Twenty-Five Years after Stand Thinning and Repeated Fertilization in Lodgepole Pine Forest: Implications for Tree Growth, Stand Structure, and Carbon Sequestration

Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Study Areas
2.2. Density and Fertilization Treatments
2.3. Growth and Size of Crop Trees
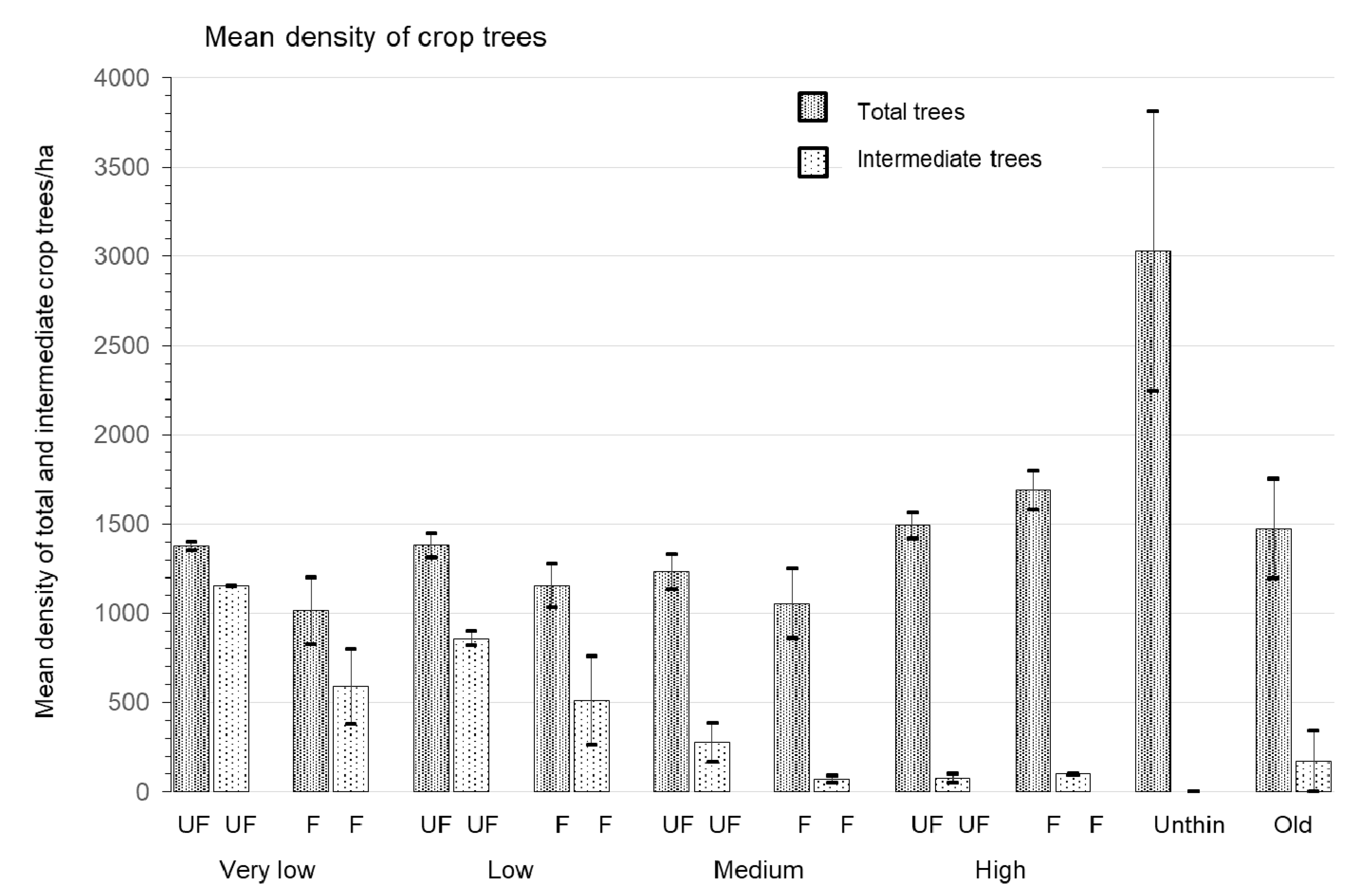
2.4. Stand Structure Measurements
2.5. Estimating Stand-Level Carbon Storage
2.6. Estimating Biomass for Herbs and Shrubs
2.7. Statistical Analysis
3. Results
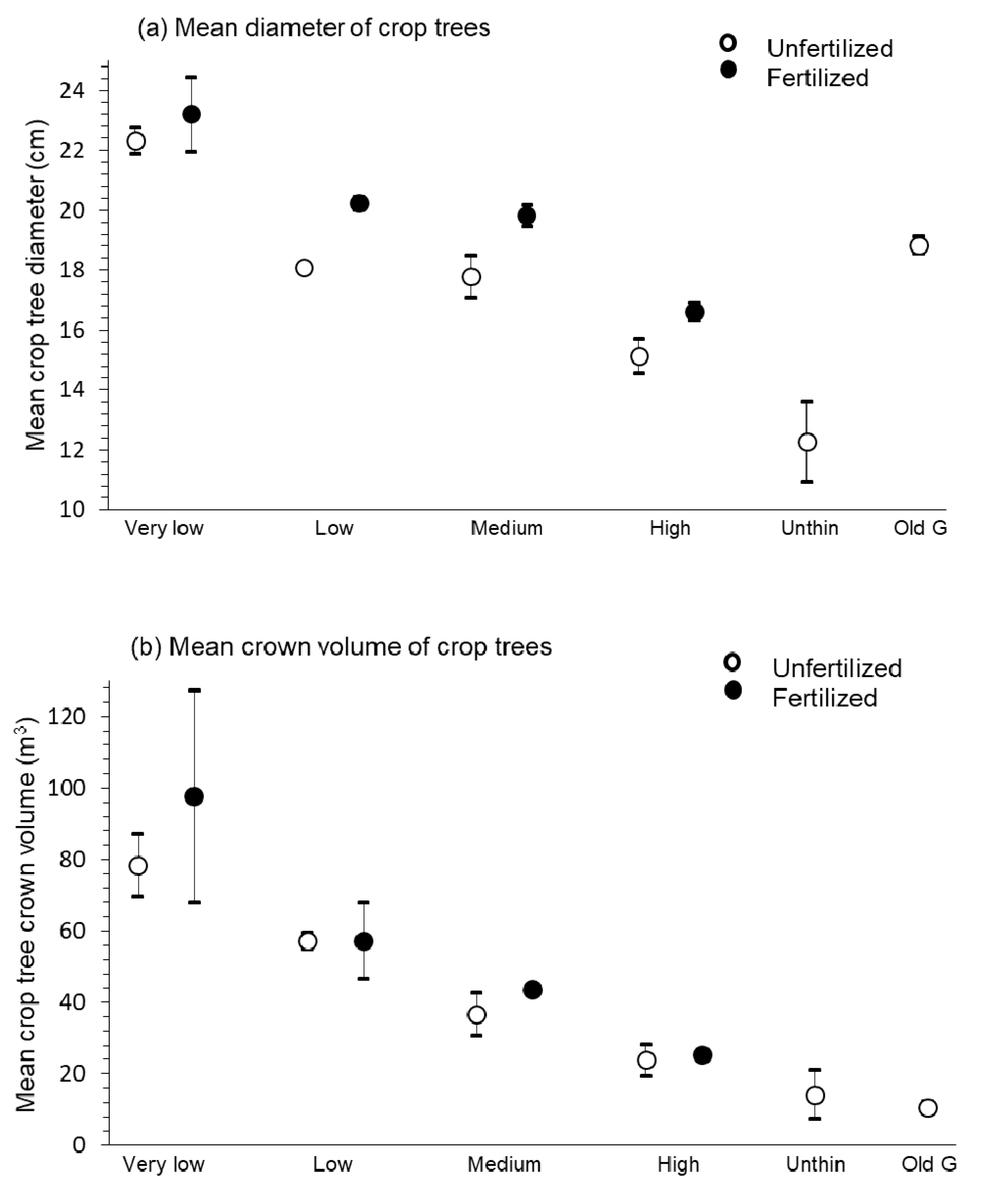
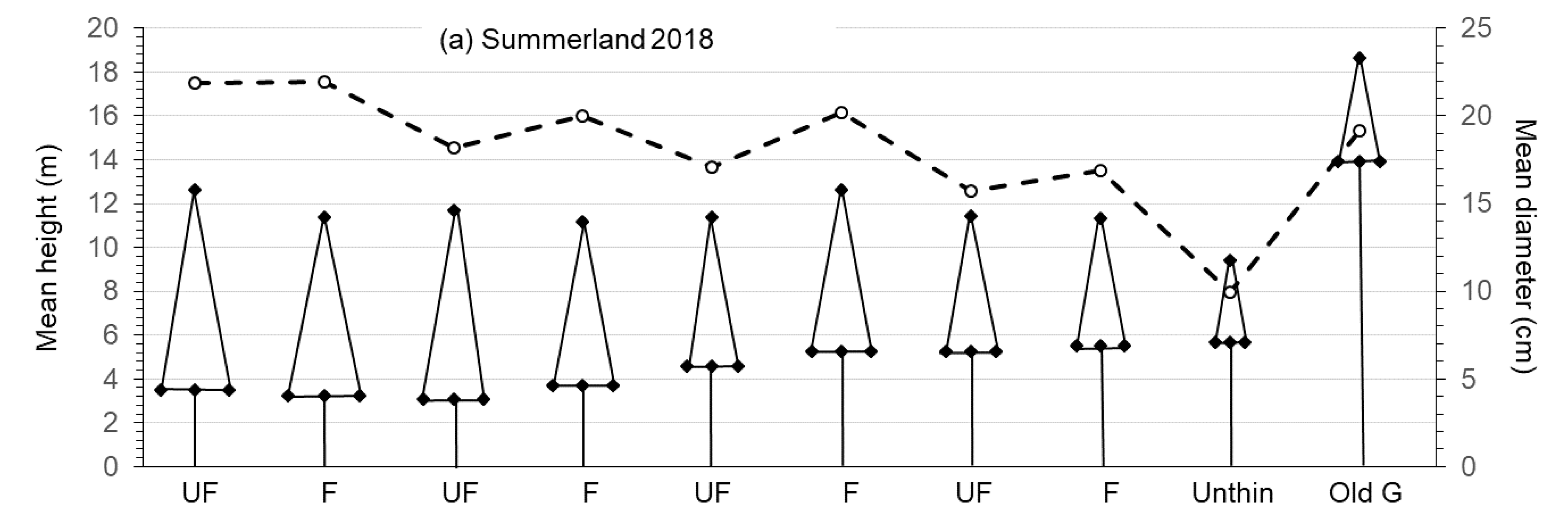
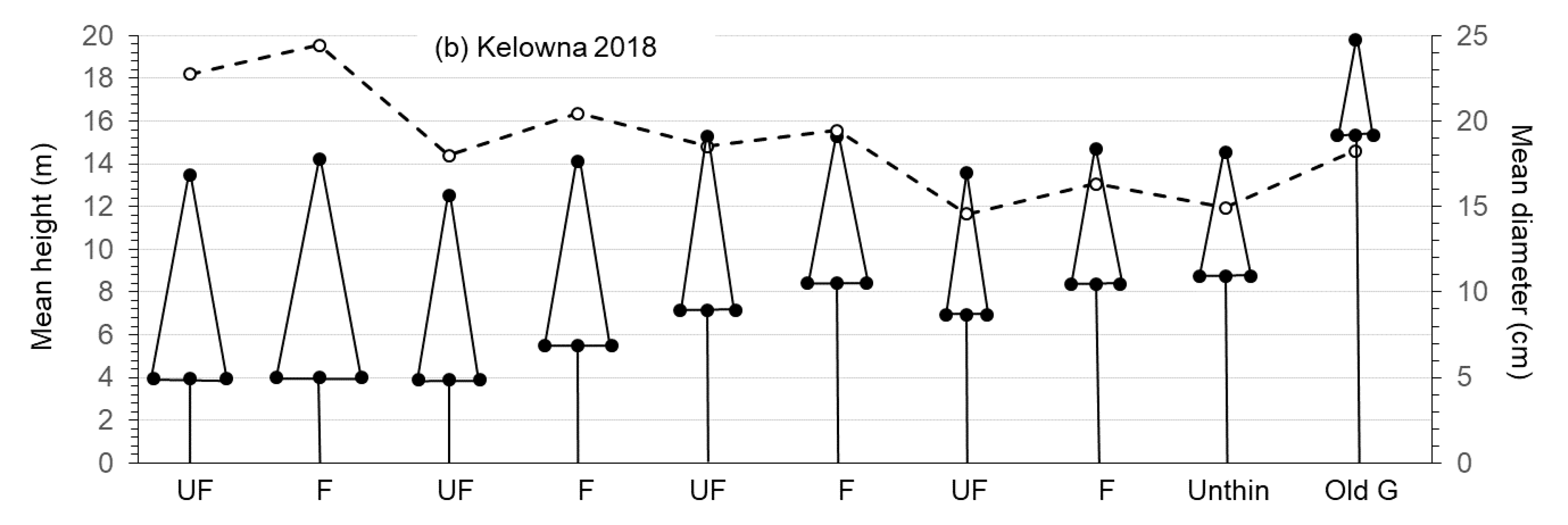
3.1. Growth and Size of Crop Trees
3.2. Architecture of Tree Crowns
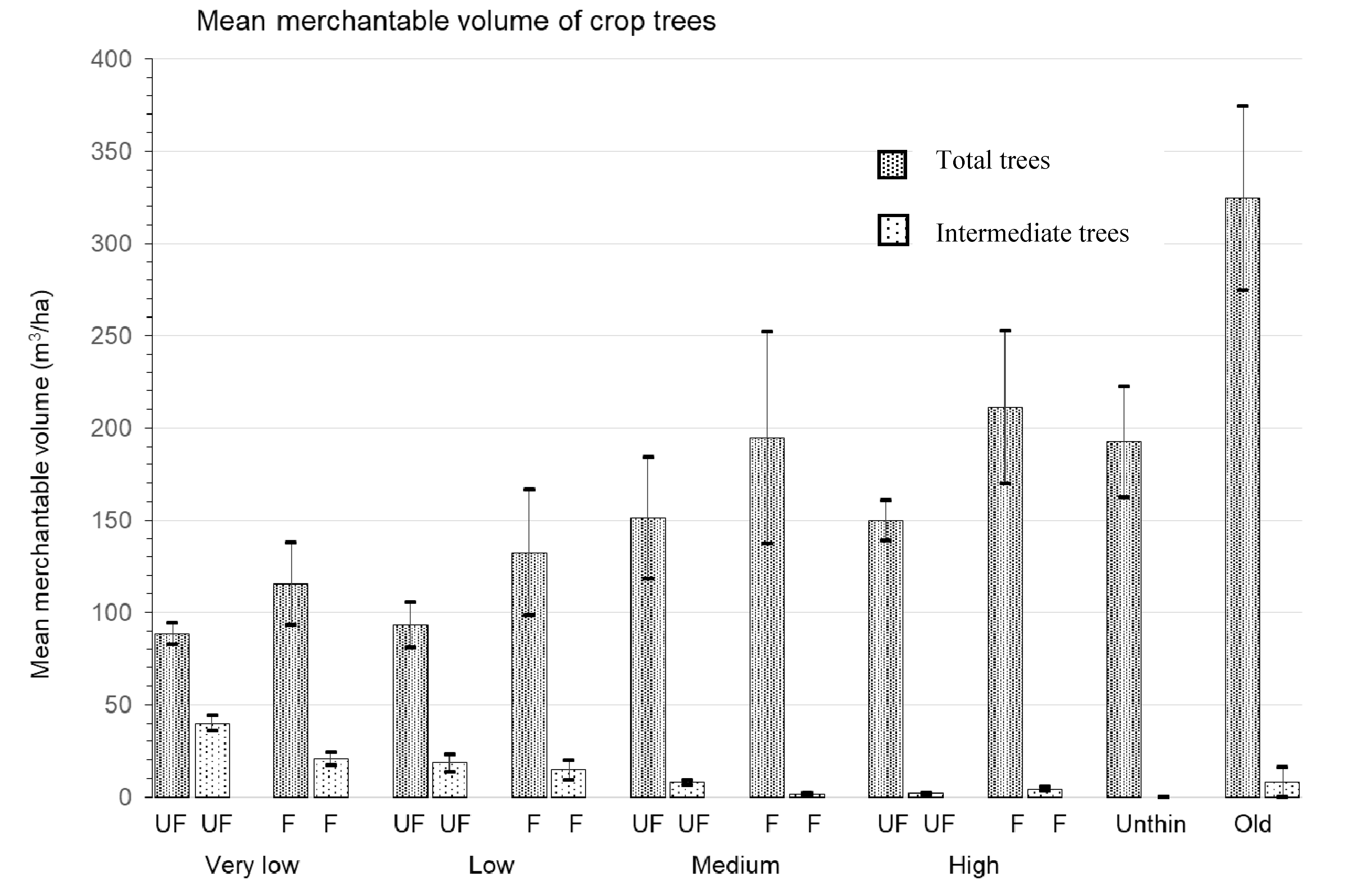
3.3. Merchantable Volume
3.4. Understory Conifers and Vascular Plants
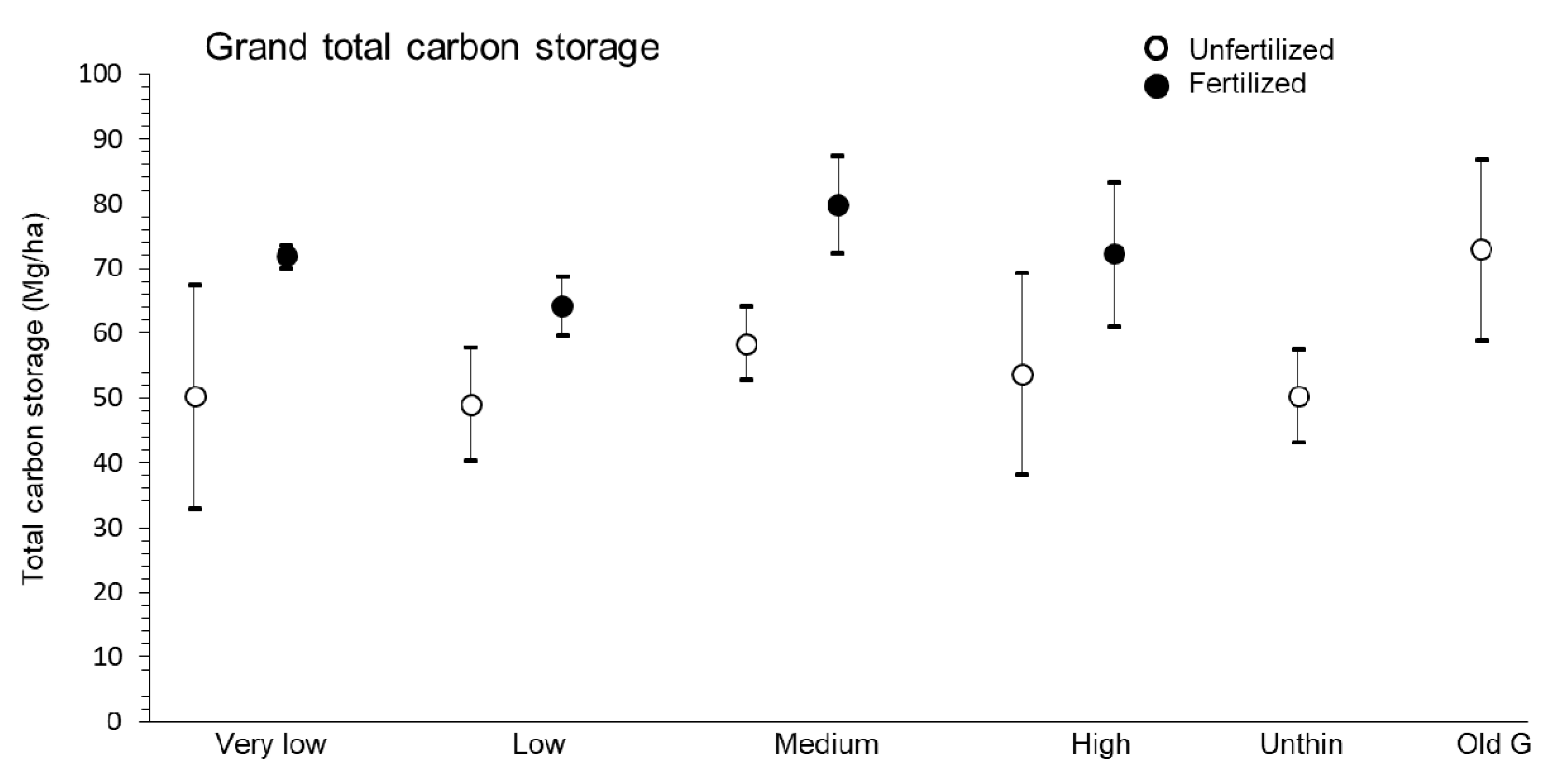
3.5. Stand-Level Carbon Storage
4. Discussion
4.1. Growth and Size of Crop Trees
4.2. Architecture of Tree Crowns
4.3. Merchantable Volume
4.4. Stand Thinning and Ecosystem Services
4.5. Understory Conifer, Herb, and Shrub Layers
4.6. Stand-Level Carbon Storage
4.7. Study Limitations
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nabuurs, G.J.; Pussinen, A.; van Brusselen, J.; Schelhaas, M.J. Future harvesting pressure on European forests. Eur. J. For. Res. 2007, 126, 391–400. [Google Scholar] [CrossRef]
- Raunikar, R.; Buongiornoa, J.; Turnerb, J.A.; Zhu, S. Global outlook for wood and forests with the bioenergy demand implied by scenarios of the Intergovernmental Panel on Climate Change. For. Pol. Econ. 2010, 12, 48–56. [Google Scholar] [CrossRef]
- Hunter, M.L.; Schmiegelow, F.K.A. Wildlife, Forests, and Forestry, 2nd ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2011; 259p. [Google Scholar]
- Agee, J.K. Fire Ecology of Pacific Northwest Forests; Island Press: Washington, DC, USA, 1993. [Google Scholar]
- Walton, A.; Hughes, J.; Eng, M.; Fall, A.; Shore, T.; Riel, B.; Hall, P. Provincial –level Projection of the Current Mountain Pine Beetle Outbreak; British Columbia Ministry of Forests and Range: Victoria, BC, Canada, 2009. [Google Scholar]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; Mcdowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Lavoie, S.; Ruel, J.-C.; Bergeron, Y.; Harvey, B.D. Windthrow after group and dispersed tree retention in eastern Canada. For. Ecol. Manag. 2012, 269, 158–167. [Google Scholar] [CrossRef]
- Moore, S.E.; Allen, H.L. Plantation Forestry. In Maintaining Biodiversity in Forest Ecosystems; Hunter, M.L., Jr., Ed.; Cambridge University Press: New York, NY, USA, 1999; pp. 400–433. [Google Scholar]
- Albaugh, T.J.; Allen, H.L.; Dougherty, P.M.; Johnsen, K.H. Long term growth responses of loblolly pine to optimal nutrient and water resource availability. For. Ecol. Manag. 2004, 192, 3–19. [Google Scholar] [CrossRef]
- Nohrstedt, H.O. Response of coniferous forest ecosystems on mineral soils to nutrient additions: A review of Swedish experiences. Scand. J. For. Res. 2001, 16, 555–573. [Google Scholar] [CrossRef]
- Bergh, J.; Nilsson, U.; Grip, H.; Hedwall, P.O.; Lundmark, T. Effects of frequency of fertilization on production, foliar chemistry, and nutrient leaching in young Norway spruce stands in Sweden. Silva. Fenn. 2008, 42, 721–733. [Google Scholar] [CrossRef]
- Fox, T.R.; Jokela, E.A.; Allen, H.L. The development of pine plantation silviculture in the southern United States. J. For. 2007, 105, 337–347. [Google Scholar]
- Jokela, E.J.; Martin, T.A.; Vogel, J.G. Twenty-five years of intensive management with southern pines: Important lessons learned. J. For. 2010, 108, 338–347. [Google Scholar]
- Riffell, S.; Verschuyl, J.; Miller, D.; Wigley, T.B. Biofuel harvests, coarse woody debris, and biodiversity–A meta-analysis. For. Ecol. Manag. 2011, 261, 878–887. [Google Scholar] [CrossRef]
