Site Index Models for Main Forest-Forming Tree Species in Poland



Abstract
1. Introduction
2. Materials and Methods
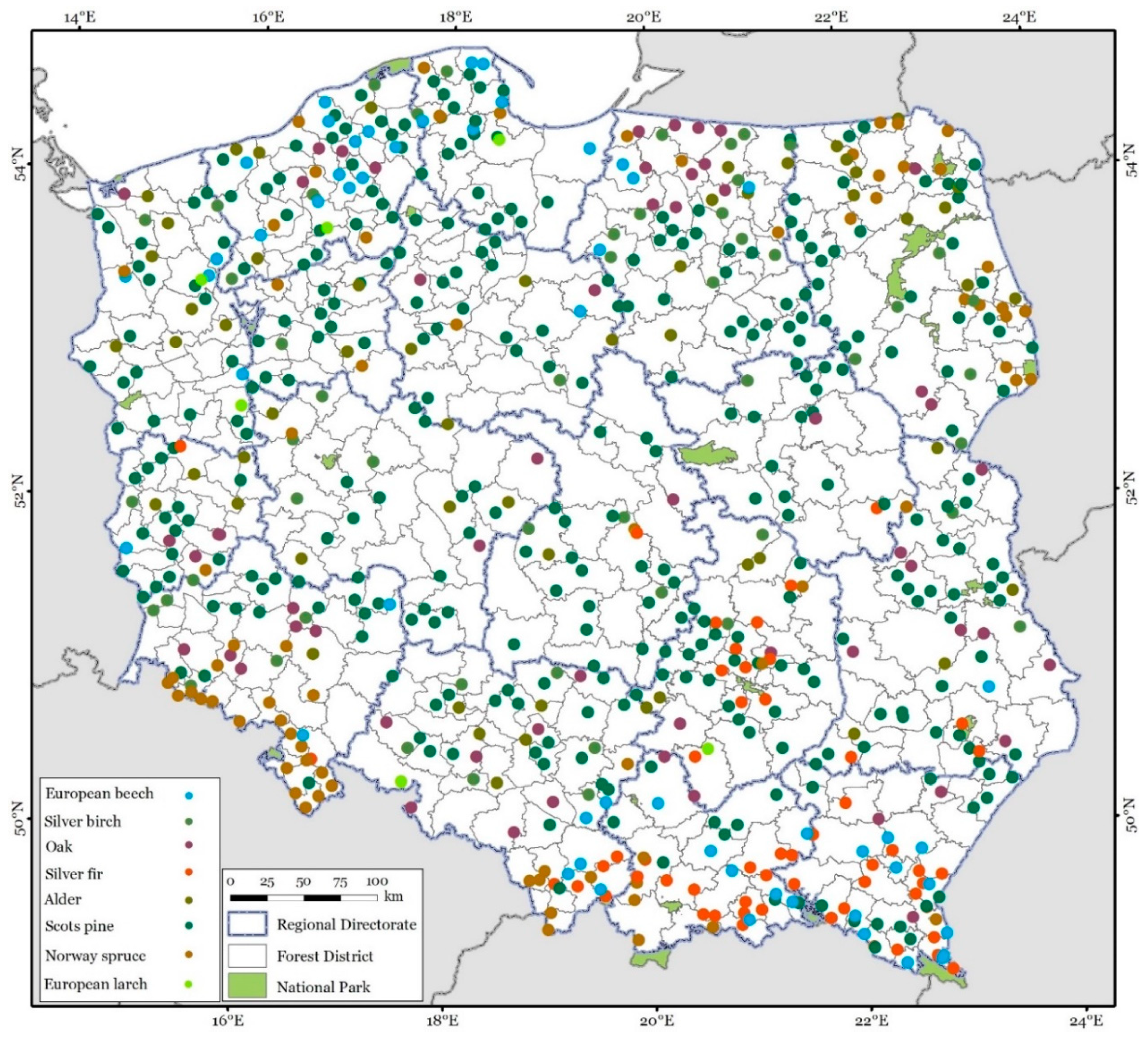
2.1. Collection of the Research Material
2.2. Stem Analysis
2.3. Models
- Polymorphism, allowing the model to take into account different site-specific growth patterns, depending on site conditions;
- Variable asymptotes for different sites;
- Equality of the site index and the height at base age;
- The possibility of using the same function as both a height growth model and a site index model.
