Genetic Improvement of Pinus koraiensis in China: Current Situation and Future Prospects



Abstract
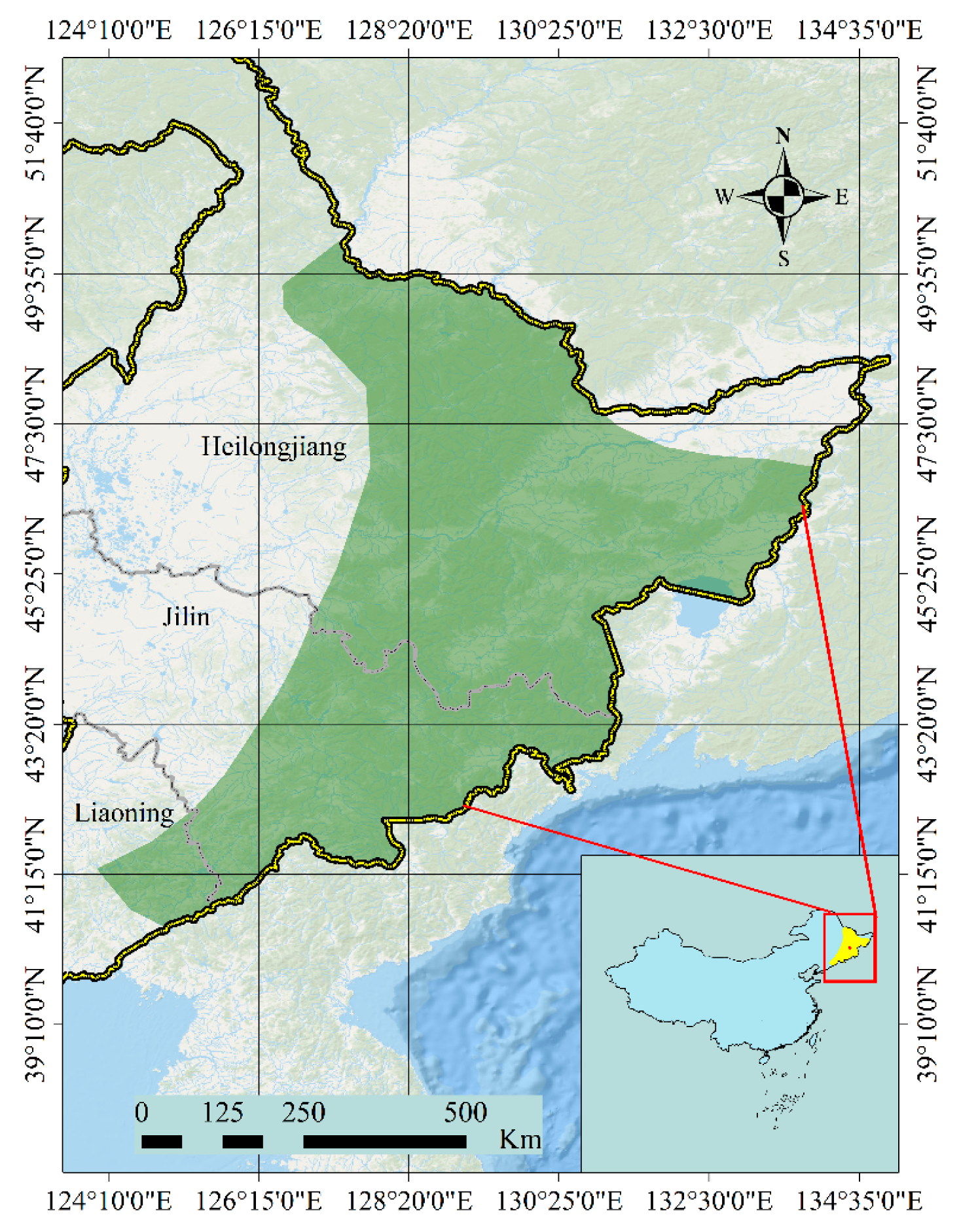
1. Introduction
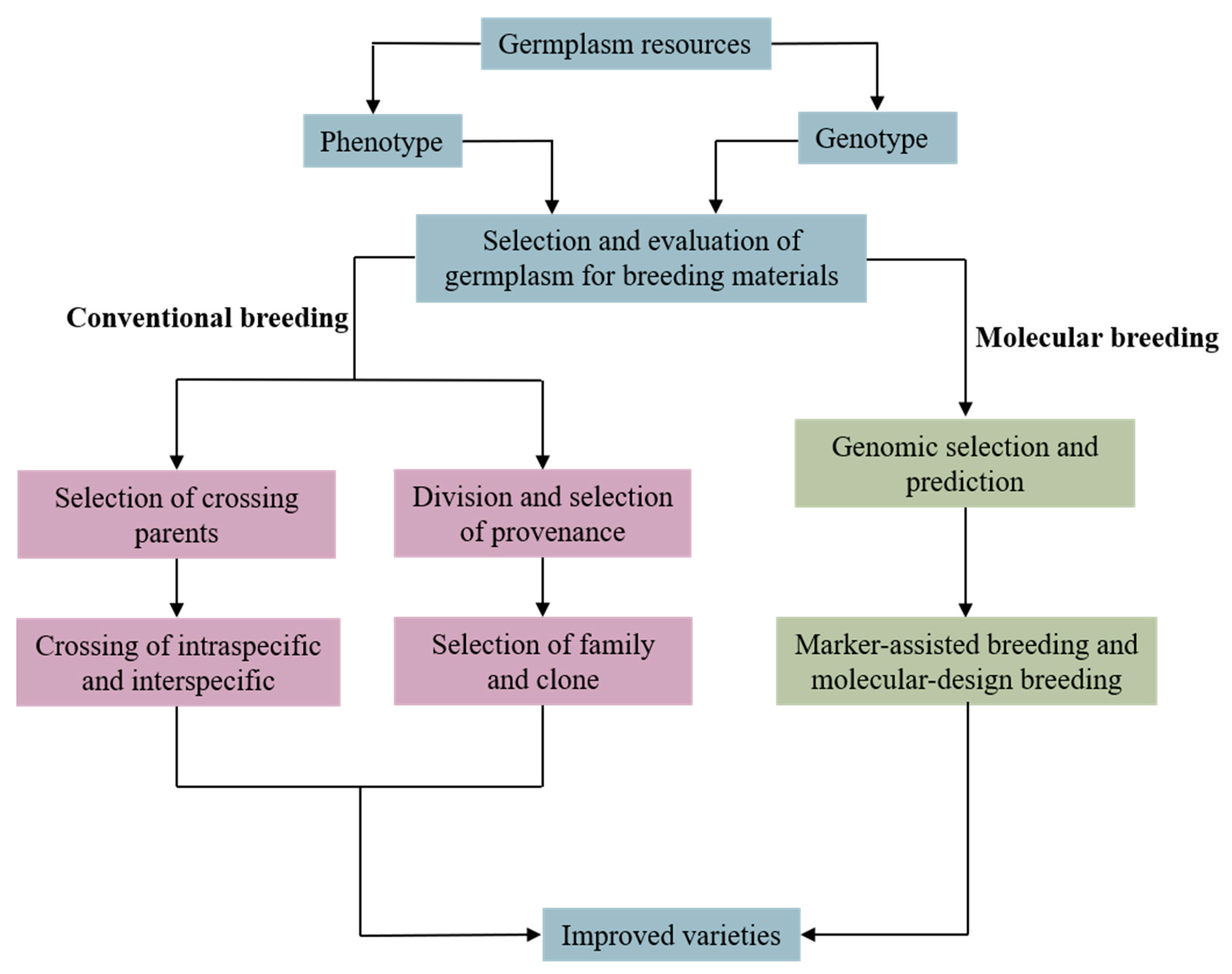
2. Research Progress on Conventional Breeding of Pinus koraiensis
3. Research Progress on Molecular Breeding of Pinus koraiensis
3.1. Genetic Structure and Genetic Diversity
3.2. Transcriptomics
3.3. Construction of Genetic Map
4. Other Related Research Works on Pinus koraiensis
5. Research Gaps and Future Research Directions
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Barnes, B.V.; Xü, Z.; Zhao, S. Forest ecosystems in an old-growth pine-mixed hardwood forest of the Changbai Mountain preserve in northeastern China. Can. J. For. Res. 1992, 22, 144–160. [Google Scholar] [CrossRef]
- Park, J.M.; Kwon, S.H.; Lee, H.J.; Na, S.J.; El-Kassaby, Y.A.; Kang, K.S. Integrating fecundity variation and genetic relatedness in estimating the gene diversity of seed crops: Pinus koraiensis seed orchard as an example. Can. J. For. Res. 2016, 47, 366–370. [Google Scholar] [CrossRef]
- Wang, L.; Li, X.; Wang, H. Physicochemical properties, bioaccessibility and antioxidant activity of the polyphenols from pine cones of Pinus koraiensis. Int. J. Biol. Macromol. 2018, 126, 385–391. [Google Scholar] [CrossRef]
- Shpatov, A.V.; Popov, S.A.; Salnikova, O.I.; Kukina, T.P.; Shmidt, E.N.; Um, B.H. Composition and bioactivity of lipophilic metabolites from needles and twigs of Korean and Siberian Pinus. Chem. Biodivers. 2017, 14, e1600203. [Google Scholar] [CrossRef]
- Lai, F.; Xie, H.; Wang, J. Functions of biology and disease control with pycnogonid from pine bark extract. Chin. Tradit. Herb. Drugs 2005, 36, 127–131. [Google Scholar]
- Liu, R.; Wang, Z.; Cui, J.; Deng, X.; Lu, J. Elicitations on the synthesis polyphenols of Pinus koraiensis. J. Beijing Univ. 2013, 35, 26–31. [Google Scholar]
- Fan, X.; Ruan, C.; Zhang, W.; Ding, J. Dynamic changes of oil content in Pinus koraiensis seed during development and fatty acid composition of its oil. China Oils Fats 2019, 44, 118–120. [Google Scholar]
- Ma, J.; Zhuang, L.; Chen, D.; Li, J. Geographical distribution of Korean pine. J. Northeast For. Univ. 1992, 20, 40–48. [Google Scholar]
- Wang, F.; Liang, D.; Pei, X.; Zhang, Q.; Zhang, P.; Zhang, J.; Lu, Z.; Yang, Y.; Liu, G.; Zhao, X. Study on the physiological indices of Pinus sibirica and Pinus koraiensis seedlings under cold stress. J. For. Res. 2018, 30, 1255–1265. [Google Scholar] [CrossRef]
- Zhang, Y.; Yi, X.; Wang, Y.; Ji, L.; Wu, P. Impact of tree harvesting on the population structure and dynamics of Pinus koraiensis (Pinaceae). Acta Ecol. Sin. 2015, 35, 38–45. [Google Scholar]