- Verschuyl, J.; Riffell, S.; Miller, D.; Wigley, T.B. Biodiversity response to intensive biomass production from forest thinning in North American forests—A meta-analysis. For. Ecol. Manag. 2011, 261, 221–232. [Google Scholar] [CrossRef]
- Demarais, S.; Verschuyl, J.P.; Roloff, G.J.; Miller, D.A.; Wigley, T.B. Tamm review: Terrestrial vertebrate biodiversity and intensive forest management in the U.S. For. Ecol. Manag. 2017, 385, 308–330. [Google Scholar] [CrossRef]
- Cole, D.M.; Koch, P. Managing Lodgepole Pine to Yield Merchantable Thinning Products and Attain Sawtimber Rotations; Research Paper INT-RP-482; USDA Forest Service Intermountain Research Station: Ogden, UT, USA, 1996. [Google Scholar]
- Koch, P. Lodgepole Pine Commercial Forests: An Essay Comparing the Natural Cycle of Insect Kill and Subsequent Wildfire with Management for Utilization and Wildlife; INT-GTR-342; USDA Forest Service Intermountain Research Station: Ogden, UT, USA, 1996. [Google Scholar]
- Lindgren, P.M.F.; Sullivan, T.P. Long-term responses of tree and stand growth of young lodgepole pine to pre-commercial thinning and repeated fertilization. For. Ecol. Manag. 2013, 307, 155–164. [Google Scholar] [CrossRef]
- Lotan, J.E.; Critchfield, W.B. Pinus contorta ssp. murrayana–lodgepole pine forest. In Silvics of North America, Volume 1. Conifers. Agriculture Handbook 654; Burns, R.M., Honkala, B.H., Eds.; Tech. Coord. USDA Forest Service: Washington, DC, USA, 1990; pp. 302–313. [Google Scholar]
- Koch, P. Lodgepole Pine in North America; Forest Production Society: Madison, WI, USA, 1996. [Google Scholar]
- Brockley, R.P.; Trowbridge, R.L.; Ballard, T.M.; Macadam, A.M. Nutrient Management in Interior Forest Types. Proceedings Forest Fertilization: Sustaining and Improving Nutrition and Growth of Western Forests, Seattle, WA, USA, 12–14 February 1991; University of Washington Institute of Forest Resources Contribution 73: Seattle, WA, USA, 12 February 1992; pp. 43–64. [Google Scholar]
- Brockley, R.P. Effects of post-thinning density and repeated fertilization on the growth and development of young lodgepole pine. Can. J. For. Res. 2005, 35, 1952–1964. [Google Scholar] [CrossRef]
- Lindgren, P.M.F.; Sullivan, T.P.; Sullivan, D.S.; Brockley, R.P.; Winter, R. Growth response of young lodgepole pine to thinning and repeated fertilization treatments: 10-year results. Forestry 2007, 80, 587–611. [Google Scholar] [CrossRef]
- Malkonen, E.; Kukkola, M. Effect of long-term fertilization on the biomass production and nutrient status of Scots pine stands. Fertilizer Res. 1991, 27, 113–127. [Google Scholar] [CrossRef]
- Tamm, C.O.; Aronsson, A.; Popovic, B.; Flower-Ellis, J. Optimum Nutrition and Nitrogen Saturation in Scots Pine Stands; Studia Forestalia Suecica: Uppsala, Sweden, 1999; p. 206. [Google Scholar]
- Thysell, D.R.; Carey, A.B. Manipulation of density of Pseudotsuga menziesii canopies: Preliminary effects on understory vegetation. Can. J. For. Res. 2001, 31, 1513–1525. [Google Scholar] [CrossRef]
- Sullivan, T.P.; Sullivan, D.S.; Lindgren, P.M.F. Stand structure and small mammals in young lodgepole pine forest: 10-year results after thinning. Ecol. Appl. 2001, 11, 1151–1173. [Google Scholar] [CrossRef]
- Lindgren, P.M.F.; Ransome, D.B.; Sullivan, D.S.; Sullivan, T.P. Plant community attributes 12 to 14 years following pre-commercial thinning in a young lodgepole pine forest. Can. J. For. Res. 2006, 36, 48–61. [Google Scholar] [CrossRef]
- Cole, E.C.; Hanley, T.A.; Newton, M. Influence of precommercial thinning on understory vegetation of young-growth Sitka spruce forests in southeastern Alaska. Can. J. For. Res. 2010, 40, 619–628. [Google Scholar] [CrossRef]