2.4. Parameter Estimation
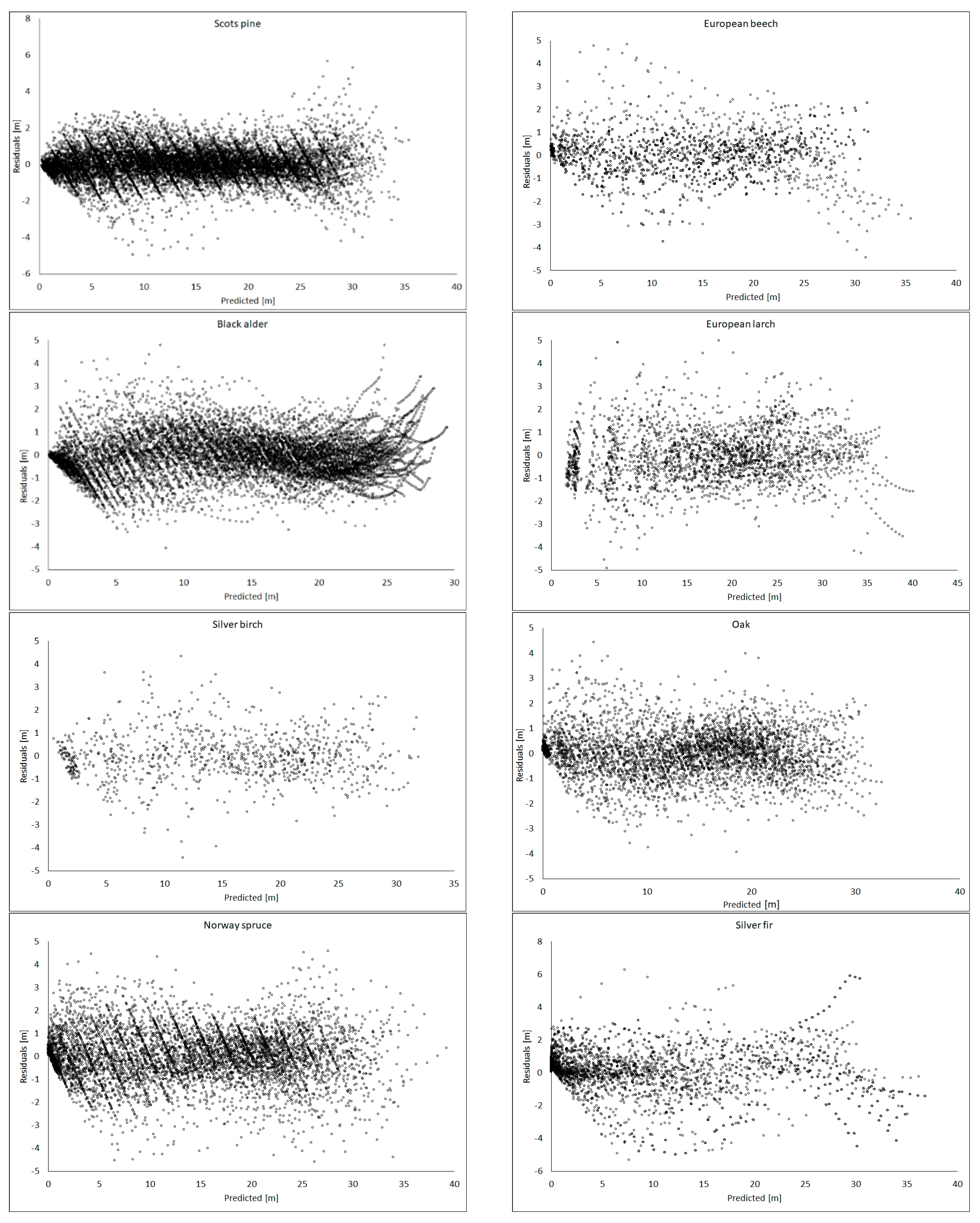
2.5. Model Evaluation
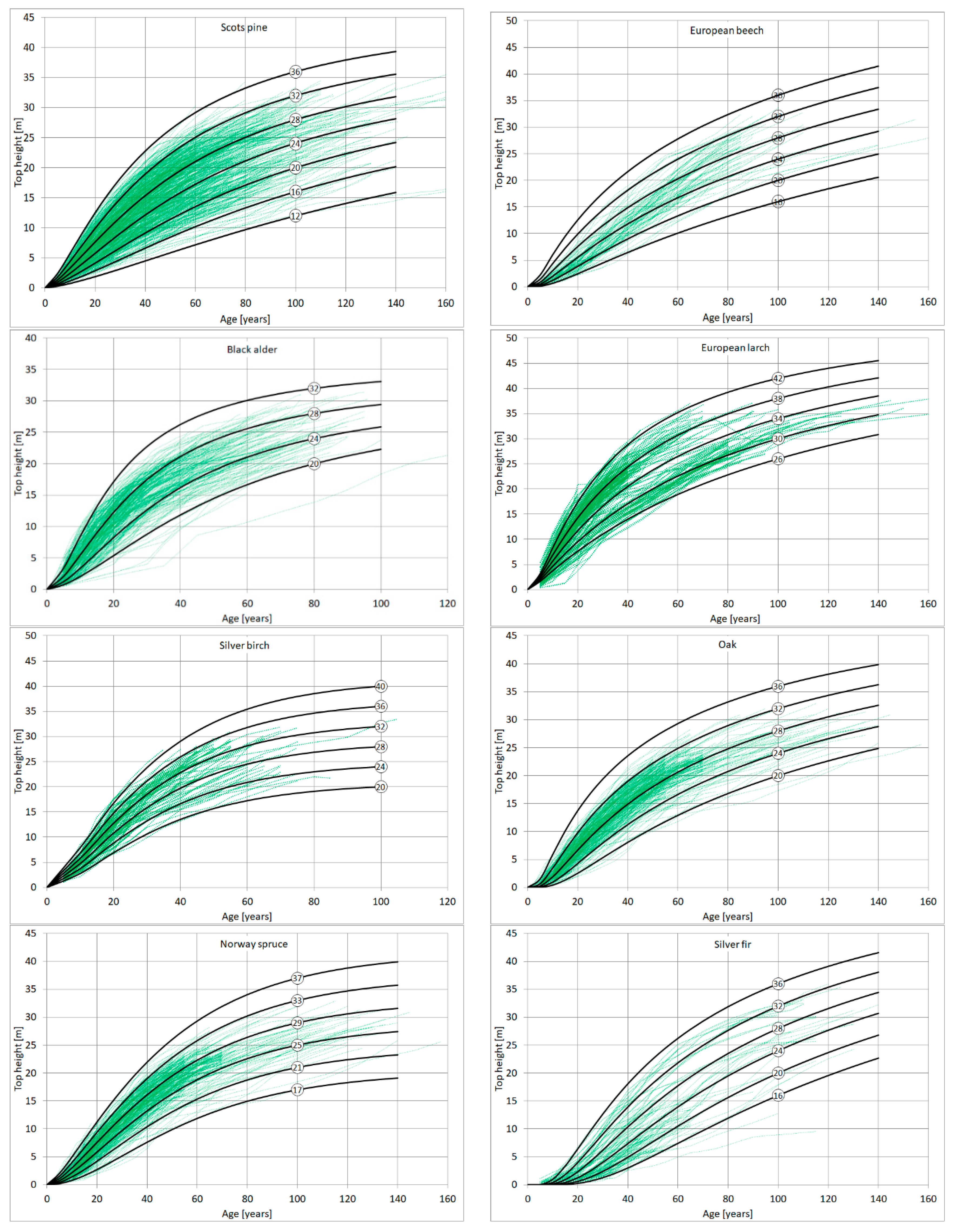
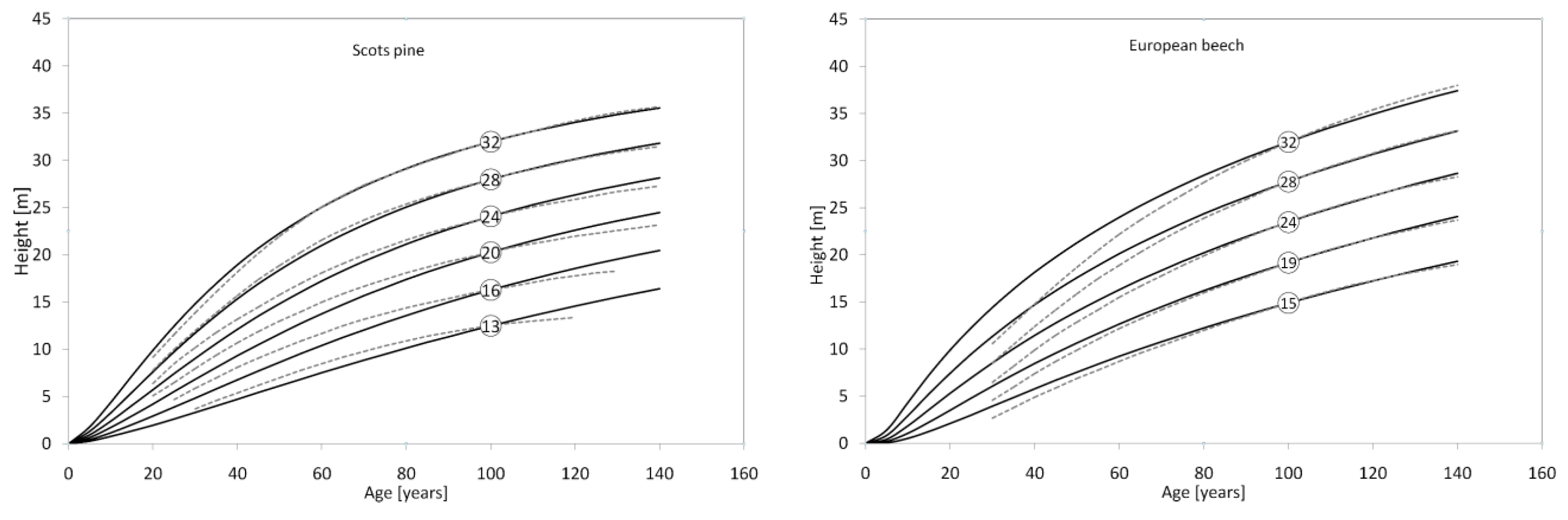
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Skovsgaard, J.P.; Vanclay, J.K. Forest site productivity: A review of the evolution of dendrometric concepts for even-aged stands. Forestry 2008, 81, 13–31. [Google Scholar] [CrossRef]
- Pretzsch, H.; Grote, R.; Reineking, B.; Rötzer, T.; Seifert, S. Models for forest ecosystem management: A European perspective. Ann. Bot. 2008, 101, 1065–1087. [Google Scholar] [CrossRef]
- Bontemps, J.J.D.; Bouriaud, O. Predictive approaches to forest site productivity: Recent trends, challenges and future perspectives. Forestry 2013, 87, 109–128. [Google Scholar] [CrossRef]
- Brandl, S.; Mette, T.; Falk, W.; Vallet, P.; Rötzer, T.; Pretzsch, H. Static site indices from different national forest inventories: Harmonization and prediction from site conditions. Ann. For. Sci. 2018, 75. [Google Scholar] [CrossRef]
- Splechtna, B.E. Height growth and site index models for Pacific silver fir in southwestern British Columbia. J. Ecosyst. Manag. 2001, 1, 1–14. [Google Scholar]
- Skovsgaard, J.P.; Vanclay, J.K. Forest site productivity: A review of spatial and temporal variability in natural site conditions. Forestry 2013, 86, 305–315. [Google Scholar] [CrossRef]
- Sharma, M.; Amateis, R.L.; Burkhart, H.E. Top height definition and its effect on site index determination in thinned and unthinned loblolly pine plantations. For. Ecol. Manage. 2002, 168, 163–175. [Google Scholar] [CrossRef]
- Hägglund, B.; Lundmark, J.E. Site index estimation by means of site properties of Scots pine and Norway spruce in Sweden. Stud. For. Suec. 1977, 138, 5–38. [Google Scholar]
- Johansson, T. Site Index Curves for Common Alder and Grey Alder Growing on Different Types of Forest Soil in Sweden. Scand. J. For. Res. 1999, 14, 441–453. [Google Scholar] [CrossRef]
- Raulier, F.; Lambert, M.C.; Pothier, D.; Ung, C.H. Impact of dominant tree dynamics on site index curves. For. Ecol. Manage. 2003, 184, 65–78. [Google Scholar] [CrossRef]
- Pretzsch, H. A Unified Law of Spatial Allometry for Woody and Herbaceous Plants. Plant Biol. 2002, 4, 159–166. [Google Scholar] [CrossRef]
- Pretzsch, H. Forest Dynamics, Growth, and Yield; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Bruchwald, A.; Zasada, M. Model wzrostu modrzewia europejskiego ( Larix decidua Mill) Growth model for European larch (Larix decidua Mill). Sylwan 2010, 158, 615–624. [Google Scholar]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Lexer, M.J.; Marchetti, M. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Boisvenue, C.; Running, S.W. Impacts of climate change on natural forest productivity—Evidence since the middle of the 20th century. Glob. Chang. Biol. 2006, 12, 862–882. [Google Scholar] [CrossRef]
- Swenson, J.J.; Waring, R.H.; Fan, W.; Coops, N. Predicting site index with a physiologically based growth model across Oregon, USA. Can. J. For. Res. 2005, 35, 1697–1707. [Google Scholar] [CrossRef]
