The Response of Erica arborea L. Tree Growth to Climate Variability at the Afro-alpine Tropical Highlands of North Ethiopia

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection and Tree-Ring Chronology
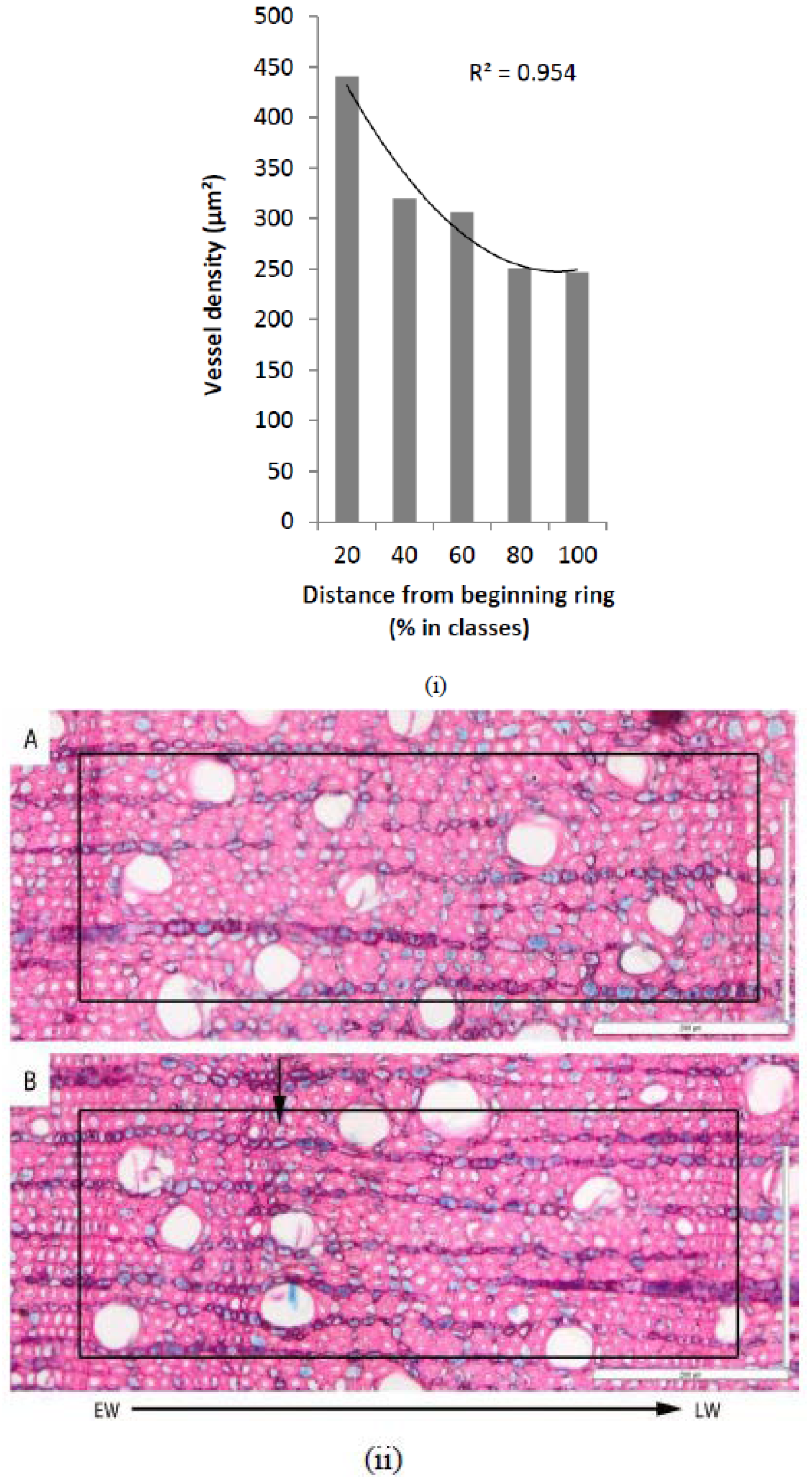
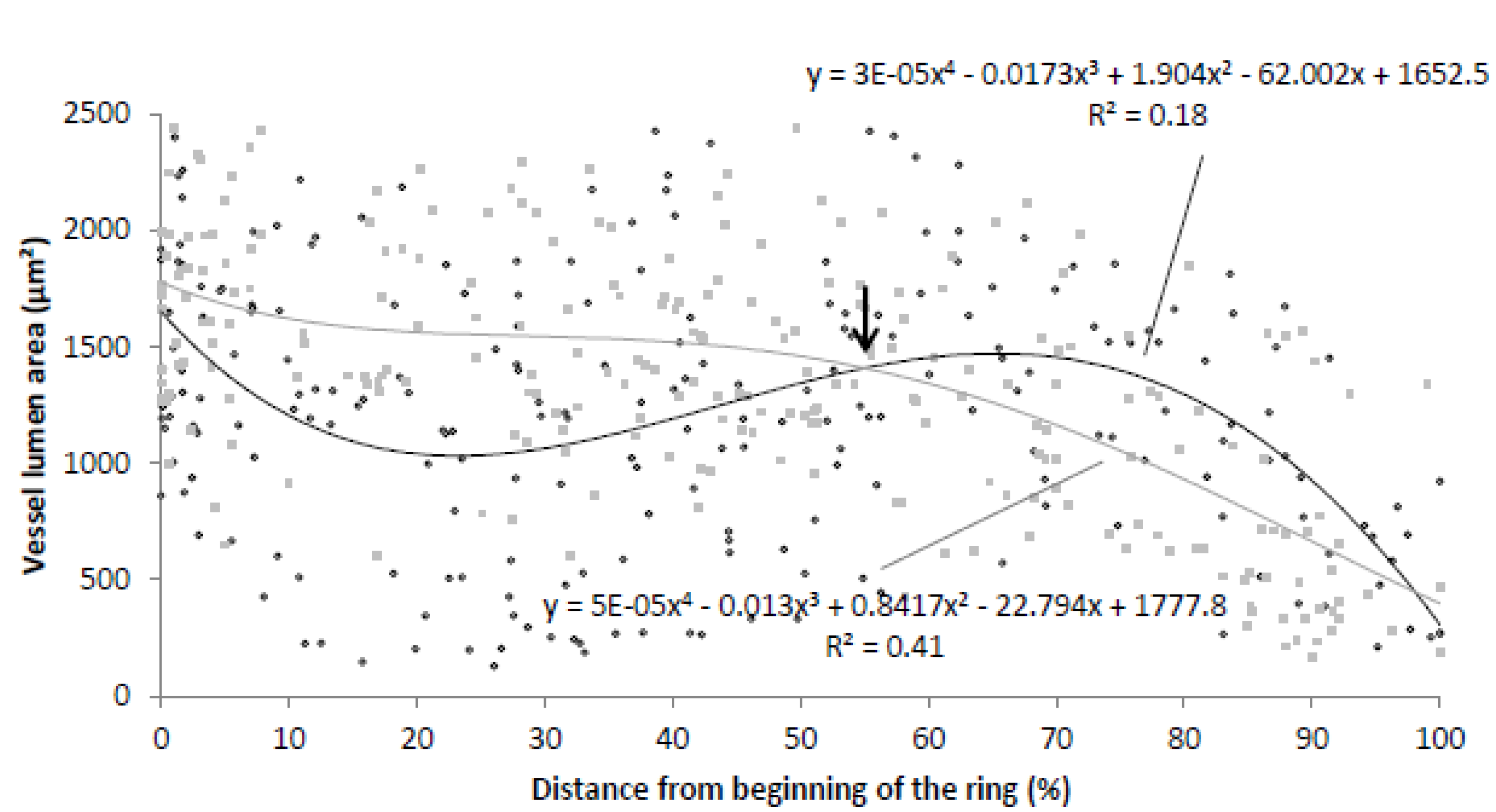
2.3. Wood Anatomical Analysis
2.4. Climate–Growth Relationship
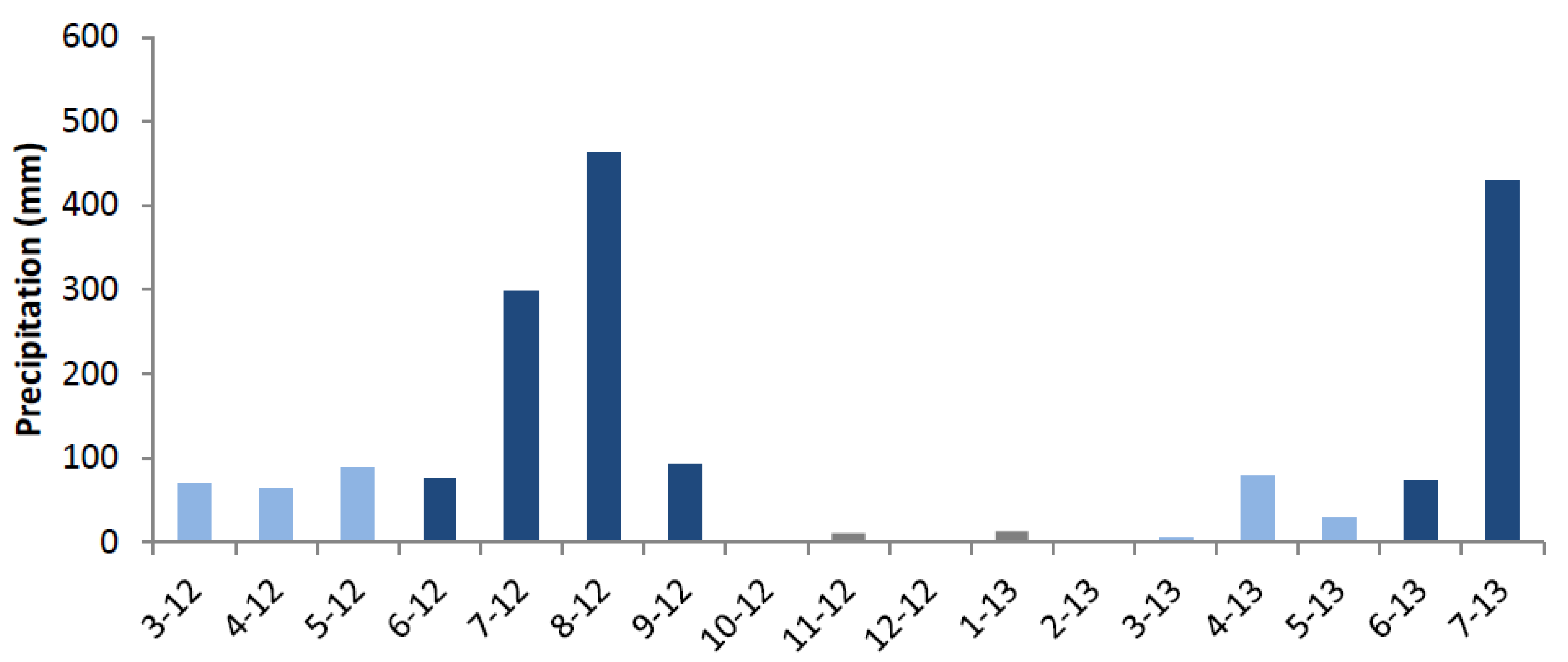
2.4.1. Relationship between the Occurrence of IADFs and Rainfall
- The variable “rainfall dip” is 1 if there is azmera (spring) rainfall with more than 100 mm followed by a dry spell of more than 10 days between the azmera (spring) and kiremt (summer) rain season, and 0 in all other cases.
- The variable “dry spell” is 1 if there is a dry spell of more than 20 days and 0 in all other cases.
2.4.2. Dendroclimatology
3. Results
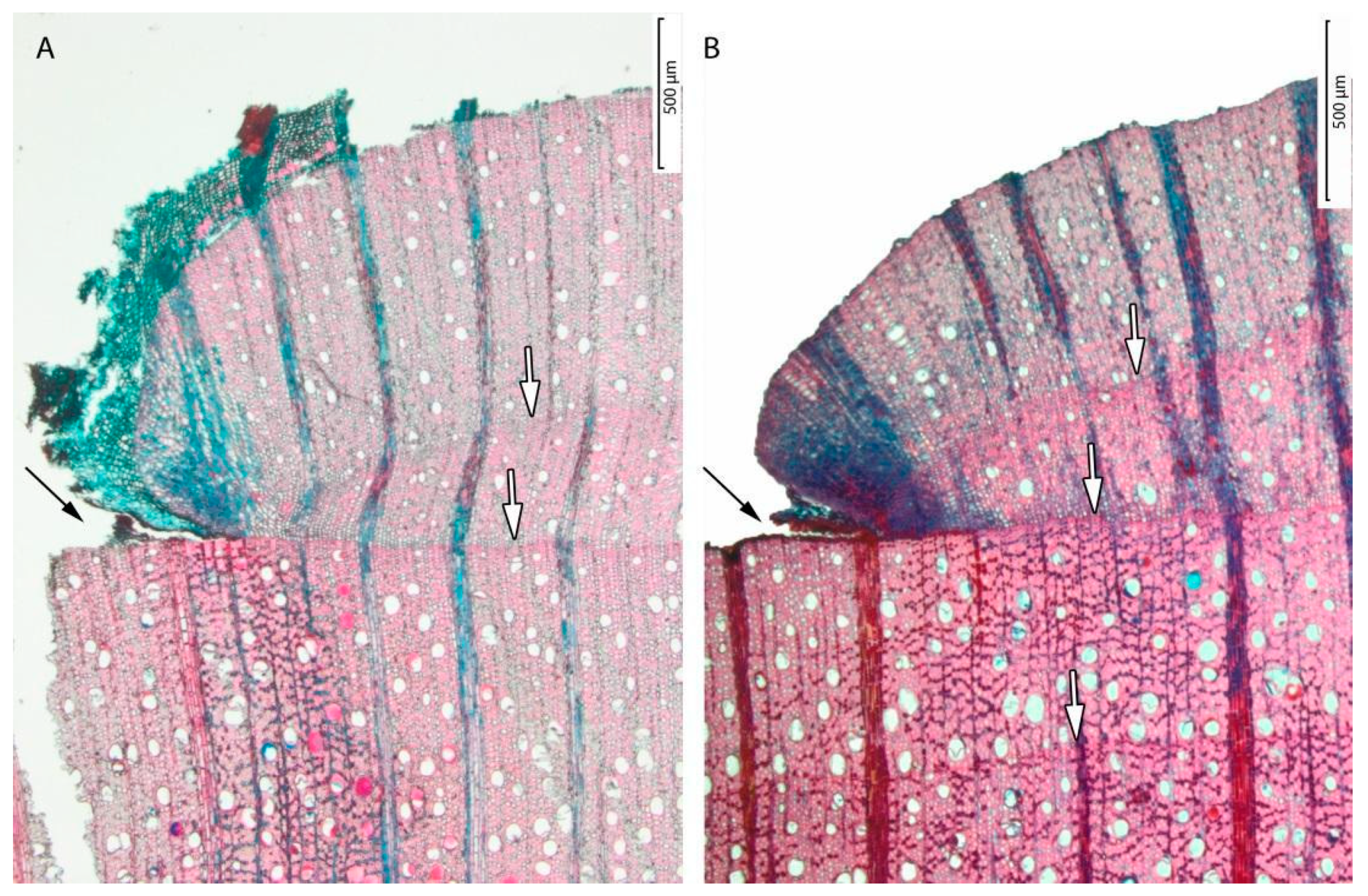
3.1. Tree-ring Formation
3.2. IADFs and Rainfall Variability
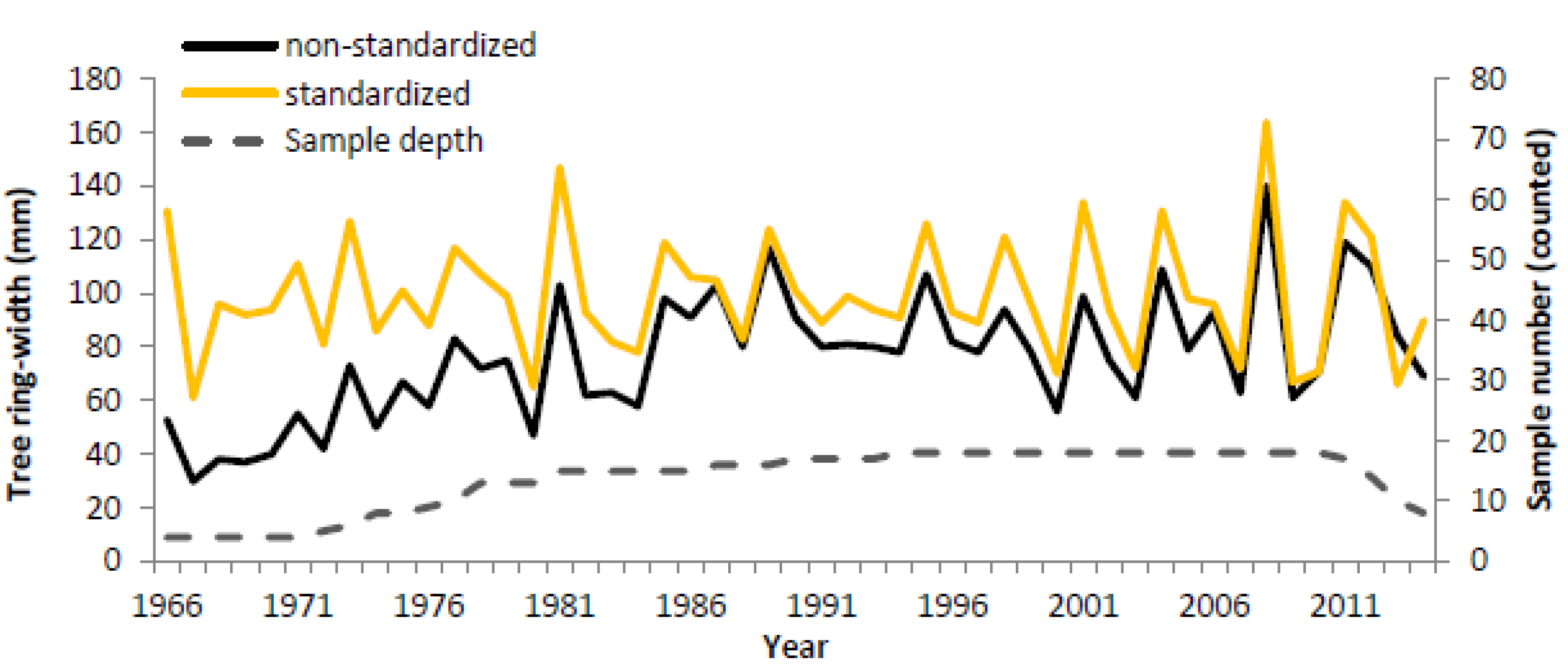
3.3. Dendrochronology
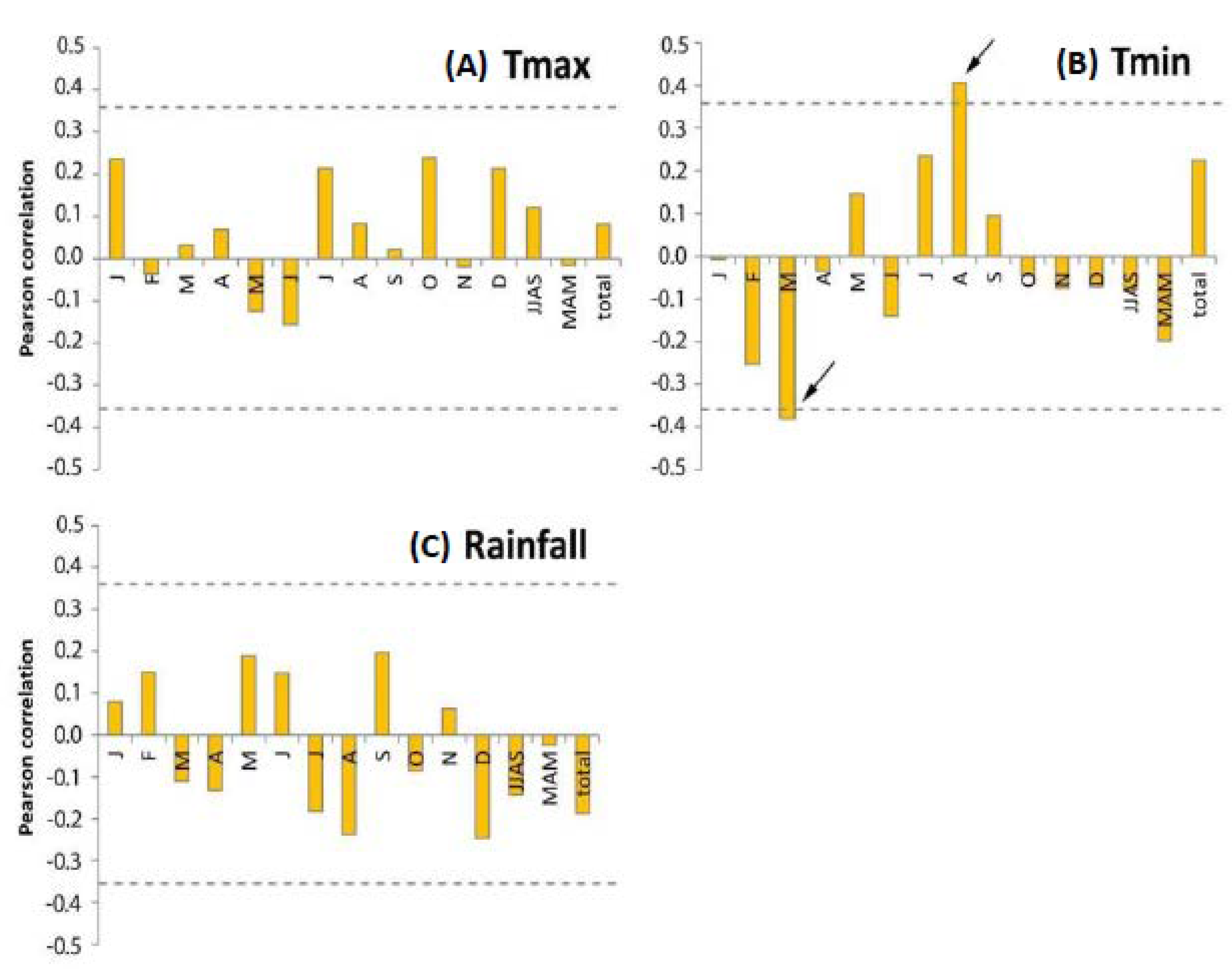
3.4. Dendroclimatology
4. Discussion
5. Conclusions and Outlook
Author Contributions
Funding
Conflicts of Interest
References
- Rossi, S.; Deslauriers, A. Intra-annual time scales in tree rings. Dendrochronologia 2007, 25, 75–77. [Google Scholar] [CrossRef]
- De Micco, V.; Battipaglia, G.; Cherubini, P.; Aronne, G. Comparing methods to analyse anatomical features of tree rings with and without intra-annual density fluctuations (IADFs). Dendrochronologia 2014, 32, 1–6. [Google Scholar] [CrossRef]
- Wils, T.; Sass-Klaassen, U.; Eshetu, Z.; Bräuning, A.; Gebrekirstos, A.; Couralet, C.; Robertson, I.; Touchan, R.; Koprowski, M.; Conway, D.; et al. Dendrochronology in the dry tropics: The Ethiopian case. Trees 2010, 25, 345–354. [Google Scholar] [CrossRef]