- Thomas, S.C.; Halpern, C.B.; Falk, D.A.; Liguori, D.A.; Austin, K.A. Plant diversity in managed forests: Understorey responses to thinning and fertilization. Ecol. Appl. 1999, 9, 864–879. [Google Scholar] [CrossRef]
- Lindgren, P.M.F.; Sullivan, T.P. Influence of stand thinning and repeated fertilization on plant community abundance and diversity in young lodgepole pine stands: 15-year results. For. Ecol. Manag. 2013, 308, 17–30. [Google Scholar] [CrossRef]
- Sullivan, T.P.; Sullivan, D.S.; Lindgren, P.M.F.; Ransome, D.B. Stand structure and small mammals in intensively managed forests: Scale, time, and testing extremes. For. Ecol. Manag. 2013, 310, 1071–1087. [Google Scholar] [CrossRef]
- Ter-Mikaelian, M.T.; Colombo, S.J.; Chen, J. Fact and fantasy about forest carbon. For. Chron. 2008, 84, 166–171. [Google Scholar] [CrossRef]
- Hedwall, P.-O.; Gong, P.; Ingerslev, M.; Bergh, J. Fertilization in northern forests–biological, economic and environmental constraints and possibilities. Scand. J. For. Res. 2014, 29, 301–311. [Google Scholar] [CrossRef]
- Birdsey, R.A. Carbon Storage and Accumulation in United States Forest Ecosystems; GGeneral Technical Report WO-59; USDA Forest Service: Washington, DC, USA, 1992. [Google Scholar]
- Luyssaert, S.; Schultze, E.-D.; Borner, A.; Knohl, A.; Hessenmoller, D.; Law, B.E.; Clais, P.; Grace, J. Old-growth forests as global carbon sinks. Nature 2008, 455, 213–215. [Google Scholar] [CrossRef]
- Ishii, H.R.; Sillett, S.C.; Caroll, A.L. Crown dynamics and wood production of Douglas-fir trees in an old-growth forest. For. Ecol. Manag. 2017, 384, 157–168. [Google Scholar] [CrossRef]
- Adams, A.B.; Harrison, R.B.; Sletton, R.S.; Strahm, B.D.; Turnblom, E.C.; Jensen, C.M. Nitrogen-fertilization impacts on carbon sequestration and flux in managed coastal Douglas-fir stands of the Pacific Northwest. For. Ecol. Manag. 2005, 220, 313–325. [Google Scholar] [CrossRef]
- Shryock, B.; Littke, K.; Ciol, M.; Briggs, D.; Harrison, R. The effects of urea fertilization on carbon sequestration in Douglas-fir plantations of the coastal Pacific Northwest. For. Ecol. Manag. 2014, 318, 341–348. [Google Scholar] [CrossRef]
- Marchi, M.; Paletto, A.; Cantiani, P.; Bianchetto, E.; De Meo, I. Comparing thinning system effects on ecosystem services provision in artificial black pine (Pinus nigra J.F. Arnold) forests. Forests 2018, 9, 188. [Google Scholar] [CrossRef]
- Sullivan, T.P.; Sullivan, D.S. Acceleration of old-growth structural attributes in lodgepole pine forest: Tree growth and stand structure 25 years after thinning. For. Ecol. Manag. 2016, 365, 96–106. [Google Scholar] [CrossRef]
- Blevins, D.P.; Prescott, C.E.; Allen, H.L.; Newsome, T.A. The effects of nutrition and density on growth, foliage biomass, and growth efficiency of high-density fire-origin lodgepole pine in central British Columbia. Can. J. For. Res. 2005, 35, 2851–2859. [Google Scholar] [CrossRef]
- Schmidt, W.C.; Seidel, K.W. Western larch and space: Thinning to optimize growth. In Proceedings of the Future Forests of the Mountain West: A Stand Culture Symposium, Missoula, MT, USA, 29 September–3 October 1986; General Technical Report INT-243. U.S. Forest Service, Intermountain Research Station: Missoula, MT, USA, 1988. [Google Scholar]
- Bailey, J.D.; Tappeiner, J.C. Effects of thinning on structural development in 40- to 100-year-old Douglas-fir stands in western Oregon. For. Ecol. Manag. 1998, 108, 99–113. [Google Scholar] [CrossRef]
- Newton, M.; Cole, L. Overstory development in Douglas-fir-dominant forests thinned to enhance late-seral features. For. Sci. 2015, 61, 809–816. [Google Scholar] [CrossRef]
- Meidinger, D.; Pojar, J. Ecosystems of British Columbia; Special Report Series No. 6; Research Branch, Ministry of Forests: Victoria, BC. Canada, 1991. [Google Scholar]