- Coops, N.C.; Hember, R.A.; Waring, R.H. Assessing the impact of current and projected climates on Douglas-Fir productivity in British Columbia, Canada, using a process-based model (3-PG). Can. J. For. Res. 2010, 40, 511–524. [Google Scholar] [CrossRef]
- Mäkinen, H.; Yue, C.; Kohnle, U. Site index changes of Scots pine, Norway spruce and larch stands in southern and central Finland. Agric. For. Meteorol. 2017, 237–238, 95–104. [Google Scholar] [CrossRef]
- Yue, C.; Mäkinen, H.; Klädtke, J.; Kohnle, U. An approach to assessing site index changes of Norway spruce based on spatially and temporally disjunct measurement series. For. Ecol. Manage. 2014, 323, 10–19. [Google Scholar] [CrossRef]
- Yue, C.; Kahle, H.P.; von Wilpert, K.; Kohnle, U. A dynamic environment-sensitive site index model for the prediction of site productivity potential under climate change. Ecol. Modell. 2016, 337, 48–62. [Google Scholar] [CrossRef]
- Albert, M.; Schmidt, M. Climate-sensitive modelling of site-productivity relationships for Norway spruce (Picea abies (L.) Karst.) and common beech (Fagus sylvatica L.). For. Ecol. Manage. 2010, 259, 739–749. [Google Scholar] [CrossRef]
- Nabuurs, G.J.; Schelhaas, M.J.; Pussinen, A. Validation of the European Forest Information Scenario Model (EFISCEN) and a projection of Finnish forests. Silva Fenn. 2000, 34, 167–179. [Google Scholar] [CrossRef]
- Verkerk, P.J.; Fitzgerald, J.B.; Datta, P.; Dees, M.; Hengeveld, G.M.; Lindner, M.; Zudin, S. Spatial distribution of the potential forest biomass availability in Europe. For. Ecosyst. 2019, 6, 1–11. [Google Scholar] [CrossRef]
- Kurz, W.A.; Dymond, C.C.; White, T.M.; Stinson, G.; Shaw, C.H.; Rampley, G.J.; Smyth, C.; Simpson, B.N.; Neilson, E.T.; Trofymow, J.A.; et al. CBM-CFS3: A model of carbon-dynamics in forestry and land-use change implementing IPCC standards. Ecol. Modell. 2009, 220, 480–504. [Google Scholar] [CrossRef]
- Kim, M.; Lee, W.K.W.; Kurz, W.W.A.; Kwak, D.D.A.; Morken, S.; Smyth, C.E.C.; Ryu, D. Estimating carbon dynamics in forest carbon pools under IPCC standards in South Korea using CBM-CFS3. iForest Biogeosci. For. 2016, 9, 1–10. [Google Scholar] [CrossRef]
- Schwappach, A. Ertragstafeln der Wichtigeren Holzarten; Druckerei Merkur: Prag, Czech, 1943. [Google Scholar]
- Schober, R. Die Lärche – Eine ertragskundlich-biologische Untersuchung; Verlag M. u. H. Schaper: Hannover, Germany, 1949. [Google Scholar]
- Szymkiewicz, B. Niektóre zagadnienia dotyczące tablic zasobności drzewostanów sosnowych. Pr. IBL 1948, Seria A, 67. [Google Scholar]
- Cieszewski, C.J.; Strub, M.; Zasada, M. New dynamic site equation that fits best the Schwappach data for Scots pine (Pinus sylvestris L.) in Central Europe. For. Ecol. Manage. 2007, 243, 83–93. [Google Scholar] [CrossRef]
- Assman, E. Nauka o Produkcyjności Lasu; Państwowe Wydawnictwo Rolne i Lesne: Warszawa, Poland, 1968. [Google Scholar]
- Pretzsch, H.; Biber, P.; Schütze, G.; Uhl, E.; Rötzer, T. Forest stand growth dynamics in Central Europe have accelerated since 1870. Nat. Commun. 2014, 5, 4967. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G.; Biber, P. Drought can favour the growth of small in relation to tall trees in mature stands of Norway spruce and European beech. For. Ecosyst. 2018, 5, 20. [Google Scholar] [CrossRef]