- Worbes, M. One hundred years of tree-ring research in the tropics: A brief history and an outlook to future challenges. Dendrochronologia 2002, 20, 217–231. [Google Scholar] [CrossRef]
- Trouet, V.; Esper, J.; Beeckman, H. Climate/growth relationships of Brachystegia spiciformis from the miombo woodland in south central Africa. Dendrochronologia 2010, 28, 161–171. [Google Scholar] [CrossRef]
- De Ridder, M.; Trouet, V.; Van den Bulcke, J.; Hubau, W.; van Acker, J.; Beeckman, H. A tree-ring based comparison of Terminalia superba climate-growth relationships in West and Central Africa. Trees-Struct. Funct. 2013, 27, 1225–1238. [Google Scholar] [CrossRef]
- Cherubini, P.; Gartner, B.L.; Tognetti, R.; Bräker, O.U.; Schoch, W.; Innes, J.L. Identification, measurement and interpretation of tree rings in woody species from mediterranean climates. Biol. Rev. Camb. Philos. Soc. 2003, 78, 119–148. [Google Scholar] [CrossRef] [PubMed]
- Worbes, M.; Staschel, R.; Roloff, A.; Junk, W. Tree ring analysis reveals age structure, dynamics and wood production of a natural forest stand in Cameroon. For. Ecol. Manag. 2003, 173, 105–123. [Google Scholar] [CrossRef]
- Schöngart, J.; Orthmann, B.; Hennenberg, K.; Porembski, S.; Worbes, M. Climate-growth relationships of tropical tree species in West Africa and their potential for climate reconstruction. Glob. Chang. Biol. 2006, 12, 1139–1150. [Google Scholar] [CrossRef]
- Couralet, C.; Sass-Klaassen, U.; Sterck, F.; Bekele, T.; Zuidema, P. Combining dendrochronology and matrix modelling in demographic studies: An evaluation for Juniperus procera in Ethiopia. For. Ecol. Manag. 2005, 216, 317–330. [Google Scholar] [CrossRef]