- International Union for Conservation of Nature and Natural Resources. Available online: https://www.iucnredlist.org/species/42373/2975987 (accessed on 13 December 2010).
- Lacaze, J.F.; Fao, R.F.D. Advances in species and provenance selection. Unasylva 1978, 31, 371–375. [Google Scholar]
- Lauteri, M.; Scartazza, A.; Guido, M.C.; Brugnoli, E. Genetic variation in photosynthetic capacity, carbon isotope discrimination and mesophyll conductance in provenances of Castanea sativa adapted to different environments. Funct. Ecol. 2003, 11, 675–683. [Google Scholar] [CrossRef]
- Reinhardt, K.; Castanha, C.; Germino, M.J.; Kueppers, L.M. Ecophysiological variation in two provenances of Pinus flexilis seedlings across an elevation gradient from forest to alpine. Tree Physiol. 2011, 31, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Hou, D.; Zhang, L.; Wei, Z.; Zhou, C.; Xia, D. Genetic variation of Pinus koraiensis from provenances and high carbon storage provenance selection. Bull. Bot. Res. 2016, 36, 452–460. [Google Scholar]
- Gao, J.; Zhang, H. Provenance selection test of building materials in Pinus koraiensis. For. Technol. Dev. 2004, 2, 20–22. [Google Scholar]
- Wang, Y.; Yang, D.; Wang, Z.; Li, G.; Gao, W.; Yang, F. Geographical variation law of biennia Pinus koraiensis seedlings and selection of best provenance in Maoershan Mountain. For. Sci. Technol. 1995, 20, 15–16. [Google Scholar]
- Zhang, Q.; Wang, H.; Jiang, G.; Shen, G.; Wang, L.; Li, Y.; Wang, L.; Wang, L.; LI, Y.; Li, R.; et al. Variation analysis and selection of Pinus koraiensis half-sib families. Bull. Bot. Res. 2019, 39, 557–567. [Google Scholar]
- Wang, F.; Wang, Y.; Wang, C.; Zhang, W.; Liu, W.; Lu, Z.; Yang, Y. Variation of the growth, fruiting and resistance to disease and insect of the half-sib families of Pinus koraiensis superior trees. Chin. J. Appl. Ecol. 2019, 30, 1679–1686. [Google Scholar]
- Wang, Z.; Hao, X.; Hu, X.; Yang, D.; Lang, F.; Wang, D.; Mu, H. Family selection of Pinus koraiensis based on growth and fructification. J. Beihua Univ. Nat. Sci. 2019, 20, 26–32. [Google Scholar]
- Liang, D.; Ding, C.; Zhao, G.; Leng, W.; Zhang, M.; Zhao, X.; Qu, G. Variation and selection analysis of Pinus koraiensis clones in northeast China. J. For. Res. 2018, 29, 611–622. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, J.; Zhang, P.; Liang, D.; Zhang, Q.; Wang, B.; Pei, X.; Zhao, X. Variation and selection of growth and fruit traits among 170 Pinus koraiensis clones. For. Res. 2019, 32, 58–64. [Google Scholar]