- Sharma, R.P.; Brunner, A.; Eid, T. Site index prediction from site and climate variables for Norway spruce and Scots pine in Norway. Scand. J. For. Res. 2012, 27, 619–636. [Google Scholar] [CrossRef]
- Socha, J.; Ochał, W.; Grabczyński, S.; Maj, M. Modele Bonitacyjne dla Gatunków Lasotwórczych Polski Opracowane na Podstawie Tablic Zasobności*. Site Index Models for Forest−Forming Tree Species in Poland Developed Basing on the Yield Tables. Sylwan 2015, 159, 639–649. [Google Scholar]
- Bruchwald, A.; Michalak, K.; Wróblewski, L.; Zasada, M. Przestrzenne Zróżnicowanie Wzrostu Sosny. Wzrost Wysokości Sosny w Różnych Regionach Polski; Fundacja Rozwój SGGW: Warszawa, Poland, 2000. [Google Scholar]
- Bruchwald, A. Change in Top Height of Pine Forest Stands with Age; Pascal and Francis Bibliographic Databases: Warszawa, Poland, 1977. [Google Scholar]
- Bruchwald, A.; Dudek, A.; Michalak, K.; Zasada, M.; Rymer-Dudzinska, T.; Wroblewski, L. Model wzrostu dla drzewostanów świerkowych. Sylwan 1999, 143, 19–31. [Google Scholar]
- Bruchwald, A.; Dudzińska, M.; Wirowski, M. Model wzrostu dla olszy czarnej (Alnus glutinosa (L.) Gaertn.). Sylwan 2003, 147, 3–10. [Google Scholar]
- Bruchwald, A.; Dmyterko, E.; Wojtan, R. A growth model for European larch (Larix decidua Mill.) based on stand characteristics. For. Res. Pap. 2011, 72, 77–81. [Google Scholar] [CrossRef]
- Orzeł, S.; Forgiel, M.; Ochał, W.; Socha, J. Nadziemna biomasa i roczna produkcja drzewostanów sosnowych Puszczy Niepołomickiej * Aboveground biomass and annual production in stands. Sylwan 2006, 150, 1–17. [Google Scholar]
- Socha, J.; Orzeł, S. Dynamic site index curves for Scots pine stands in Niepołomice Primeval ForestDynamiczne krzywe bonitacyjne dla drzewostanów sosnowych Puszczy Niepołomickiej. Sylwan 2011, 155, 301–312. [Google Scholar]
- Cieszewski, C.J.; Zasada, M. Dynamiczna forma anamorficznego modelu bonitacyjnego dla sosny pospolitej w Polsce. Sylwan 2002, 146, 17–24. [Google Scholar]
- Goelz, J.C.G.; Burk, T.E. Measurement error causes bias in site index equations. Can. J. For. Res. 1996, 26, 1585–1593. [Google Scholar] [CrossRef]
- Strub, M.; Cieszewski, C.J. Base-age invariance properties of two techniques for estimating the parameters of site index models. For. Sci. 2006, 52, 182–186. [Google Scholar]
- Martín-Benito, D.; Gea-Izquierdo, G.; del Río, M.; Cañellas, I. Long-term trends in dominant-height growth of black pine using dynamic models. For. Ecol. Manage. 2008, 256, 1230–1238. [Google Scholar] [CrossRef]
- Bailey, R.L.; Clutter, J.L. Base-Age Invariant Polymorphic Site Curves. For. Sci. 1974, 20, 155–159. [Google Scholar]
- Cieszewski, J.; Bailey, L. Generalized Algebraic Difference Approach: Theory Based Derivation of Dynamic Site Equations with Polymorphism and Variable Asymptotes. For. Sci. 2000, 46, 116–126. [Google Scholar]
- Cieszewski, C.J. Three methods of deriving advanced dynamic site equations demonstrated on inland Douglas-fir site curves. Can. J. For. Res. For. 2001, 31, 165–173. [Google Scholar] [CrossRef]
- Kitikidou, K.; Kaymakis, M.; Milios, E. Site index curves for young Populus tremula stands on Athos Peninsula (northern Greece). Turk. J. Agric. For. 2012, 36, 55–63. [Google Scholar]