- Gebrekirstos, A.; Mitlöhner, R.; Teketay, D.; Worbes, M. Climate—Growth relationships of the dominant tree species from semi-arid savanna woodland in Ethiopia. Trees 2008, 22, 631–641. [Google Scholar] [CrossRef]
- Trouet, V.; Coppin, P.; Beeckman, H. Annual growth ring patterns in Brachystegia spiciformis reveal influence of precipitation on tree growth. Biotropica 2006, 38, 375–382. [Google Scholar] [CrossRef]
- Fichtler, E.; Trouet, V.; Beeckman, H.; Coppin, P.; Worbes, M. Climatic signals in tree rings of Burkea africana and Pterocarpus angolensis from semiarid forests in Namibia. Trees 2004, 18, 442–451. [Google Scholar] [CrossRef]
- Therrell, M.; Stahle, D.; Ries, L.; Shugart, H. Tree-ring reconstructed rainfall variability in Zimbabwe. Clim. Dyn. 2006, 26, 677–685. [Google Scholar] [CrossRef]
- Stahle, D. Useful strategies for the development of tropical tree-ring chronologies. IAWA J. 1999, 20, 249–253. [Google Scholar] [CrossRef]
- Wils, T.; Robertson, I.; Eshetu, Z.; Touchan, R.; Sass-Klaassen, U.; Koprowski, M. Crossdating Juniperus procera from North Gondar, Ethiopia. Trees 2010, 25, 71–82. [Google Scholar] [CrossRef]
- Miehe, G.; Miehe, S. Ericaceous Forests and Heathlands in the Bale Mountains of South Ethiopia: Ecology and Man’s Impact; Stiftung Walderhaltung in Afrika: Hamburg, Germany, 1994. [Google Scholar]
- Aerts, R.; November, E.; Behailu, M.; Deckers, J.; Hermy, M.; Muys, B. Forest rehabilitation: One approach to water conservation in central Tigray. Water Sci. Technol. 2002, 6, 34–37. [Google Scholar]
