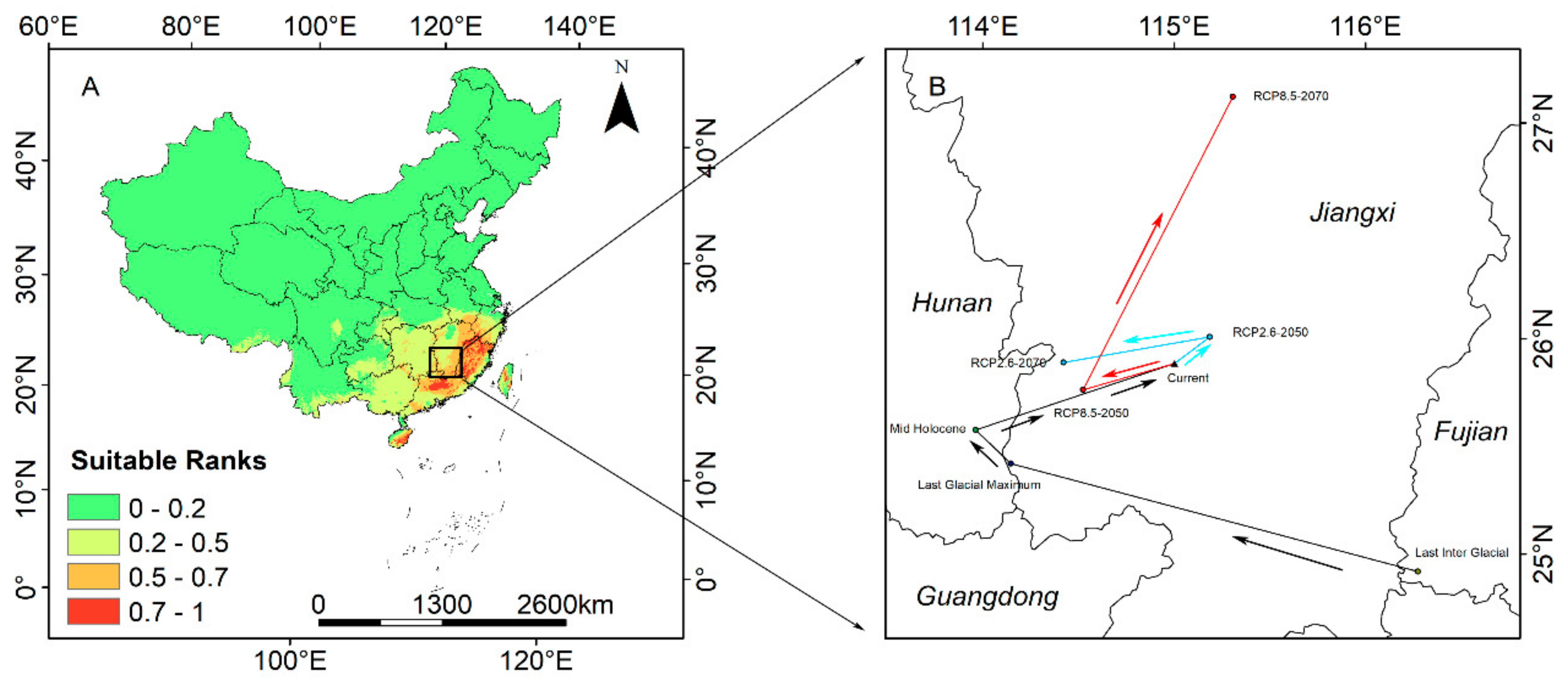
3.4. Modern Potential Distribution Areas
3.5. Potential Distribution Area in the Past
3.6. Future Changes in Suitable Habitat Area
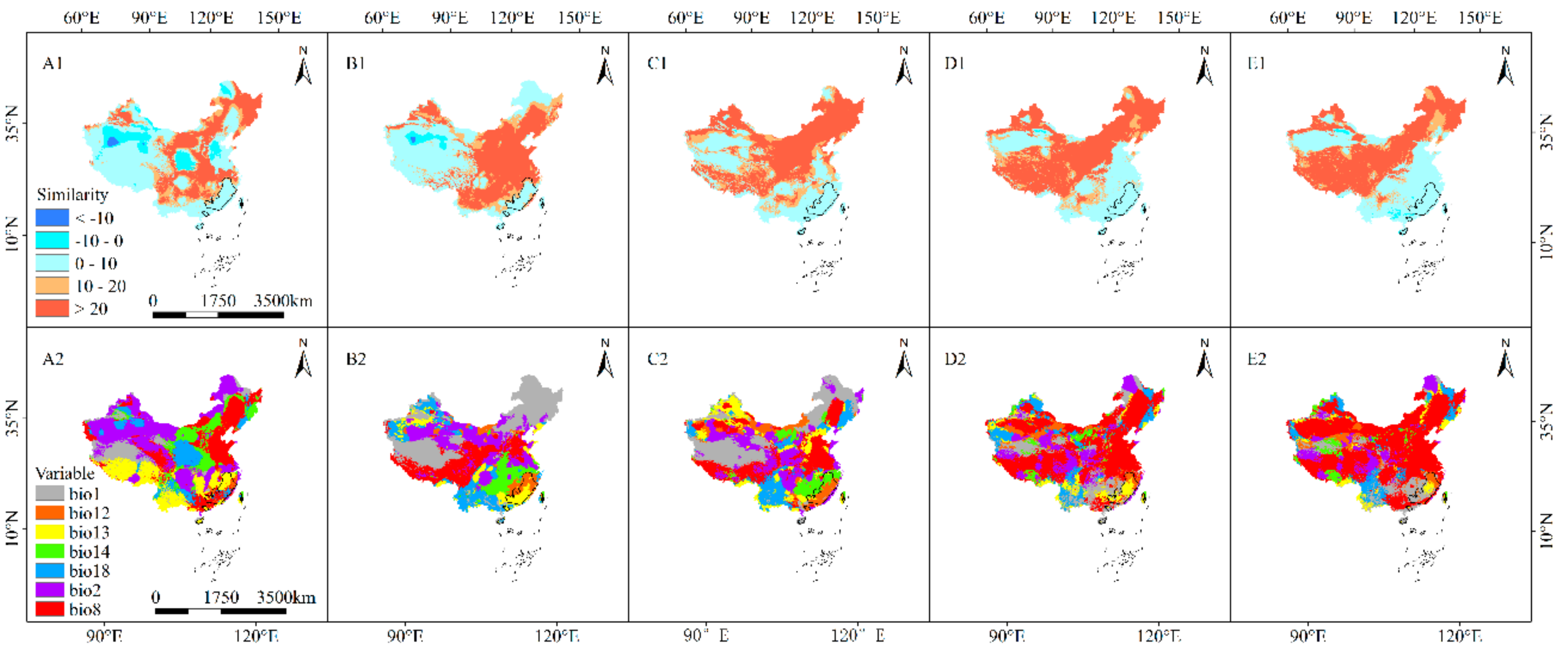
3.7. MESS and MoD Variable Analysis
3.8. Shift in the Distribution Center of the Suitable Habitat
4. Discussion
4.1. Bioclimatic Predictors and Model Performance
4.2. Changes in Potential Distribution Areas
4.3. Refugia from Climate Change
4.4. Restriction of Climate Variables on Geographical Distribution
4.5. Protection of Genetic Resources
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Waldvogel, A.M.; Feldmeyer, B.; Rolshausen, G.; Exposito-Alonso, M.; Rellstab, C.; Kofler, R.; Mock, T.; Schmid, K.; Schmitt, I.; Bataillon, T.; et al. Evolutionary genomics can improve prediction of species’ responses to climate change. Evol. Lett. 2020, 4, 4–18. [Google Scholar] [CrossRef] [PubMed]
- Warren, R.; Price, J.; VanDerWal, J.; Cornelius, S.; Sohl, H. The implications of the United Nations Paris Agreement on climate change for globally significant biodiversity areas. Clim. Chang. 2018, 147, 395–409. [Google Scholar] [CrossRef]
- Pecl, G.T.; Araujo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengard, B.; et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, 9. [Google Scholar] [CrossRef]
- Fackovcova, Z.; Senko, D.; Svitok, M.; Guttova, A. Ecological niche conservatism shapes the distributions of lichens: Geographical segregation does not reflect ecological differentiation. Preslia 2017, 89, 63–85. [Google Scholar] [CrossRef]
- Martinez-Freiria, F.; Velo-Anton, G.; Brito, J.C. Trapped by climate: Interglacial refuge and recent population expansion in the endemic Iberian adder Vipera seoanei. Divers. Distrib. 2015, 21, 331–344. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Zhu, R.W.; Zhong, D.L.; Zhang, J.Q. Nunataks or massif de refuge? A phylogeographic study of Rhodiola crenulata (Crassulaceae) on the world’s highest sky islands. BMC Evol. Biol. 2018, 18, 154. [Google Scholar] [CrossRef]
- Cubry, P.; De Bellis, F.; Pot, D.; Musoli, P.; Leroy, T. Global analysis of coffea canephora pierre ex froehner (Rubiaceae) from the guineo-congolese region reveals impacts from climatic refugia and migration effects. Genet. Resour. Crop Evol. 2013, 60, 483–501. [Google Scholar] [CrossRef]