- Nigh, G.D. Total and Merchantable Volume Equations for Common Tree Species in British Columbia: By Region and Biogeoclimatic Zone; Technical Report 106, British Columbia Province; Region and Biogeoclimatic Zone: Victoria, BC, Canada, 2016; 22p. [Google Scholar]
- Englund, S.R.; O’Brien, J.J.; Clark, D.B. Evaluation of digital and film hemispherical photography and spherical densiometry for measuring forest light environments. Can. J. For. Res. 2000, 30, 1999–2005. [Google Scholar] [CrossRef]
- Parish, R.; Coupe, R.; Lloyd, D. Plants of Southern Interior British Columbia; Lone Pine Publishing: Vancouver, BC, Canada, 1996; 464p. [Google Scholar]
- Bert, D.; Danjon, F. Carbon concentration variation in the roots, stem and crown of mature Pinus pinaster (Ait.). For. Ecol. Manag. 2006, 222, 279–295. [Google Scholar] [CrossRef]
- Ter-Mikaelian, M.T.; Korzukhin, M.D. Biomass equations for sixty-five North American tree species. For. Ecol. Manag. 1997, 97, 1–24. [Google Scholar] [CrossRef]
- Li, Z.; Kurz, W.A.; Apps, M.J.; Beukema, S.J. Belowground biomass dynamics in the Carbon Budget Model of the Canadian Forest Sector: Recent improvements and implications for the estimation of NPP and NEP. Can. J. For. Res. 2003, 33, 126–136. [Google Scholar] [CrossRef]
- Jain, T.; Graham, R.T.; Adams, D. Carbon concentrations and carbon pool distributions in dry, moist, and cold mid-aged forests of the Rocky Mountains. In Integrated management of carbon sequestration and biomass utilization opportunities in a changing climate; Proceedings of the 2009 National Silviculture Workshop, Fort Collins, CO, USA, 15–18 June 2009; Jain, T.B., Graham, R.T., Sandquist, J., Eds.; Boise, ID. Proceedings RMRS-P-61; Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2010; pp. 39–59. [Google Scholar]
- IBM Corp. Inc. IBM SPSS Statistics for Windows; Version 25.0; IBM Corp: Armonk, NY, USA, 2017. [Google Scholar]
- Fowler, J.; Cohen, L.; Jarvis, P. Practical Statistics for Field Biology, 2nd ed.; John Wiley and Sons: Chichester, UK, 1998; 259p. [Google Scholar]
- Saville, D.J. Multiple comparison procedures: The practical solution. Amer. Stat. 1990, 44, 174–180. [Google Scholar]
- Zar, J.H. Biostatistical Analysis; Prentice-Hall, Inc.: Englewood Cliffs, NJ, USA, 1999; 663p. [Google Scholar]
- Sullivan, T.P.; Sullivan, D.S. Old-growth characteristics 20 years after thinning and repeated fertilization of lodgepole pine forest: Tree growth, structural attributes, and red-backed voles. For. Ecol. Manag. 2017, 391, 207–220. [Google Scholar] [CrossRef]
- Hess, A.F.; da Silveira, A.C.; Krefta, S.M.; dos Santo, D.V.; Filho, M.D.; Atanzio, K.A.; Schorr, L.P.B.; dos Santo, I.A.; Borsoi, G.A.; Stepka, T.F.; et al. Crown dynamics of Brazilian pine (Aracauria angustifolia) in Santa Catarina region of Brazil. Austr. J. Crop Sci. 2018, 12, 449–457. [Google Scholar] [CrossRef]
- Millar, C.I.; Stephenson, N.L.; Stephens, S.L. Climate change and forests of the future: Managing in the face of uncertainty. Ecol. Appl. 2007, 17, 2145–2151. [Google Scholar] [CrossRef] [PubMed]
- D’Amato, A.W.; Bradford, J.B.; Fraver, S.; Palik, B.J. Effects of thinning on drought variability and climate response in north temperate forest ecosystems. Ecol. Appl. 2013, 23, 1735–1742. [Google Scholar]
- Sohn, J.A.; Saha, S.; Bauhus, J. Potential of forest thinning to mitigate drought stress: A meta-analysis. For. Ecol. Manag. 2016, 380, 261–273. [Google Scholar] [CrossRef]
- Vacek, S.; Vacek, Z.; Bilek, L.; Simon, J.; Remes, J.; Hunova, I.; Kral, J.; Putalova, T.; Mikeska, M. Structure, regeneration and growth of Scots pine (Pinus sylvestris L.) stands with respect to changing climate and environmental pollution. Silva Fenn 2016, 50. [Google Scholar] [CrossRef]