- Nyssen, J.; Poesen, J.; Moeyersons, J.; Deckers, J.; Haile, M.; Lang, A. Human impact on the environment in the Ethiopian and Eritrean highlands—A state of the art. Earth-Sci. Rev. 2004, 64, 273–320. [Google Scholar] [CrossRef]
- Conway, D.; Brooks, N.; Briffa, K.R.; Desta, S.; Merrin, P.D.; Jones, P.D. Exploring the Potential for Dendroclimatic Analysis in Northern Ethiopia; University of East Anglia, Climate Research Unit: Norwich, UK, 1997. [Google Scholar]
- Kaeppeli, M. Regeneration and Age structure of Relict Ericaceous Forests—A Dendrochronological Study Near the Timberline in the Simen Mountains, Ethiopia; University of Berne: Berne, Switzerland, 1998. [Google Scholar]
- Paulsen, J.; Weber, U.M.; Körner, C. Tree growth near treeline: Abrupt or gradual reduction with altitude? Arct. Antarct. Alp. Res. 2000, 32, 113–119. [Google Scholar] [CrossRef]
- Gea-Izquierdo, G.; Cherubini, P.; Battipaglia, G.; Gärtner, H. Xylem Adjustment in Erica Arborea to Temperature and Moisture Availability in Contrasting Climates. IAWA J. 2013, 34, 109–126. [Google Scholar] [CrossRef]
- Battipaglia, G.; DE Micco, V.; Brand, W.A.; Saurer, M.; Aronne, G.; Linke, P.; Cherubini, P. Drought impact on water use efficiency and intra-annual density fluctuations in Erica arborea on Elba (Italy). Plant. Cell Environ. 2014, 37, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Wesche, K.; Miehe, G.; Kaeppeli, M. The significance of fire for afroalpine ericaceous vegetation. Mt. Res. Dev. 2000, 20, 340–347. [Google Scholar] [CrossRef]
- Hurni, H.; Stähli, P. Simen Mountains, Ethiopia: Climate and Dynamics of Altitudinal Belts from the Last Cold Period to the Present Day; Geographisches Institut der Universität Bern: Bern, Switzerland, 1982; Volume 13. [Google Scholar]