- Garcia, R.A.; Cabeza, M.; Rahbek, C.; Araujo, M.B. Multiple dimensions of climate change and their implications for biodiversity. Science 2014, 344, 486. [Google Scholar] [CrossRef]
- Blois, J.L.; Zarnetske, P.L.; Fitzpatrick, M.C.; Finnegan, S. Climate change and the past, present, and future of biotic interactions. Science 2013, 341, 499–504. [Google Scholar] [CrossRef]
- Bennett, K.D.; Provan, J. What do we mean by ‘refugia’? Quat. Sci. Rev. 2008, 27, 2449–2455. [Google Scholar] [CrossRef]
- Thomas, C.D. Climate, climate change and range boundaries. Divers. Distrib. 2010, 16, 488–495. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, X.W.; Fang, Y.M. Landscape features and climatic forces shape the genetic structure and evolutionary history of an oak species (quercus chenii) in East China. Front. Plant Sci. 2019, 10, 1060. [Google Scholar] [CrossRef] [PubMed]
- Walas, L.; Sobierajska, K.; Ok, T.; Donmez, A.A.; Kanoglu, S.S.; Dagher-Kharrat, M.B.; Douaihy, B.; Romo, A.; Stephan, J.; Jasinska, A.K.; et al. Past, present, and future geographic range of an oro-Mediterranean Tertiary relict: The juniperus drupacea case study. Reg. Envir. Chang. 2019, 19, 1507–1520. [Google Scholar] [CrossRef]
- Gristwood, A. Red lists, green lists and conservation an interview with Thomas Brooks, chief scientist, international union for the conservation of nature. EMBO Rep. 2020, 21, 4. [Google Scholar] [CrossRef]
- Ho, H.C.; Chan, T.C.; Xu, Z.W.; Huang, C.; Li, C. Individual-and community-level shifts in mortality patterns during the January 2016 East Asia cold wave associated with a super El Nino event: Empirical evidence in Hong Kong. Sci. Total Environ. 2020, 711, 135050. [Google Scholar] [CrossRef]
- Hu, T.; Sun, Y. Projected changes in extreme warm and cold temperatures in China from 1.5 to 5 °C global warming. Int. J. Climatol. 2019, 3. [Google Scholar] [CrossRef]
- Ha, Y.; Zhong, Z.; Zhang, Y.; Ding, J.F.; Yang, X.R. Relationship between interannual changes of summer rainfall over Yangtze River Valley and South China Sea-Philippine Sea: Possible impact of tropical zonal sea surface temperature gradient. Int. J. Climatol. 2019, 39, 5522–5538. [Google Scholar] [CrossRef]