- Podolak, K.; Edelson, D.; Kruse, S.; Aylward, B.; Zimring, M.; Wobbrock, N. Estimating the Water Supply Benefits from Forest Restoration in the Northern Sierra Nevada; The Nature Conservancy and Ecosystem Economics: San Francisco, CA, USA, 2015. [Google Scholar]
- Stephens, S.L.; McIver, J.D.; Boerner, R.E.J.; Fettig, C.J.; Fontaine, J.B.; Hartsough, B.R.; Kennedy, P.L.; Schwilk, D.W. The effects of forest fuel reduction treatments in the United States. BioScience 2012, 62, 549–560. [Google Scholar] [CrossRef]
- Amman, G.D.; McGregor, M.D.; Schmitz, R.F.; Oakes, R.D. Susceptibility of lodgepole pine to infestation by mountain pine beetles following partial cutting of stands. Can. J. For. Res. 1988, 18, 688–695. [Google Scholar] [CrossRef]
- Whitehead, R.J.; Russo, G.L. “Beetle-Proofed” Lodgepole Pine Stands in interior British Columbia Have Less Damage from Mountain Pine Beetle; Report BCX-402; Natural Resources Canada, Canadian Forest Service, Pacific Forestry Centre: Victoria, BC, Canada, 2005; 18p. [Google Scholar]
- Widenfalk, O.; Weslien, J. Plant species richness in managed boreal forests – Effects of stand succession and thinning. For. Ecol. Manag. 2009, 257, 1386–1394. [Google Scholar] [CrossRef]
- Lindh, B.C.; Muir, P.S. Understory vegetation in young Douglas-fir forests: Does thinning help restore old-growth composition? For. Ecol. Manag. 2004, 192, 285–296. [Google Scholar] [CrossRef]
- Sathre, R.; Gustavsson, L.; Bergh, J. Primary energy and greenhouse gas implications of increasing biomass production through forest fertilization. Biomass Bioenergy 2010, 34, 572–581. [Google Scholar] [CrossRef]
- Albaugh, T.J.; Vance, E.D.; Gaudreault, C.; Fox, T.R.; Allen, H.L.; Stape, J.L.; Rubilar, R.A. Carbon emissions and sequestration from fertilization of pine in the southeastern United States. For. Sci. 2012, 58, 419–429. [Google Scholar] [CrossRef]
- Hoen, H.F.; Solberg, B. Potential and economic efficiency of carbon sequestration in forest biomass through management. For. Sci. 1994, 40, 429–451. [Google Scholar]
- Huettl, R.F.; Zoettl, H.W. Forest fertilization: Its potential to increase the CO2 storage capacity and to alleviate the decline of the global forests. Water Air Soil Poll. 1992, 64, 229–249. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Density (stems/ha) | Pine Crop Trees (Mean ± SE) | ||||||
|---|---|---|---|---|---|---|---|
| Study Area and Stand | Original Pine Crop Trees 1 | Crop Trees Other Species | Total Intermediate Crop Trees 2 | Total Crop Trees All Species 3 | DBH (cm) | Height (m) | Total Canopy Closure (%) |
| PCT study | |||||||
| Penticton | |||||||
| Low | 550 | 95 | 315 | 960 | 21.8 ± 0.3 | 13.9 ± 0.1 | 87.4 ± 1.3 |
| Medium | 1190 | 240 | 190 | 1620 | 18.7 ± 0.2 | 15.4 ± 0.1 | 89.8 ± 0.7 |
| High | 1670 | 15 | 0 | 1685 | 16.9 ± 0.2 | 14.7 ± 0.1 | 92.3 ± 0.5 |
| Unthinned | 4462 | 40 | 0 | 4502 | 10.6 ± 0.3 | 11.7 ± 0.2 | 88.9 ± 1.4 |
| Old-growth | 790 | 360 | 0 | 1150 | 19.3 ± 0.5 | 21.0 ± 0.3 | 87.3 ± 1.7 |
| Summerland | |||||||
| Low | 495 | 0 | 820 | 1315 | 18.2 ± 0.4 | 11.7 ± 0.2 | 88.6 ± 1.5 |
| Medium | 935 | 7 | 389 | 1331 | 17.1 ± 0.2 | 11.4 ± 0.1 | 85.7 ± 1.1 |
| High | 1306 | 59 | 54 | 1419 | 15.7 ± 0.4 | 11.4 ± 0.2 | 85.6 ± 2.1 |
| Unthinned | 2636 | 120 | 0 | 2756 | 11.3 ± 0.2 | 9.9 ± 0.1 | 89.0 ± 1.2 |
| Old-growth | 1480 | 40 | 510 | 2030 | 19.2 ± 0.7 | 18.7 ± 0.7 | 86.5 ± 1.8 |
| Kelowna | |||||||
| Low | 550 | 0 | 900 | 1450 | 18.0 ± 2.4 | 12.5 ± 0.4 | 94.3 ± 0.5 |
| Medium | 800 | 167 | 166 | 1133 | 18.5 ± 0.5 | 15.3 ± 0.2 | 87.2 ± 3.2 |
| High | 1467 | 0 | 100 | 1567 | 14.6 ± 0.5 | 13.6 ± 0.1 | 92.1 ± 2.6 |
| Unthinned | 1600 | 233 | 0 | 1833 | 14.9 ± 1.0 | 14.5 ± 0.3 | 92.6 ± 1.2 |