- Cieszewski, C.J. Developing a Well-Behaved Dynamic Site Equation Using a Modified Hossfeld IV Function Y 3 = (axm)/(c + x m–1), a Simplified Mixed-Model and Scant Subalpine Fir Data. For. Sci. 2003, 49, 539–554. [Google Scholar]
- Cieszewski, C.J.; Nigh, G.D. A dynamic equation for a published Sitka spruce site-dependent height-age model. For. Chron. 2002, 78, 690–694. [Google Scholar] [CrossRef][Green Version]
- Diéguez-Aranda, U.; Grandas-Arias, J.A.; Álvarez-González, J.G.; Von Gadow, K. Site quality curves for birch stands in North-Western Spain. Silva Fenn. 2006, 40, 631–644. [Google Scholar] [CrossRef]
- Diéguez-Aranda, U.; Burkhart, H.E.; Amateis, R.L. Dynamic site model for loblolly pine (Pinus taeda L.) plantations in the United States. For. Sci. 2006, 52, 262–272. [Google Scholar]
- Nord-Larsen, T. Developing Dynamic Site Index Curves for European Beech. For. Sci. 2006, 52, 173–181. [Google Scholar]
- Adame, P.; Hynynen, J.; Cañellas, I.; del Río, M. Individual-tree diameter growth model for rebollo oak (Quercus pyrenaica Willd.) coppices. For. Ecol. Manage. 2008, 255, 1011–1022. [Google Scholar] [CrossRef]
- Bravo-Oviedo, A.; del Río, M.; Montero, G. Geographic variation and parameter assessment in generalized algebraic difference site index modelling. For. Ecol. Manage. 2007, 247, 107–119. [Google Scholar] [CrossRef]
- Carmean, W.H. Site index curves for upland oaks in the central states. For. Sci. 1972, 18, 109–120. [Google Scholar]
- Kiviste, A.; Álvarez González, J.G.; Rojo-Alboreca, A.; Ruiz González, A.D. Funciones de Crecimiento de Aplicación en el Ámbito Forestal Faculty of Forestry; Ministerio de Ciencia y Tecnología: Madrid, Spain, 2002. [Google Scholar]
- Diéguez-Aranda, U.; Castedo Dorado, F.; Álvarez González, J.G.; Rojo Alboreca, A. Dynamic growth model for Scots pine (Pinus sylvestris L.) plantations in Galicia (north-western Spain). Ecol. Modell. 2006, 191, 225–242. [Google Scholar] [CrossRef]
- Sharma, R.P.; Brunner, A.; Eid, T.; Øyen, B.H. Modelling dominant height growth from national forest inventory individual tree data with short time series and large age errors. For. Ecol. Manage. 2011, 262, 2162–2175. [Google Scholar] [CrossRef]
- Ercanli, I.; Kahriman, A.; Yavuz, H. Dynamic base-age invariant site index models based on generalized algebraic difference approach for mixed Scots pine (Pinus sylvestris L.) and Oriental beech (Fagus orientalis Lipsky) stands. Turk. J. Agric. For. 2014, 38, 134–147. [Google Scholar] [CrossRef]
- Barrio Anta, M.; Dieguez-Aranda, U. Site quality of pedunculate oak (Quercus robur L.) stands in Galicia (northwest Spain). Eur. J. For. Res. 2005, 124, 19–28. [Google Scholar] [CrossRef]
- Elfving, B.; Kiviste, A. Construction of site index equations for Pinus sylvestris L. using permanent plot data in Sweden. For. Ecol. Manage. 1997, 98, 125–134. [Google Scholar] [CrossRef]
- Karlsson, K. Height growth patterns of Scots pine and Norway spruce in the coastal areas of western Finland. For. Ecol. Manag. 2000, 135, 205–216. [Google Scholar] [CrossRef]
- Bravo, F.; Montero, G. Site index estimation in Scots pine ( Pinus sylvestris L.) stands in the High Ebro Basin ( northern Spain) using soil attributes. Forestry 2001, 74, 395–406. [Google Scholar] [CrossRef]