- Schipper, A.M.; Posthuma, L.; de Zwart, D.; Huijbregts, M.A.J. Deriving field-based species sensitivity distributions (f-SSDs) from stacked species distribution models (S-SDMs). Environ. Sci. Technol. 2014, 48, 14464–14471. [Google Scholar] [CrossRef]
- Prieto-Torres, D.A.; Navarro-Siguenza, A.G.; Santiago-Alarcon, D.; Rojas-Soto, O.R. Response of the endangered tropical dry forests to climate change and the role of Mexican Protected Areas for their conservation. Glob. Change Biol. 2016, 22, 364–379. [Google Scholar] [CrossRef]
- La Marca, W.; Elith, J.; Firth, R.S.C.; Murphy, B.P.; Regan, T.J.; Woinarski, J.C.Z.; Nicholson, E. The influence of data source and species distribution modelling method on spatial conservation priorities. Divers. Distrib. 2019, 25, 1060–1073. [Google Scholar] [CrossRef]
- Benito, B.M.; Martinez-Ortega, M.M.; Munoz, L.M.; Lorite, J.; Penas, J. Assessing extinction-risk of endangered plants using species distribution models: A case study of habitat depletion caused by the spread of greenhouses. Biodivers. Conserv. 2009, 18, 2509–2520. [Google Scholar] [CrossRef]
- Pecchi, M.; Marchi, M.; Burton, V.; Giannetti, F.; Moriondo, M.; Bernetti, I.; Bindi, M.; Chirici, G. Species distribution modelling to support forest management. A literature review. Ecol. Model. 2019, 411, 1–12. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudik, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Baldwin, R.A. Use of maximum entropy modeling in wildlife research. Entropy 2009, 11, 854–866. [Google Scholar] [CrossRef]
- Liu, H.Y.; Qi, X.Z.; Gong, H.B.; Li, L.H.; Zhang, M.Y.; Li, Y.F.; Lin, Z.S. Combined effects of global climate suitability and regional environmental variables on the distribution of an invasive marsh species spartina alterniflora. Estuaries Coasts 2009, 42, 99–111. [Google Scholar] [CrossRef]
- Adhikari, D.; Barik, S.K.; Upadhaya, K. Habitat distribution modelling for reintroduction of Ilex khasiana Purk., a critically endangered tree species of northeastern India. Ecol. Eng. 2012, 40, 37–43. [Google Scholar] [CrossRef]
- Qin, A.; Liu, B.; Guo, Q.; Bussmann, R.W.; Ma, F.; Jian, Z.; Xu, G.; Pei, S. Maxent modeling for predicting impacts of climate change on the potential distribution of Thuja sutchuenensis Franch., an extremely endangered conifer from southwestern China. Glob. Ecol. Conserv. 2017, 10, 139–146. [Google Scholar] [CrossRef]