| Old-growth 4 | 1243 | 0 | 0 | 1243 | 18.3 ± 0.5 | 19.8 ± 0.6 | 80.5 ± 1.4 |
| PCT-FERT study | |||||||
| Summerland | |||||||
| Very low | 210 | 40 | 1155 | 1405 | 21.9 ± 0.3 | 12.6 ± 0.2 | 81.6 ± 3.0 |
| Very low F | 265 | 180 | 380 | 825 | 22.0 ± 0.3 | 11.4 ± 0.1 | 80.4 ± 3.1 |
| Low F | 505 | 15 | 760 | 1280 | 20.0 ± 0.4 | 11.2 ± 0.2 | 91.4 ± 1.1 |
| Medium F | 745 | 27 | 90 | 862 | 20.2 ± 0.4 | 12.6 ± 0.3 | 89.0 ± 2.3 |
| High F | 1330 | 156 | 95 | 1581 | 16.9 ± 0.4 | 11.3 ± 0.2 | 91.4 ± 1.1 |
| Kelowna | |||||||
| Very low | 200 | 0 | 1150 | 1350 | 22.8 ± 0.6 | 13.5 ± 1.7 | 80.9 ± 3.5 |
| Very low F | 350 | 50 | 800 | 1200 | 24.5 ± 0.2 | 14.2 ± 0.4 | 88.8 ± 1.8 |
| Low F | 700 | 67 | 266 | 1033 | 20.4 ± 1.0 | 14.1 ± 1.3 | 93.9 ± 1.6 |
| Medium F | 1200 | 0 | 50 | 1250 | 19.5 ± 0.1 | 15.3 ± 0.2 | 93.2 ± 1.6 |
| High F | 1700 | 0 | 100 | 1800 | 16.3 ± 1.1 | 14.7 ± 0.5 | 93.5 ± 0.5 |
| Stand Attribute | Treatment | ANOVA | |||||
|---|---|---|---|---|---|---|---|
| Low | Medium | High | Unthinned | Old-Growth | F4,8 | p | |
| DBH (cm) (Original pine crop trees) | 19.33 ± 1.23ab | 18.09 ± 0.50ab | 15.76 ± 0.70b | 12.27 ± 1.35c | 18.84 ± 0.29ab | 9.29 | <0.01 |
| Height (m) (Original pine crop trees) | 12.70 ± 0.64b | 14.00 ± 1.30b | 13.23 ± 0.96b | 12.05 ± 1.34b | 19.72 ± 0.59a | 26.29 | <0.01 |
| Pine crop trees/ha | 531.7 ± 18.3b | 975.0 ± 114.3b | 1481.0 ± 105.3b | 2899.3 ± 836.6a | 1171.0 ± 202.4b | 5.29 | 0.02 |
| Other species of crop trees/ha | 31.7 ± 31.7 | 138.0 ± 68.8 | 24.7 ± 17.7 | 131.0 ± 56.0 | 133.3 ± 113.9 | 0.78 | 0.57 |
| Total intermediate crop trees/ha | 678.3 ± 183.1a | 248.3 ± 70.7b | 51.3 ± 28.9b | 0 ± 0b | 170.0 ± 170.0b | 6.99 | 0.01 |
| Total crop trees/ha | 1241.7 ± 146.1 | 1361.3 ± 141.4 | 1557.0 ± 77.0 | 3030.3 ± 782.6 | 1474.3 ± 279.1 | 3.51 | 0.06 |
| Canopy closure (%) | 90.1 ± 2.1 | 87.6 ± 1.2 | 90.0 ± 2.2 | 90.2 ± 1.2 | 84.8 ± 2.1 | 1.54 | 0.28 |
| Crown volume (m3/tree) | 67.75 ± 10.47a | 33.52 ± 4.73b | 26.19 ± 3.49b | 14.24 ± 6.74b | 10.74 ± 1.70b | 11.26 | <0.01 |
| Total merchantable volume (m3/ha) | 113.3 ± 21.4b | 195.5 ± 48.4ab | 185.7 ± 36.4b | 192.4 ± 30.2ab | 324.6 ± 49.9a | 3.76 | 0.053 |
| Total basal area (m2/ha) | 19.4 ± 1.9 | 26.9 ± 3.5 | 29.6 ± 4.1 | 31.2 ± 4.1 | 34.0 ± 7.0 | 1.45 | 0.30 |
| Understory conifers (trees/ha) | 2138 ± 209b | 1420 ± 158b | 1343 ± 429b | 2621 ± 649b | 17535 ± 6644a | 5.80 | 0.02 |
| Abundance of herbs (m3/0.01 ha) | 5.45 ± 1.13a | 5.37 ± 0.80a | 3.92 ± 0.82a | 3.52 ± 1.49ab | 0.42 ± 0.11b | 4.17 | 0.04 |
| Abundance of shrubs (m3/0.01 ha) | 29.87 ± 16.24 | 14.98 ± 2.53 | 10.14 ± 4.64 | 17.02 ± 11.88 | 8.77 ± 1.63 | 1.24 | 0.37 |
| ANOVA | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Density | Fertilizer | Density × Fertilizer | ||||||||
| Mean Values | F3,8 | p | F1,8 | p | F3,8 | p | ||||
| DBH (cm) (Original pine crop trees) | very low | low | medium | high | 44.80 | <0.01 | 14.93 | <0.01 | 0.46 | 0.72 |
| 22.76 ± 0.60a | 19.16 ± 0.62b | 18.81 ± 0.67b | 15.88 ± 0.50c | |||||||
| unfertilized | fertilized | |||||||||
| 18.33 ± 0.99b | 19.97 ± 0.92a | |||||||||
| Height (m) (Original pine crop trees) | very low | low | medium | high | 0.32 | 0.81 | 0.14 | 0.72 | 0.05 | 0.99 |
| 12.93 ± 0.61 | 12.38 ± 0.65 | 13.65 ± 0.98 | 12.76 ± 0.84 | |||||||
| unfertilized | fertilized | |||||||||
| 12.76 ± 0.47 | 13.10 ± 0.60 | |||||||||
| Pine crop trees/ha | very low | low | medium | high | 38.61 | <0.01 | 1.59 | 0.24 | 0.01 | 0.99 |
| 256 ± 34d | 562 ± 47c | 920 ± 101b | 1451 ± 90a | |||||||
| unfertilized | fertilized | |||||||||
| 745 ± 167 | 849 ± 180 | |||||||||
| Other species of crop trees/ha | very low | low | medium | high | 0.33 | 0.80 | 0.65 | 0.44 | 1.09 | 0.41 |
| 67 ± 39 | 21 ± 16 | 51 ± 39 | 54 ± 37 | |||||||