- Mariaux, A. Les cernes dans les bois tropicaux africains nature et périodicité. Bois Forêts des Trop. 1967, 113, 3–14. [Google Scholar]
- Grissino Mayer, H. A manual and tutorial for the proper use of an increment borer. Tree-Ring Res. 2003, 59, 63–79. [Google Scholar]
- Gärtner, H.; Nievergelt, D. The core-microtome: A new tool for surface preparation on cores and time series analysis of varying cell parameters. Dendrochronologia 2010, 28, 85–92. [Google Scholar] [CrossRef]
- Gärtner, H.; Banzer, L.; Schneider, L.; Schweingruber, F.; Bast, A. Preparing micro sections of entire (dry) conifer increment cores for wood anatomical time-series analyses. Dendrochronologia 2015, 34, 19–23. [Google Scholar] [CrossRef]
- Rinn, F. TSAP-WinTM User Reference; RinnTech: Heidelberg, Germany, 2003. [Google Scholar]
- Douglass, A. Crossdating in Dendrochronology. J. For. 1941, 39, 825–831. [Google Scholar]
- De Ridder, M.; Toirambe, B.; van den Bulcke, J.; Bourland, N.; van Acker, J.; Beeckman, H. Dendrochronological potential in a semi-deciduous rainforest: The case of Pericopsis elata in Central Africa. Forests 2014, 5, 3087–3106. [Google Scholar] [CrossRef]
- Baillie, M.; Pilcher, J. A simple program for tree-ring research. Tree-Ring Bull. 1973, 33, 7–14. [Google Scholar]
- Wigley, T.; Briffa, K.; Jones, P. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J. Clim. Appl. Meteorol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Gärtner, H.; Cherubini, P.; Fonti, P.; von Arx, G.; Schneider, L.; Nievergelt, D.; Verstege, A.; Bast, A.; Schweingruber, F.; Büntgen, U. A technical perspective in modern tree-ring research-How to overcome dendroecological and wood anatomical challenges. J. Vis. Exp. 2015, 97, 52337. [Google Scholar] [CrossRef] [PubMed]