- Huang, L.L.; Jin, J.H.; Quan, C.; Oskolski, A.A. Mummified Magnoliaceae woods from the upper Oligocene of South China, with biogeography, paleoecology, and wood trait evolution implications. J. Syst. Evol. 2020, 58, 89–100. [Google Scholar] [CrossRef]
- Zhang, M.Z.; Jiang, Y.T.; Ye, X.Z.; Chen, S.P.; Fan, H.H.; Liu, B. The complete chloroplast genome of Semiliquidambar cathayensis (Hamamelidaceae). Mitochondrial DNA Part B Resour. 2020, 5, 695–696. [Google Scholar] [CrossRef]
- Yagi, H.; Xu, J.; Moriguchi, N.; Miyagi, R.; Moritsuka, E.; Sato, E.; Sugai, K.; Setsuko, S.; Torimaru, T.; Yamamoto, S.; et al. Population genetic analysis of two species of Distylium: D. racemosum growing in East Asian evergreen broad-leaved forests and D. lepidotum endemic to the Ogasawara (Bonin) Islands. Tree Genet. Genomes 2019, 15, 77. [Google Scholar] [CrossRef]
- Sun, J.Z.; Zheng, X.L.; Cui, X.Z.; Meng, L. A study of inhibition effects on hepatitis B virus of Semi-cathayensis in vitro. Lishizhen Med. Mater. Med. Res. 2014, 25, 2391–2393. [Google Scholar] [CrossRef]
- Li, W.; Shi, M.; Huang, Y.; Chen, K.; Sun, H.; Chen, J. Climatic change can influence species diversity patterns and potential habitats of salicaceae plants in China. Forests 2019, 10, 220. [Google Scholar] [CrossRef]
- Shi, Y.C.; Duan, N.; Liu, B.B. Complete chloroplast genome sequence of Semiliquidambar cathayensis (Hamamelidaceae), a rare and endangered species endemic to China. Mitochondrial DNA Part B Resour. 2019, 4, 3252–3253. [Google Scholar] [CrossRef]
- Wu, W.; Zhou, R.C.; Huang, Y.L.; Boufford, D.E.; Shi, S.H. Molecular evidence for natural intergeneric hybridization between Liquidambar and Altingia. J. Plant Res. 2010, 123, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhao, Z.-G.; Liu, Q.; Chen, B.; Cheng, X.-Y. A New Record of the National Secondary Protection Species—Semiliquidambar cathayensis H. T. Chang in Sichuan Province. J. Sichuan For. Sci. Technol. 2018, 39, 63–65. [Google Scholar] [CrossRef]
- Global Biodiversity Information Facility. Available online: https://www.gbif.org (accessed on 2 February 2020).