| unfertilized | fertilized | |||||||||
| 34 ± 21 | 62 ± 25 | |||||||||
| Total intermediate crop trees | very low | low | medium | high | 19.60 | <0.01 | 10.01 | 0.01 | 2.00 | 0.19 |
| 871 ± 183a | 687 ± 143a | 174 ± 76b | 87 ± 11b | |||||||
| unfertilized | fertilized | |||||||||
| 592 ± 165a | 318 ± 108b | |||||||||
| Total crop trees | very low | low | medium | high | 5.37 | 0.03 | 2.69 | 0.14 | 1.91 | 0.21 |
| 1195 ± 131b | 1270 ± 87b | 1144 ± 102b | 1592 ± 78a | |||||||
| unfertilized | fertilized | |||||||||
| 1371 ± 44 | 1229 ± 119 | |||||||||
| Canopy closure (%) | very low | low | medium | high | 5.33 | 0.03 | 3.61 | 0.09 | 0.20 | 0.89 |
| 82.9 ± 2.0b | 92.1 ± 1.3a | 88.8 ± 1.6ab | 90.7 ± 1.7a | |||||||
| unfertilized | fertilized | |||||||||
| 87.0 ± 1.6 | 90.2 ± 1.6 | |||||||||
| Crown volume (m3/tree) | very low | low | medium | high | 10.44 | <0.01 | 0.66 | 0.44 | 0.27 | 0.85 |
| 88.04 ± 13.76a | 57.32 ± 4.50b | 40.06 ± 3.19bc | 24.61 ± 1.99c | |||||||
| unfertilized | fertilized | |||||||||
| 49.09 ± 8.16 | 55.93 ± 11.69 | |||||||||
| Total merch volume (m3/ha) | very low | low | medium | high | 3.23 | 0.08 | 3.64 | 0.09 | 0.10 | 0.96 |
| 102.0 ± 12.2 | 112.8 ± 18.7 | 172.9 ± 29.9 | 180.60 ± 24.9 | |||||||
| unfertilized | fertilized | |||||||||
| 120.6 ± 13.3 | 163.5 ± 21.7 | |||||||||
| Total BA/ha (m2) | very low | low | medium | high | 7.76 | <0.01 | 7.29 | 0.03 | 0.51 | 0.69 |
| 17.46 ± 1.74c | 20.34 ± 1.83bc | 26.78 ± 3.04ab | 29.50 ± 2.62a | |||||||
| unfertilized | fertilized | |||||||||
| 20.82 ± 1.45b | 26.23 ± 2.70a | |||||||||
| Understory conifers (trees/ha) | very low | low | medium | high | 1.69 | 0.25 | 3.28 | 0.11 | 0.47 | 0.71 |
| 3388 ± 683 | 3046 ± 400 | 2283 ± 285 | 2546 ± 411 | |||||||
| unfertilized | fertilized | |||||||||
| 3252 ± 259 | 2379 ± 345 | |||||||||
| Herbs (grasses+forbs) (m3/0.01ha) | very low | low | medium | high | 1.13 | 0.40 | 0.38 | 0.55 | 0.31 | 0.82 |
| 5.80 ± 0.59 | 5.03 ± 0.90 | 5.81 ± 0.78 | 3.96 ± 0.59 | |||||||
| unfertilized | fertilized | |||||||||
| 5.40 ± 0.66 | 4.89 ± 0.41 | |||||||||
| Shrubs (m3/0.01ha) | very low | low | medium | high | 4.16 | 0.05 | 0.01 | 0.93 | 1.22 | 0.36 |
| 22.98 ± 3.09a | 12.33 ± 0.92b | 16.57 ± 1.33ab | 9.82 ± 4.34b | |||||||
| unfertilized | fertilized | |||||||||
| 15.56 ± 2.89 | 15.29 ± 2.33 | |||||||||
| Stand Attribute | Treatment | ANOVA | |||||
|---|---|---|---|---|---|---|---|
| Low | Medium | High | Unthinned | Old-Growth | F4,8 | p | |
| Pine crop trees | 63.64 ± 24.62 | 61.74 ± 12.64 | 67.40 ± 18.36 | 41.37 ± 6.51 | 60.15 ± 14.51 | 0.53 | 0.72 |
| Other species of crop trees | 3.72 ± 3.72 | 9.17 ± 5.91 | 3.03 ± 1.90 | 2.81 ± 1.66 | 6.16 ± 4.72 | 0.84 | 0.54 |
| Total intermediate crop trees | 5.25 ± 0.79 | 2.70 ± 0.14 | 0.35 ± 0.18 | 0 ± 0 | 2.32 ± 2.32 | 3.44 | 0.07 |
| Total crop trees | 72.62 ± 27.89 | 73.61 ± 17.39 | 70.77 ± 20.08 | 44.18 ± 6.84 | 68.63 ± 15.22 | 0.66 | 0.64 |
| Understory conifers | 2.77 ± 0.86abc | 1.57 ± 0.28bc | 0.71 ± 0.21c | 5.94 ± 0.68a | 4.15 ± 1.65ab | 4.47 | 0.03 |
| Total conifers | 75.39 ± 27.03 | 75.18 ± 17.24 | 71.48 ± 19.99 | 50.12 ± 7.13 | 72.78 ± 14.05 | 0.54 | 0.71 |
| Grasses | 0.07 ± 0.01 | 0.06 ± 0.02 | 0.06 ± 0.01 | 0.06 ± 0.01 | 0.03 ± 0.00 | 1.94 | 0.20 |
| Herbs (grasses+forbs) | 0.12 ± 0.01 | 0.11 ± 0.01 | 0.10 ± 0.01 | 0.10 ± 0.02 | 0.06 ± 0.001 | 3.31 | 0.07 |
| Shrubs | 0.09 ± 0.04 | 0.06 ± 0.01 | 0.04 ± 0.01 | 0.06 ± 0.03 | 0.04 ± 0.004 | 1.33 | 0.34 |
| Grand total | 75.61 ± 27.06 | 75.35 ± 17.24 | 71.62 ± 20.02 | 50.28 ± 7.13 | 72.89 ± 14.04 | 0.54 | 0.71 |
| ANOVA | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Density | Fertilizer | Density × Fertilizer | ||||||||
| Mean Values | F3,8 | p | F1,8 | p | F3,8 | p | ||||