- Verheyden, A.; Kairo, J.G.; Beeckman, H.; Koedam, N. Growth rings, growth ring formation and age determination in the mangrove Rhizophora mucronata. Ann. Bot. 2004, 94, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Seleshi, Y.; Camberlin, P. Recent changes in dry spell and extreme rainfall events in Ethiopia. Theor. Appl. Climatol. 2005, 83, 181–191. [Google Scholar] [CrossRef]
- Campbell, I. Chi-squared and Fisher-Irwin tests of two-by-two tables with small sample recommendations. Stat. Med. 2007, 26, 3661–3675. [Google Scholar] [CrossRef]
- Trouet, V.; Van Oldenborgh, G. KNMI Climate Explorer: A web-based research tool for high-resolution paleoclimatology. Tree-Ring Res. 2013, 69, 3–13. [Google Scholar] [CrossRef]
- De Martonne, E. L’indice d’aridité. Bulletin de l’Associaction de Géographes Francais 1926, 9, 3–5. [Google Scholar] [CrossRef]
- Olivar, J.; Bogino, S.; Spiecker, H.; Bravo, F. Changes in climate-growth relationships and IADF formation over time of pine species (Pinus halepensis, P. pinaster and P. sylvestris) in Mediterranean environments. For. Syst. 2015, 24. [Google Scholar] [CrossRef]
- Olivar, J.; Rathgeber, C.; Bravo, F. Climate change, tree-ring width and wood density of pines in Mediterranean environments. IAWA 2015, 36, 267–269. [Google Scholar] [CrossRef]
- Bräuning, A.; Volland-Voigt, F.; Burchardt, I.; Ganzhi, O.; Nauss, T.; Peters, T. Climatic control of radial growth of Cedrela montana in a humid mountain rainforest in southern Ecuador. Erdkunde 2009, 63, 337–345. [Google Scholar] [CrossRef]