- Specimen Resources Sharing Platform for Education. Available online: http://mnh.scu.edu.cn/main.aspx (accessed on 2 February 2020).
- Chinese herbarium of nature. Available online: http://www.cfh.ac.cn (accessed on 2 February 2020).
- Chinese Virtual Herbarium. Available online: http://www.cvh.org.cn (accessed on 2 February 2020).
- Zhang, K.; Yao, L.; Meng, J.; Tao, J. Maxent modeling for predicting the potential geographical distribution of two peony species under climate change. Sci. Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef]
- Oldfather, M.F.; Kling, M.M.; Sheth, S.N.; Emery, N.C.; Ackerly, D.D. Range edges in heterogeneous landscapes: Integrating geographic scale and climate complexity into range dynamics. Glob. Chang. Biol. 2019, 26, 1055–1067. [Google Scholar] [CrossRef]
- Liao, Z.; Zhang, L.; Nobis, M.P.; Wu, X.; Pan, K.; Wang, K.; Dakhil, M.A.; Du, M.; Xiong, Q.; Pandey, B.; et al. Climate change jointly with migration ability affect future range shifts of dominant fir species in Southwest China. Divers. Distrib. 2019, 26, 1–16. [Google Scholar] [CrossRef]
- World Clim. Global Climate Data Free Climate Data for Ecological Modeling and GIS. Available online: http://worldclim.org (accessed on 2 February 2020).
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Harris, I.; Jones, P.D.; Osborn, T.J.; Lister, D.H. Updated high-resolution grids of monthly climatic observations-the CRU TS3.10 Dataset. Int. J. Climatol. 2014, 34, 623–642. [Google Scholar] [CrossRef]
- Radosavljevic, A.; Anderson, R.P. Making better MAXENT models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Ranjitkar, S.; Xu, J.; Shrestha, K.K.; Kindt, R. Ensemble forecast of climate suitability for the Trans-Himalayan Nyctaginaceae species. Ecol. Model. 2014, 282, 18–24. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudik, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for MAXENT ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Steen, B.; Cardoso, A.C.; Tsiamis, K.; Nieto, K.; Engel, J.; Gervasini, E. Modelling hot spot areas for the invasive alien plant Elodea nuttallii in the EU. Manag. Biol. Invasions 2019, 10, 151–170. [Google Scholar] [CrossRef]
- Guevara, L.; Gerstner, B.E.; Kass, J.M.; Anderson, R.P. Toward ecologically realistic predictions of species distributions: A cross-time example from tropical montane cloud forests. Glob. Chang. Biol. 2018, 24, 1511–1522. [Google Scholar] [CrossRef]
- Velasco, J.A.; Gonzalez-Salazar, C. Akaike information criterion should not be a “test” of geographical prediction accuracy in ecological niche modelling. Ecol. Inform. 2019, 51, 25–32. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Kurpis, J.; Serrato-Cruz, M.A.; Arroyo, T.P.F. Modeling the effects of climate change on the distribution of Tagetes lucida Cav. (Asteraceae). Glob. Ecol. Conserv. 2019, 20, 11. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, X.W.; Fang, Y.M. Responses of the distribution pattern of Quercus chenii to climate change following the Last Glacial Maximum. Chin. J. Plant Ecol. 2016, 40, 1164–1178. [Google Scholar] [CrossRef]
- Dhyani, S.; Kadaverugu, R.; Dhyani, D.; Verma, P.; Pujari, P. Predicting impacts of climate variability on habitats of Hippophae salicifolia (D. Don) (Seabuckthorn) in Central Himalayas: Future challenges. Ecol. Inform. 2018, 48, 135–146. [Google Scholar] [CrossRef]
- Elith, J.; Kearney, M.; Phillips, S. The art of modelling range-shifting species. Methods In Ecol. Evolution 2010, 1, 330–342. [Google Scholar] [CrossRef]