| Pine crop trees | very low | low | medium | high | 3.44 | 0.07 | 11.03 | 0.01 | 0.23 | 0.88 |
| 39.55 ± 6.80 | 47.37 ± 6.11 | 63.40 ± 8.85 | 58.34 ± 7.66 | |||||||
| unfertilized | fertilized | |||||||||
| 42.55 ± 5.17b | 61.79 ± 4.34a | |||||||||
| Other species of crop trees | very low | low | medium | high | 1.44 | 0.30 | 2.47 | 0.16 | 1.61 | 0.26 |
| 5.71 ± 2.62 | 0.97 ± 0.59 | 2.33 ± 1.56 | 2.85 ± 2.09 | |||||||
| unfertilized | fertilized | |||||||||
| 1.66 ± 0.88 | 4.27 ± 1.61 | |||||||||
| Total intermediate crop trees | very low | low | medium | high | 12.71 | <0.01 | 1.81 | 0.22 | 1.31 | 0.34 |
| 11.10 ± 2.35a | 5.55 ± 1.30b | 1.72 ± 0.67bc | 0.95 ± 0.25c | |||||||
| unfertilized | fertilized | |||||||||
| 5.70 ± 2.08 | 3.95 ± 1.31 | |||||||||
| Total crop trees | very low | low | medium | high | 0.80 | 0.53 | 8.72 | 0.02 | 0.03 | 0.99 |
| 56.36 ± 8.58 | 53.89 ± 6.44 | 67.46 ± 7.46 | 62.14 ± 9.42 | |||||||
| unfertilized | fertilized | |||||||||
| 49.91 ± 4.75b | 70.01 ± 3.49a | |||||||||
| Understory conifers | very low | low | medium | high | 3.05 | 0.09 | 0.99 | 0.35 | 0.21 | 0.89 |
| 4.45 ± 1.49 | 2.55 ± 0.63 | 1.51 ± 0.32 | 0.71 ± 0.14 | |||||||
| unfertilized | fertilized | |||||||||
| 2.76 ± 0.87 | 1.84 ± 0.58 | |||||||||
| Total conifers | very low | low | medium | high | 0.51 | 0.69 | 6.89 | 0.03 | 0.04 | 0.99 |
| 60.81 ± 9.45 | 56.44 ± 5.97 | 68.96 ± 7.27 | 62.85 ± 9.46 | |||||||
| unfertilized | fertilized | |||||||||
| 52.68 ± 5.01b | 71.85 ± 3.42a | |||||||||
| Grasses | very low | low | medium | high | 0.46 | 0.72 | 0.59 | 0.46 | 0.64 | 0.61 |
| 0.070 ± 0.004 | 0.059 ± 0.013 | 0.055 ± 0.018 | 0.050 ± 0.007 | |||||||
| unfertilized | fertilized | |||||||||
| 0.063 ± 0.008 | 0.054 ± 0.008 | |||||||||
| Herbs (grasses+forbs) | very low | low | medium | high | 0.72 | 0.57 | 0.57 | 0.47 | 0.48 | 0.70 |
| 0.121 ± 0.005 | 0.109 ± 0.011 | 0.112 ± 0.014 | 0.098 ± 0.007 | |||||||
| unfertilized | fertilized | |||||||||
| 0.114 ± 0.008 | 0.106 ± 0.007 | |||||||||
| Shrubs | very low | low | medium | high | 2.32 | 0.15 | 0.15 | 0.71 | 1.32 | 0.33 |
| 0.073 ± 0.007 | 0.050 ± 0.007 | 0.061 ± 0.005 | 0.046 ± 0.012 | |||||||
| unfertilized | fertilized | |||||||||
| 0.059 ± 0.007 | 0.056 ± 0.006 | |||||||||
| Grand total | very low | low | medium | high | 0.51 | 0.69 | 6.88 | 0.03 | 0.04 | 0.99 |
| 61.00 ± 9.45 | 56.60 ± 5.96 | 69.14 ± 7.27 | 62.99 ± 9.46 | |||||||
| unfertilized | fertilized | |||||||||
| 52.85 ± 5.01b | 72.01 ± 3.42a | |||||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sullivan, T.P.; Sullivan, D.S.; Lindgren, P.M.F.; Ransome, D.B.; Zabek, L. Twenty-Five Years after Stand Thinning and Repeated Fertilization in Lodgepole Pine Forest: Implications for Tree Growth, Stand Structure, and Carbon Sequestration. Forests 2020, 11, 337. https://doi.org/10.3390/f11030337
Sullivan TP, Sullivan DS, Lindgren PMF, Ransome DB, Zabek L. Twenty-Five Years after Stand Thinning and Repeated Fertilization in Lodgepole Pine Forest: Implications for Tree Growth, Stand Structure, and Carbon Sequestration. Forests. 2020; 11(3):337. https://doi.org/10.3390/f11030337
Chicago/Turabian StyleSullivan, Thomas P., Druscilla S. Sullivan, Pontus M.F. Lindgren, Douglas B. Ransome, and Lisa Zabek. 2020. "Twenty-Five Years after Stand Thinning and Repeated Fertilization in Lodgepole Pine Forest: Implications for Tree Growth, Stand Structure, and Carbon Sequestration" Forests 11, no. 3: 337. https://doi.org/10.3390/f11030337
APA StyleSullivan, T. P., Sullivan, D. S., Lindgren, P. M. F., Ransome, D. B., & Zabek, L. (2020). Twenty-Five Years after Stand Thinning and Repeated Fertilization in Lodgepole Pine Forest: Implications for Tree Growth, Stand Structure, and Carbon Sequestration. Forests, 11(3), 337. https://doi.org/10.3390/f11030337