- Jacob, M. Treeline Dynamics and Forest Cover Change in Afroalpine Ethiopia, As Affected by Climate Change and Anthropo-Zoogenic Impacts. Ph.D. Thesis, Ghent University, Belgium, Germany, 2015. [Google Scholar]
- Morales, M.; Villalba, R.; Grau, R.; Paolini, L. Rainfall-controlled tree growth in high-elevation subtropical treelines. Ecology 2004, 85, 3080–3089. [Google Scholar] [CrossRef]
- Paulsen, J.; Körner, C. A climate-based model to predict potential treeline position around the globe. Alp. Bot. 2014, 124, 1–12. [Google Scholar] [CrossRef]
- Körner, C. Plant adaptation to cold climates. F1000Research 2016, 5, 2769. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Rainfall (mm) | Azmera * Rainfall (mm) | Dry Spell (<1 mm) in days | Rainfall Dip ** | Dry Spell ** | IADF ** | ||
|---|---|---|---|---|---|---|---|
| Azmera * | Kiremt * | ||||||
| 1997 | 1002.1 | 178.6 | 14 | 19 | 1 | 0 | 0 |
| 1998 | 789.8 | 81.4 | 17 | 23 | 0 | 1 | 1 |
| 1999 | 793.7 | 23 | 42 | 21 | 0 | 1 | 1 |
| 2000 | 693.9 | 60.6 | 28 | 9 | 0 | 0 | 0 |
| 2001 | 993.9 | 115.6 | 25 | 15 | 0 | 0 | 1 |
| 2002 | 722.1 | 87.1 | 29 | 14 | 0 | 0 | 0 |
| 2003 | 868.8 | 104.1 | 18 | 14 | 1 | 0 | 0 |
| 2004 | 567.3 | 35.3 | 38 | 15 | 0 | 0 | 0 |
| 2005 | 774.3 | 150.1 | 20 | 17 | 1 | 0 | 1 |
| 2006 | 851.3 | 97.2 | 17 | 13 | 0 | 0 | 0 |
| 2007 | 837.6 | 50.3 | 16 | 15 | 0 | 0 | 0 |
| 2008 | 704.4 | 79.8 | 38 | 14 | 0 | 0 | 0 |
| 2009 | 575.5 | 53.3 | 33 | 24 | 0 | 1 | 1 |
| 2010 | 771.7 | 78.9 | 17 | 15 | 0 | 0 | 0 |
| 2011 | 784.7 | 200.0 | 13 | 13 | 1 | 0 | 0 |
| 2013 | 1075.6 | 100.2 | 23 | 12 | 1 | 0 | 0 |
| Sample Type | Stem Disks | Cores | Combined |