- Palahı́, M.; Tomé, M.; Pukkala, T.; Trasobares, A.; Montero, G.; Palahí, M. Site index model for Pinus sylvestris in north-east Spain. For. Ecol. Manage. 2004, 187, 35–47. [Google Scholar] [CrossRef]
- Weiskittel, A.R.; Hann, D.W.; Kershaw, J.A., Jr.; Vanclay, J.K.; Kershaw, J.A.; Vanclay, J.K. Forest Growth and Yield Modeling; Wiley: Chichester, UK, 2011. [Google Scholar]
- Weiskittel, A.; Kuehne, C.; McTague, J.P.; Oppenheimer, M. Development and evaluation of an individual tree growth and yield model for the mixed species forest of the Adirondacks Region of New York, USA. For. Ecosyst. 2016, 3. [Google Scholar] [CrossRef]
- Socha, J.; Pierzchalski, M.; Bałazy, R.; Ciesielski, M. Modelling top height growth and site index using repeated laser scanning data. For. Ecol. Manage. 2017, 406, 307–3017. [Google Scholar] [CrossRef]
- Goelz, J.C.G.; Burk, T.E. Development of a well-behaved site index equation: Jack pine in north central Ontario. Can. J. Res. 1992, 22, 776–784. [Google Scholar] [CrossRef]
- Nord-Larsen, T.; Meilby, H.; Skovsgaard, J.P. Site-specific height growth models for six common tree species in Denmark. Scand. J. For. Res. 2009, 24, 194–204. [Google Scholar] [CrossRef]
- Socha, J. Site index curves for Norway spruce on mountain habitatsKrzywe bonitacyjne świerka pospolitego na siedliskach górskich. Sylwan 2011, 155, 816–826. [Google Scholar]
- Krumland, B.; Eng, H. Site Index Systems For Major Young-Growth Forest and Woodland Species in North California. Calif. For. 2005, 4, 1–220. [Google Scholar]
- Cieszewski, C.J. GADA derivation of dynamic site equations with polymorphism and variable asymptotes from Richards to Weibull and other exponential functions. For. Sci. 2004, 54, 303–315. [Google Scholar]
- Seki, M.; Sakici, O.E. Dominant height growth and dynamic site index models for Crimean pine in the Kastamonu-Taşköprü region of Turkey. Can. J. For. Res. 2017, 47, 1441–1449. [Google Scholar] [CrossRef]
- Anta, M.B.; Dorado, F.C.; Diéguez-Aranda, U.; Álvarez González, J.G.; Parresol, B.R.; Soalleiro, R.R. Development of a basal area growth system for maritime pine in northwestern Spain using the generalized algebraic difference approach. Can. J. For. Res. 2006, 36, 1461–1474. [Google Scholar] [CrossRef]
- Cieszewski, C.J.; Harrison, M.; Martin, S.W. Examples of Practical Methods for Unbiased Parameter Estimation in Self-Referencing Functions; University of Georgia: Athens, GA, USA, 2000. [Google Scholar]
- Parresol, B.R.; Vissage, J.S. White Pine Site Index for the Southern Forest Survey White Pine Site Index for the Southern Forest Survey; U.S. Department of Agriculture, Forest Service, Southern Research: Asheville, NC, USA, 1998.
- Cieszewski, C.J.; Zasada, M. Model bonitacyjny dla sosny na podstawie tablic zasobności Szymkiewicza. Sylwan 2003, 147, 51–62. [Google Scholar]
- Szymkiewicz, B. Tablice Zasobności i Przyrostu Drzewostanów; Państwowe Wydawnictwo Rolnicze i Leśne: Warszawa, Poland, 2001. [Google Scholar]
- Magin, V.R. Ertragskundliche Unteruchungen in Montanen Mischwaldern. Forstwiss. Cbl. 1954, 73, 103–113. [Google Scholar] [CrossRef]