- Liang, D.; Jin, Y.; Zhao, G.; Dong, Y.; Len, W.; Chen, C.; Wang, H.; Zhao, X. Variance analyses of growth and wood characteristics of 50 Pinus koraiensis clones. J. Beijing Univ. 2016, 38, 51–59. [Google Scholar]
- Yang, J.; Wang, Y.; Wang, Y.; Zhou, C.; Wang, H. Selection of excellent provenance of Pinus koraiensis. Jilin For. Technol. 1998, 14–16. [Google Scholar]
- Wang, Y.; Dong, Y.; Wu, P.; Han, Y.; Wu, D.; Wu, Z. Effect analysis of establishing seedling seed orchard of Pinus koraiensis. Jilin For. Technol. 2007, 36, 1–5. [Google Scholar]
- Wang, X.; Hou, D.; Xia, D.; Yang, C.; Wei, Z. Correlation characteristics variation of Pinus koraiensis provenance and its excellent provenance selection. Prot. For. Sci. Technol. 2016, 11–16. [Google Scholar]
- Xia, D.; Yang, S.; Yang, C.; Lv, Q.; Liu, G.; Zhang, P. Study of Pinus koraiensis provenance test (1):Provenance regionalization. J. Northeast Univ. 1991, 122–128. [Google Scholar]
- Zhou, S.; Lin, S.; Zhang, D.; Zhao, S.; Lv, Z.; Zhao, H.; Zhu, H.; Chen, Y.; Wu, Y.; Zhang, G.; et al. Study on the genetic improvement of Pinus koraiensis Sieb.et Zucc. in river of Lushui and its germplasm preservation and utilization. Chin. Countrys. Well Off Technol. 2010, 59–61. [Google Scholar]
- Wang, H.; Xia, D.; Wang, W.; Yang, S. Genetic variations of wood properties and growth characters of Korean pines from different provenances. J. For. Res. 2002, 13, 277–280. [Google Scholar]
- Hill, W.G. Selective breeding. Brenners Encycl. Genet. 2013, 1, 371–373. [Google Scholar]
- Liu, D. Research progress on the selection and breeding of Pinus koraiensis in China. Prot. For. Sci. Technol. 2017, 96–99. [Google Scholar]
- Hawkins, B.J.; Xue, J.; Bown, H.E.; Clinton, P.W. Relating nutritional and physiological characteristics to growth of Pinus radiata clones planted on a range of sites in New Zealand. Tree Physiol. 2010, 30, 1174–1191. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.J.; Deng, J.; Ting, D.U.; Shi, L.; Zhou, C.L. Superior plant and clone selection and evaluation of introduced dalbergia sissoo provenances. For. Res. 2017, 30, 916–920. [Google Scholar]
- Sharma, R.M.; Kour, K.; Singh, B.; Yadav, S.; Kotwal, N.; Rana, J.C.; Anand, R. Selection and characterization of elite walnut (Juglans regia L.) clone from seedling origin trees in north western himalayan region of India. Aust. J. Crop. Sci. 2014, 8, 257–262. [Google Scholar]
- Liu, Y.; Yan, K.; He, W.; Pan, H. Genetic variation of fiber traits in Populus Deltoides clones. J. Nanjing For. Univ. 2019, 1–11. [Google Scholar]