- Li, Y.; Cao, W.; He, X.Y.; Chen, W.; Xu, S. Prediction of suitable habitat for lycophytes and ferns in northeast China: A case study on athyrium brevifrons. Chin. Geogr. Sci. 2019, 29, 1011–1023. [Google Scholar] [CrossRef]
- Yi, Y.J.; Cheng, X.; Yang, Z.F.; Zhang, S.H. Maxent modeling for predicting the potential distribution of endangered medicinal plant (H. riparia Lour) in Yunnan, China. Ecol. Eng. 2016, 92, 260–269. [Google Scholar] [CrossRef]
- Wei, J.F.; Li, X.Z.; Lu, Y.Y.; Zhao, L.; Zhang, H.F.; Zhao, Q. Modeling the potential global distribution of phenacoccus madeirensis green under various climate change scenarios. Forests 2019, 10, 773. [Google Scholar] [CrossRef]
- Sony, R.K.; Sen, S.; Kumar, S.; Sen, M.; Jayahari, K.M. Niche models inform the effects of climate change on the endangered Nilgiri Tahr (Nilgiritragus hylocrius) populations in the southern Western Ghats, India. Ecol. Eng. 2018, 120, 355–363. [Google Scholar] [CrossRef]
- Xiang, X.G.; Xiang, K.L.; Ortiz, R.D.; Jabbour, F.; Wang, W. Integrating palaeontological and molecular data uncovers multiple ancient and recent dispersals in the pantropical Hamamelidaceae. J. Biogeogr. 2019, 46, 1–12. [Google Scholar] [CrossRef]
- Maslova, N.P.; Kodrul, T.M.; Song, Y.S.; Volkova, L.D.; Jin, J.H. Liquidambar maomingensis sp nov (Altingiaceae) from the late Eocene of South China. Am. J. Bot. 2015, 102, 1356–1370. [Google Scholar] [CrossRef]
- Jiang, X.L.; Xu, G.B.; Deng, M. Spatial genetic patterns and distribution dynamics of the rare oak quercus chungii: Implications for biodiversity conservation in Southeast China. Forests 2019, 10, 13. [Google Scholar] [CrossRef]
- Pellissier, L.; Eidesen, P.B.; Ehrich, D.; Descombes, P.; Schonswetter, P.; Tribsch, A.; Westergaard, K.B.; Alvarez, N.; Guisan, A.; Zimmermann, N.E.; et al. Past climate-driven range shifts and population genetic diversity in arctic plants. J. Biogeogr. 2016, 43, 461–470. [Google Scholar] [CrossRef]
- Wang, S.; Lu, H.; Han, J.; Chu, G.; Liu, J.; Negendank, J.F.W. Palaeovegetation and palaeoclimate in low-latitude southern China during the Last Glacial Maximum. Quat. Int. 2012, 248, 79–85. [Google Scholar] [CrossRef]
- Napier, J.D.; Fernandez, M.C.; de Lafontaine, G.; Hu, F.S. Ice-age persistence and genetic isolation of the disjunct distribution of larch in Alaska. Ecol. Evol. 2020, 10, 1692–1702. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Shen, Z.; Ying, L.; Zang, R.; Jiang, Y. Effects of current climate, paleo-climate, and habitat heterogeneity in determining biogeographical patterns of evergreen broad-leaved woody plants in China. J. Geogr. Sci. 2019, 29, 1142–1158. [Google Scholar] [CrossRef]
- Waltari, E.; Hijmans, R.J.; Peterson, A.T.; Nyari, A.S.; Perkins, S.L.; Guralnick, R.P. Locating pleistocene refugia: Comparing phylogeographic and ecological Niche model predictions. PloS ONE 2007, 2, 11. [Google Scholar] [CrossRef]
- Feng, G.; Mao, L.; Sandel, B.; Swenson, N.G.; Svenning, J.-C. High plant endemism in China is partially linked to reduced glacial-interglacial climate change. J. Biogeogr. 2016, 43, 145–154. [Google Scholar] [CrossRef]
- Tian, Z.P.; Jiang, D.B. Mid Holocene and last glacial maximum changes in monsoon are and precipitation over China (in Chinese). Chin. Sei. Bull. 2015, 60, 400–410. [Google Scholar] [CrossRef]
- Meng, Y.H.; Xu, X.; Jing, X.L.; Xu, G.B. Potential distribution modeling and analysis of Disanthus Maxim. Actae Ecol. Sin. 2018, 39, 2816–2825. [Google Scholar] [CrossRef]