- Huang, S. Development of compatible height and site in- dex models for young and mature stands within an ecosys- tem based management framework. In Empirical and Process-Based Models for Forest Tree and Stand Growth Simulation; Technical University of Lisboa: Oeiras, Portugal, 1999; pp. 61–98. [Google Scholar]
- Perin, J.; Hébert, J.; Brostaux, Y.; Lejeune, P.; Claessens, H. Modelling the top-height growth and site index of Norway spruce in Southern Belgium. For. Ecol. Manage. 2013, 298, 62–70. [Google Scholar] [CrossRef]
- Nieróbca, A.; Wróblewska, E.; Mizak, K.; Kozyra, J. Zmiana długości okresu wegetacyjnego w Polsce. Woda Środowisko Obsz. Wiej. 2013, 13, 81–94. [Google Scholar]
- Jabłoński, M.; Budniak, P. Estimating aboveground woody biomass of forests in Poland for FAO/ECE and UNFCCC reporting. For. Res. Pap. 2014, 75, 277–289. [Google Scholar] [CrossRef][Green Version]
- Bøhler, F.; Bernt-Håvar, Ø. Estimation of site-index in old, semi-natural stands of Norway spruce at high altitude. Book of abstracts for the conference. Rapport fra Skog og landskap. In Proceedings of the Forest Management and Silviculture in the North—Balancing Future Needs, Stjørdal, Norway, 6–8 September 2011; p. 53. [Google Scholar]
- Coops, N.C.; Hember, R.A. Physiologically derived predictions of Douglas-fir site index in British Columbia. For. Chron. 2009, 85, 733–744. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
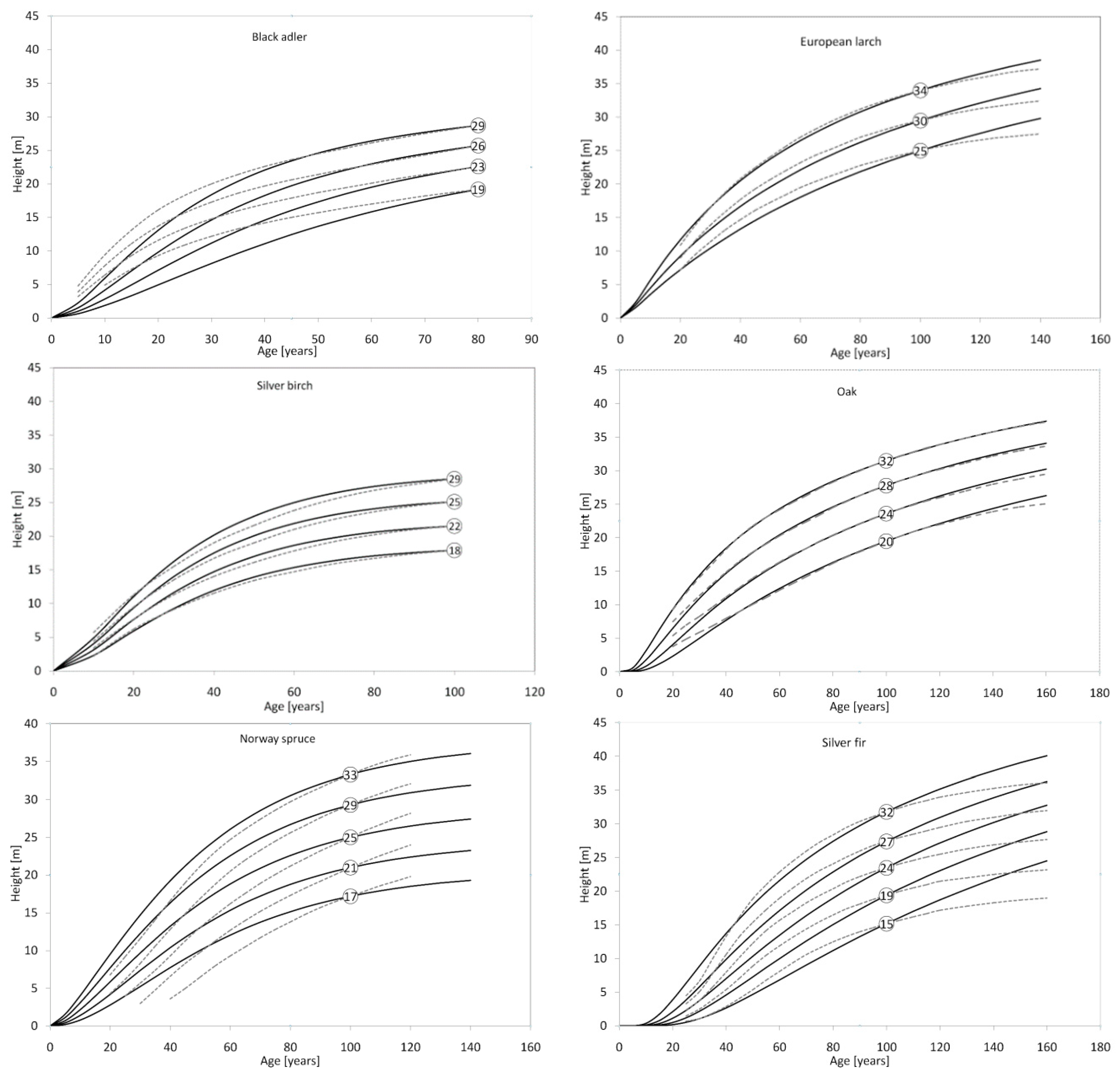{kind=link}
| Species | Number of Growth Series | Age (Years) | Height (m) | ||
|---|---|---|---|---|---|
| Min | Max | Min | Max | ||
| Scots pine (Pinus sylvestris) | 988 | 12 | 175 | 7.07 | 36.75 |
| European beech (Fagus sylvatica) | 327 | 60 | 180 | 15.79 | 33.56 |
| Black alder (Alnus glutinosa) | 448 | 7 | 120 | 5.15 | 31.4 |
| European larch (Larix decidua) | 227 | 20 | 170 | 16.38 | 38.55 |
| Silver birch (Betula pendula) | 99 | 20 | 105 | 10.46 | 33.46 |
| Oak (Quercus sp.) | 317 | 18 | 145 | 9.5 | 32.91 |
| Norway spruce (Picea abies) | 419 | 10 | 154 | 1.27 | 41.57 |
| Silver fir (Abies alba) | 227 | 20 | 215 | 1.7 | 36.16 |
| 3052 | |||||
| Model No. | Base Model Forms | Parameter Related to Site and Solution for theoretical Variable X | Dynamic GADA Formulation and Reference |
|---|---|---|---|
| M1 | Cieszewski [48] | ||
| M2 | Krumland and Eng [73] | ||
| M3 | with | Cieszewski [74] | |
| M4 | Sharma et al. [60] | ||
| M5 | Anta et al. [76] |
| Species | Function | Parameters | Estimate | R2adj | AIC | MAE |
|---|---|---|---|---|---|---|
| Scots pine | M1 | b1 | 1.366 | 0.989 | 35324 | 0.6 |
| b2 | 5802.862 | |||||
| b3 | 30.581 | |||||
| M2 | b2 | 0.017 | 0.986 | 39205 | 0.65 | |
| b3 | 1.236 | |||||
| M3 | b1 | 0.020 | 0.988 | 36937 | 0.61 | |
| b2 | 0.860 | |||||
| b3 | 8.425 | |||||
| M4 | b1 | 10.848 | 0.989 | 35608 | 0.61 | |
| b2 | 0.019 | |||||
| M5 | b0 | 101.255 | 0.988 | 36518 | 0.64 | |
| b2 | 0.436 | |||||
| European beech | M1 | b1 | 1.366 | 0.987 | 8604 | 0.677 |
| b2 | 5802.952 | |||||
| b3 | 30.581 | |||||
| M2 | b2 | 0.015 | 0.987 | 8730 | 0.684 | |
| b3 | 1.675 | |||||
| M3 | b1 | 0.017 | 0.988 | 8561 | 0.672 | |
| b2 | 1.243 | |||||
| b3 | 7.797 | |||||
| M4 | b1 | 47.661 | 0.986 | 8895 | 0.707 | |
| b2 | 0.013 | |||||
| M5 | b0 | 206.947 | 0.987 | 8797 | 0.697 | |