- Wang, B.; Zhao, X.; Wang, H.; Wang, G.; Shen, G.; Wang, L.; Li, Y.; Lin, J.; Wang, Z. Variance analysis of growth characteristics of 30 Pinus koraiensis half-sib families. J. Northeast Univ. 2019, 47, 10–13. [Google Scholar]
- Mo, C.; Zhang, H.; Zhang, L.; Hou, D.; Zhang, H. Variations in nutrition compositions and morphology characteristics in different hybrid combination of Korean Pine (Pinus koraiensis). Bull. Bot. Res. 2017, 37, 700–708. [Google Scholar]
- Liang, D.; Wang, B.; Song, S.; Wang, J.; Wang, L.; Wang, Q.; Ren, X.; Zhao, X. Analysis of genetic effects on a complete diallel cross test of Pinus koraiensis. Euphytica 2019, 215, 1–12. [Google Scholar] [CrossRef]
- Sweet, G.B. Seed orchards in development. Tree Physiol. 1995, 15, 527–530. [Google Scholar] [CrossRef]
- Svensson, G.P.; Wang, H.-L.; Jirle, E.V.; Rosenberg, O.; Liblikas, I.; Chong, J.M.; Löfstedt, C.; Anderbrant, O. Challenges of pheromone-based mating disruption of Cydia strobilella and Dioryctria abietella in spruce seed orchards. J. Pest Sci. 2018, 91, 639–650. [Google Scholar] [CrossRef]
- Feng, F.; Chen, M.; Zhang, D.; Sui, X.; Han, S. Application of SRAP in the genetic diversity of Pinus koraiensis of different provenances. Afr. J. Biotechnol. 2010, 8, 1000–1008. [Google Scholar]
- Zhang, D. Stuty on Genetic Structure in Different Populations of Pinus Koraiensis by SRAP. Master’s Thesis, Northeast Forestry University, Harbin, China, 2008. [Google Scholar]
- Xia, M.; Zhou, X.; Zhao, S. RAPD analysis on genetic diversity of natural populations of Pinus koraiensis. Acta Ecol. Sin. 2000, 21, 730–737. [Google Scholar]
- Kim, B.Z.S.; Hwang, J.W.; Lee, S.W.; Yang, C.; Gorovoy, P.G. Genetic variation of Korean Pine (Pinus koraiensis Sieb. et Zucc.) at allozyme and RAPD markers in Korea, China and Russia. Silvae Genet. 2005, 54, 235–246. [Google Scholar] [CrossRef]
- Feng, F.; Sui, X.; Zhang, D. Study on genetic diversity of Korean pine from different provenance. For. Sci. Technol. 2008, 33, 1–4. [Google Scholar]
- Jiang, Y. Genetic Diversity of Piuns Koraiensis Populations from Hei Longjiang Area Revealed by ISSR Analysis. Master’s Thesis, Liaoning Normal University, Dalian, China, 2010. [Google Scholar]
- Jia, J. Analysis of Genetic Diversity of Jilin Natural Pinus Koraiensis Population by ISSR Marker. Master’s Thesis, Liaoning Normal University, Dalian, China, 2011. [Google Scholar]
- Shao, D.; Pei, Y.; Zhang, H. cpSSR analysis of variation of genetic diversity in temporal dimension of natural population of Pinus koraiensis in Liangshui National Nature Reserve. Bull. Bot. Res. 2007, 27, 473–477. [Google Scholar]