- Morrison, B.D.; Heath, K.; Greenberg, J.A. Spatial scale affects novel and disappeared climate change projections in Alaska. Ecol. Evol. 2019, 9, 12026–12044. [Google Scholar] [CrossRef]
- Tian, B.; Fu, Y.; Milne, R.I.; Mao, K.S.; Sun, Y.S.; Ma, X.G.; Sun, H. A complex pattern of post-divergence expansion, contraction, introgression, and asynchronous responses to Pleistocene climate changes in two Dipelta sister species from western China. J. Syst. Evol. 2019, 59, 1–16. [Google Scholar] [CrossRef]
- Das, S.; Baumgartner, J.B.; Esperon-Rodriguez, M.; Wilson, P.D.; Yap, J.Y.S.; Rossetto, M.; Beaumont, L.J. Identifying climate refugia for 30 Australian rainforest plant species, from the last glacial maximum to 2070. Landsc. Ecol. 2019, 34, 2883–2896. [Google Scholar] [CrossRef]
- Zhao, H.R.; Hua, L.T.; Hong, Y.L.; Tao, L.; De, Q.P. Analysis of the Endangered causes of Semiliquidambar cathayensis H.T. Chang in Guizhou Province. Guizhou For. Sci. Technol. 2014, 42, 34–36. [Google Scholar]
- Shen, Y.M.; Jin, L.H.; Zhi, X.T.; Hou, L.W.; Wen, Q.T.; Jian, H.C. Studies on the rare, endangered and national key protected plants at Tianxin Natrual Reserves of Lianzhou, Guangdong. J. Guangzhou Univ. (Nat. Sci. Ed.) 2013, 12, 29–34. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Environmental Variable | Unit | Percent Contribution | Permutation Importance |
|---|---|---|---|---|
| bio1 | Annual mean temperature | 10 °C | 37.3 | 25.9 |
| bio2 | Mean diurnal range | 10 °C | 23 | 8.2 |
| bio12 | Annual precipitation | mm | 17.8 | 25.1 |
| bio14 | Precipitation of driest month | mm | 10.9 | 10 |
| bio8 | Mean temperature of wettest quarter | 10 °C | 6.6 | 13.4 |
| bio18 | Precipitation of warmest quarter | mm | 2.6 | 6.6 |
| bio13 | Precipitation of wettest month | mm | 1.9 | 10.9 |
| Type | FC | RM | OR10 | Δ AICc | Mean AUC | avg.diff.AUC |
|---|---|---|---|---|---|---|
| default | LQPHT | 1 | 0.16473 | 53.545 | 0.8855 | 0.020149 |
| optimized | LQ | 1.5 | 0.135415 | 0 | 0.9388 | 0.0198 |
| Item | LIG | LGM | MH | 2050s | 2070s | ||
|---|---|---|---|---|---|---|---|
| RCP2.6 | RCP8.5 | RCP2.6 | RCP8.5 | ||||
| Unchanged rate | 2.33% | 79.65% | 91.78% | 57.98% | 69.13% | 90.81% | 46.41% |
| Contraction rate | −1.12% | −27.53% | −40.81% | −41.76% | −31.32% | −9.04% | −53.36% |
| Expansion rate | 97.75% | 20.31% | 7.85% | 5.48% | 24.27% | 31.64% | 9.04% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, X.-z.; Zhao, G.-h.; Zhang, M.-z.; Cui, X.-y.; Fan, H.-h.; Liu, B. Distribution Pattern of Endangered Plant Semiliquidambar cathayensis (Hamamelidaceae) in Response to Climate Change after the Last Interglacial Period. Forests 2020, 11, 434. https://doi.org/10.3390/f11040434
Ye X-z, Zhao G-h, Zhang M-z, Cui X-y, Fan H-h, Liu B. Distribution Pattern of Endangered Plant Semiliquidambar cathayensis (Hamamelidaceae) in Response to Climate Change after the Last Interglacial Period. Forests. 2020; 11(4):434. https://doi.org/10.3390/f11040434
Chicago/Turabian StyleYe, Xing-zhuang, Guang-hua Zhao, Ming-zhu Zhang, Xin-yue Cui, Hui-hua Fan, and Bao Liu. 2020. "Distribution Pattern of Endangered Plant Semiliquidambar cathayensis (Hamamelidaceae) in Response to Climate Change after the Last Interglacial Period" Forests 11, no. 4: 434. https://doi.org/10.3390/f11040434
APA StyleYe, X.-z., Zhao, G.-h., Zhang, M.-z., Cui, X.-y., Fan, H.-h., & Liu, B. (2020). Distribution Pattern of Endangered Plant Semiliquidambar cathayensis (Hamamelidaceae) in Response to Climate Change after the Last Interglacial Period. Forests, 11(4), 434. https://doi.org/10.3390/f11040434