| b2 | 0.415 | |||||
| Black alder | M1 | b1 | 1.565 | 0.985 | 28359 | 0.728 |
| b2 | 1644.059 | |||||
| b3 | 29.161 | |||||
| M2 | b2 | 0.035 | 0.981 | 30673 | 0.83 | |
| b3 | 1.472 | |||||
| M3 | b1 | 0.039 | 0.983 | 29327 | 0.77 | |
| b2 | 0.159 | |||||
| b3 | 31.764 | |||||
| M4 | b1 | 9.244 | 0.984 | 28803 | 0.74 | |
| b2 | 0.025 | |||||
| M5 | b0 | 101.255 | 0.983 | 36518 | 0.645 | |
| b2 | 0.436 | |||||
| European larch | M1 | b1 | 1.328 | 0.984 | 8783 | 0.843 |
| b2 | 625.507 | |||||
| b3 | 43.486 | |||||
| M2 | b2 | 0.017 | 0.978 | 9661 | 1.044 | |
| b3 | 1.084 | |||||
| M3 | b1 | 0.023 | 0.982 | 9075 | 0.897 | |
| b2 | 0.346 | |||||
| b3 | 21.046 | |||||
| M4 | b1 | 4.639 | 0.983 | 8948 | 0.878 | |
| b2 | 0.018 | |||||
| M5 | b0 | 78.357 | 0.983 | 8879 | 0.848 | |
| b2 | 0.514 | |||||
| Silver birch | M1 | b1 | 1.366 | 0.984 | 3007 | 0.744 |
| b2 | 5802.952 | |||||
| b3 | 30.581 | |||||
| M2 | b2 | 0.036 | 0.983 | 3079 | 0.777 | |
| b3 | 1.580 | |||||
| M3 | b1 | 0.038 | 0.983 | 3042 | 0.768 | |
| b2 | 1.120 | |||||
| b3 | 12.810 | |||||
| M4 | b1 | 11.337 | 0.984 | 2999 | 0.731 | |
| b2 | 0.023 | |||||
| M5 | b0 | 101,255 | 0.984 | 3172 | 0.845 | |
| b2 | 0,436 | |||||
| Oak | M1 | b1 | 1.664 | 0.987 | 12376 | 0.691 |
| b2 | 7635.281 | |||||
| b3 | 30.359 | |||||
| M2 | b2 | 0.022 | 0.984 | 13253 | 0.776 | |
| b3 | 1.591 | |||||
| M3 | b1 | 0.025 | 0.986 | 12632 | 0.708 | |
| b2 | 0.699 | |||||
| b3 | 18.602 | |||||
| M4 | b1 | 27.831 | 0.986 | 12632 | 0.708 | |
| b2 | 0.022 | |||||
| M5 | b0 | 60.331 | 0.986 | 12511 | 0.697 | |
| b2 | 0.654 | |||||
| Norway spruce | M1 | b1 | 1.833 | 0.986 | 19814 | 0.775 |
| b2 | 6121.850 | |||||
| b3 | 40.956 | |||||
| M2 | b2 | 0.016 | 0.977 | 23053 | 1.009 | |
| b3 | 1.606 | |||||
| M3 | b1 | 0.025 | 0.985 | 20405 | 0.828 | |
| b2 | 0.770 | |||||
| b3 | 24.012 | |||||
| M4 | b1 | 31.549 | 0.979 | 22498 | 0.96 | |
| b2 | 0.014 | |||||
| M5 | b0 | 86.625 | 0.986 | 20024 | 0.8 | |
| b2 | 0.642 | |||||
| Silver fir | M1 | b1 | 1.833 | 0,976 | 12075 | 0.992 |
| b2 | 6121.850 | |||||
| b3 | 40.956 | |||||
| M2 | b2 | 0.007 | 0.959 | 13629 | 1.285 | |
| b3 | 1.478 | |||||
| M3 | b1 | 0.017 | 0.975 | 12018 | 0.993 | |
| b2 | 1.439 | |||||
| b3 | 6.830 | |||||
| M4 | b1 | 66.678 | 0.959 | 13614 | 1.285 | |
| b2 | 0.011 | |||||
| M5 | b0 | 63.533 | 0.978 | 11596 | 0.916 | |
| b2 | 0.867 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Socha, J.; Tymińska-Czabańska, L.; Grabska, E.; Orzeł, S. Site Index Models for Main Forest-Forming Tree Species in Poland. Forests 2020, 11, 301. https://doi.org/10.3390/f11030301
Socha J, Tymińska-Czabańska L, Grabska E, Orzeł S. Site Index Models for Main Forest-Forming Tree Species in Poland. Forests. 2020; 11(3):301. https://doi.org/10.3390/f11030301
Chicago/Turabian StyleSocha, Jarosław, Luiza Tymińska-Czabańska, Ewa Grabska, and Stanisław Orzeł. 2020. "Site Index Models for Main Forest-Forming Tree Species in Poland" Forests 11, no. 3: 301. https://doi.org/10.3390/f11030301
APA StyleSocha, J., Tymińska-Czabańska, L., Grabska, E., & Orzeł, S. (2020). Site Index Models for Main Forest-Forming Tree Species in Poland. Forests, 11(3), 301. https://doi.org/10.3390/f11030301