- Zhang, Y.; Yi, X.; Ji, L. Screening of Pinus koraiensis microsatellite makers from relative species of Pinus and analysis of population genetic diversity. Chin. J. Ecol. 2013, 32, 2307–2313. [Google Scholar]
- Tong, Y.W.; Lewis, B.J.; Zhou, W.M.; Mao, C.R.; Wang, Y.; Zhou, L.; Yu, D.P.; Dai, L.M.; Qi, L. Genetic diversity and population structure of natural Pinus koraiensis populations. Forests 2020, 11, 39. [Google Scholar] [CrossRef]
- Jia, D.; Zhen, Z.; Zhang, H.; Tang, J. EST–SSR marker development and transcriptome sequencing analysis of different tissues of Korean pine (Pinus koraiensis Sieb. et Zucc.). Biotechnol. Biotechnol. Equip. 2017, 31, 1–11. [Google Scholar] [CrossRef]
- Yu, J.-H.; Chen, C.-M.; Tang, Z.-H.; Yuan, S.-S.; Wang, C.-J.; Zu, Y.-G. Isolation and characterization of 13 novel polymorphic microsatellite markers for Pinus koraiensis (Pinaceae). Am. J. Bot. 2012, 99, e421–e424. [Google Scholar] [CrossRef]
- Li, Y.; Liang, L.; Ge, X.J. Development of microsatellite loci for Pinus koraiensis (Pinaceae). Am. J. Bot. 2010, 97, e39–e41. [Google Scholar] [CrossRef]
- Yun, J.; Juan, Z.; Ying, W.; Wei-Bing, F.; Gui-Fang, Z.; Zhong-Hu, L. Effects of geological and environmental events on the diversity and genetic divergence of four closely related Pines: Pinus koraiensis, P. armandii, P. griffithii, and P. pumila. Front. Plant Sci. 2018, 9, 1264. [Google Scholar]
- Aizawa, M.; Kim, Z.; Yoshimaru, H. Phylogeography of the Korean pine (Pinus koraiensis) in northeast Asia: Inferences from organelle gene sequences. J. Plant Res. 2012, 125, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Zha, M.; Imran, M.; Wang, Y.; Xu, J.; Ding, Y.; Wang, S. Transcriptome analysis revealed the interaction among strigolactones, auxin, and cytokinin in controlling the shoot branching of rice. Plant Cell Rep. 2019, 38, 279–293. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, X.; Li, Q.; Chen, X.; Li, X. Comparative transcriptome analysis to elucidate the enhanced thermotolerance of tea plants (Camellia sinensis) treated with exogenous calcium. Planta 2019, 249, 775–786. [Google Scholar] [CrossRef] [PubMed]
- Fox, H.; Doron-Faigenboim, A.; Kelly, G.; Bourstein, R.; David-Schwartz, R. Transcriptome analysis of Pinus halepensis under drought stress and during recovery. Tree Physiol. 2017, 38, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Meng, L.; Lu, H.; Zhiyong, Z.; Xiaoming, P.; Yingyue, L. De novo transcriptome assembly and population genetic analyses for an endangered Chinese endemic Acer miaotaiense (Aceraceae). Genes 2018, 9, 378. [Google Scholar] [CrossRef] [PubMed]
- Zhen, Z.; Hanguo, Z.; Chi, M.; Lei, Z. Transcriptome sequencing analysis and development of EST-SSR markers for Pinus koraiensis. Sci. Silvae Sin. 2015, 51, 114–120. [Google Scholar]