|---|---|---|---|
| Total n tree samples | 10 | 20 | 30 |
| n samples in chronology | 6 (60%) | 12 (60%) | 18 (60%) |
| Mean number of tree rings | 46 ± 11 | 36 ± 9 | 39 ± 11 |
| Time span chronology | 1972–2012 (40 years) | 1976–2014 (38 years) | 1966–2014 (48 years) |
| Mean ring width chronology (mm) | 0.48 ± 0.15 | 1.03 ± 0.26 | 0.76 ± 0.24 |
| AC1 before standardization | −0.01 | −0.23 | 0.27 |
| MS2 after standardization | 35 | 33 | 29 |
| Pearson r3 | 0.20 | 0.07 | 0.08 |
| EPS4 | 0.60 | 0.46 | 0.63 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jacob, M.; De Ridder, M.; Vandenabeele, M.; Asfaha, T.; Nyssen, J.; Beeckman, H. The Response of Erica arborea L. Tree Growth to Climate Variability at the Afro-alpine Tropical Highlands of North Ethiopia. Forests 2020, 11, 310. https://doi.org/10.3390/f11030310
Jacob M, De Ridder M, Vandenabeele M, Asfaha T, Nyssen J, Beeckman H. The Response of Erica arborea L. Tree Growth to Climate Variability at the Afro-alpine Tropical Highlands of North Ethiopia. Forests. 2020; 11(3):310. https://doi.org/10.3390/f11030310
Chicago/Turabian StyleJacob, Miro, Maaike De Ridder, Marlies Vandenabeele, Tesfaalem Asfaha, Jan Nyssen, and Hans Beeckman. 2020. "The Response of Erica arborea L. Tree Growth to Climate Variability at the Afro-alpine Tropical Highlands of North Ethiopia" Forests 11, no. 3: 310. https://doi.org/10.3390/f11030310
APA StyleJacob, M., De Ridder, M., Vandenabeele, M., Asfaha, T., Nyssen, J., & Beeckman, H. (2020). The Response of Erica arborea L. Tree Growth to Climate Variability at the Afro-alpine Tropical Highlands of North Ethiopia. Forests, 11(3), 310. https://doi.org/10.3390/f11030310