- Ran, L. The Mechanism of Pine Polyphenols Synthesis in Pinus Koraiensis Cell Regulated by Elicitor. Ph.D. Thesis, Noreheast Forestry University, Harbin, China, 2015. [Google Scholar]
- Chen, M.; Feng, F.; Sui, X.; Han, S. Comparison of application of three molecular marker methods in map construction of Pinus koraiensis. Nonwood For. Res. 2011, 29, 13–16. [Google Scholar]
- Schaedle, M. Tree photosynthesis. Annu. Rev. Plant Physiol. 2003, 26, 101–115. [Google Scholar] [CrossRef]
- Tanai, C.; Shengxi, S.; Peter, J.N. Enhancing photosynthesis in plants: The light reactions. Essays Biochem. 2018, 62, 85–94. [Google Scholar]
- Sui, X.; Shan, N.; Hu, L.; Zhang, C.; Yu, C.; Ren, H.; Turgeon, R.; Zhang, Z. The complex character of photosynthesis in cucumber fruit. J. Exp. Bot. 2017, 68, 1625–1637. [Google Scholar] [CrossRef]
- Sello, S.; Meneghesso, A.; Alboresi, A.; Baldan, B.; Morosinotto, T. Plant biodiversity and regulation of photosynthesis in the natural environment. Planta 2019, 249, 1217–1228. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Wang, W.Y.; Meng, J.X.; Liu, H.; Xing, Z.; Mao, J.-F.; Wang, X.-R.; Li, Y. Comparison of the photosynthetic characteristics of hybrid and parental species of Pinus tabuliformis ×P. yunnanensis and P. densata. J. Beijing Univ. 2016, 38, 37–43. [Google Scholar]
- Takagi, K.; Hirata, R.; Ide, R.; Ueyama, M.; Ichii, K.; Saigusa, N.; Hirano, T.; Asanuma, J.; Li, S.-G.; Machimura, T. Spatial and seasonal variations of CO2 flux and photosynthetic and respiratory parameters of larch forests in East Asia. Soil Sci. Plant Nutr. 2014, 61, 61–75. [Google Scholar] [CrossRef]
- Ambrosini, V.G.; Rosa, D.J.; Melo, G.W.B.D.; Zalamena, J.; Brunetto, G. High copper content in vineyard soils promotes modifications in photosynthetic parameters and morphological changes in the root system of ‘Red Niagara’ plantlets. Plant Physiol. Biochem. 2018, 128, 89–98. [Google Scholar] [CrossRef]
- Shang, B.; Feng, Z.; Li, P.; Yuan, X.; Xu, Y.; Calatayud, V. Ozone exposure-and flux-based response relationships with photosynthesis, leaf morphology and biomass in two poplar clones. Sci. Total Environ. 2017, 603, 185–195. [Google Scholar] [CrossRef]
- Cheng, X.B.; Wu, J.; Han, S.J.; Zhou, Y.M.; Wang, X.X.; Wang, C.G.; Zhao, J.; Hu, Q.H. Photosynthesis, leaf morphology and chemistry of Pinus koraiensis and Quercus mongolicain broadleaved Korean pine mixed forest. Photosynthetica 2012, 50, 56–66. [Google Scholar] [CrossRef]
- Zhou, Y.M.; Han, S.J. Photosynthetic response and stomatal behaviour of Pinus koraiensis during the fourth year of exposure to elevated CO2 concentration. Photosynthetica 2005, 43, 445–449. [Google Scholar] [CrossRef]
- Huo, H.; Wang, C. Effects of canopy position and leaf age on photosynthesis and transpiration of Pinus koraiensis. Chin. J. Appl. Ecol. 2007, 18, 9–14. [Google Scholar]
- Dong, Y.; Liu, Y. Changes in the response of leaf traits in Pinus koraiensis (Korean pine) seedlings of different ages to controlled temperatures and light conditions. Acta Ecol. Sin. 2017, 37, 5662–5672. [Google Scholar]
- Wang, W.J.; Zu, Y.G.; Wang, H.M.; Li, X.Y.; Koike, T. Newly-formed photosynthates and the respiration rate of girdled stems of Korean pine (Pinus koraiensis Sieb. et Zucc.). Photosynthetica 2006, 44, 147–150. [Google Scholar] [CrossRef]
- Sun, Y.; Zhu, J.; Yu, L.; Yan, Q.; Wang, K. Photosynthetic characteristics of Pinus koraiensis seedlings under different light regime. Chin. J. Ecol. 2009, 28, 72–79. [Google Scholar]
- Mou, W.; Yan, T.; Li, J.; Meng, W. Overview of studies on photosynthesis of Korean pine. Agric. Internet Inf. 2009, 99–102. [Google Scholar]
- Sun, Y.; Zhu, J.; Sun, O.J.; Yan, Q. Photosynthetic and growth responses of Pinus koraiensis seedlings to canopy openness: Implications for the restoration of mixed-broadleaved Korean pine forests. Environ. Exp. Bot. 2016, 129, 118–126. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, K.; Sun, Y.; Yan, Q. Response of Pinus koraiensis seedling growth to different light conditions based on the assessment of photosynthesis in current and one-year-old needles. J. Res. 2014, 25, 53–62. [Google Scholar] [CrossRef]
- Liang, D.; Jiang, Y.; Zhao, G.; Dong, Y.; Len, W.; Zhao, X. Comparative study of photosynthetic characteristics of Pinus koraiensis clones. Genomics Appl. Biol. 2018, 37, 3996–4006. [Google Scholar]
- Wang, M. Comparison of photosynthetic and growth characteristics between Korean pine and Pinus sibirica seedlings. J. Liaoning For. Sci. Technol. 2017, 35, 33–35. [Google Scholar]



{kind=link}
{kind=link}
| Study Level | Number | Trait | Name of Superior Provenances/Improved Varieties | References |
|---|---|---|---|---|
| Provenance | 18 | Growth, wood characteristics and carbon storage | Lushuihe, Linjiang, Dahailin | [15] |
| Provenance | 23 | wood characteristics | Caohekou, Wangqing, Dahailin | [16] |
| Provenance | 17 | Growth traits | Caohekou, Bajiazi, Changbaixian | [17] |
| Family | 53 | Growth traits and shape traits | 115, 117, 138, 133, 112 | [18] |
| Family | 551 | Growth, fruiting and resistance to disease | PK315, PK187, PK539, PK547, PK544, PK541, PK520, PK156, PK323, PK534, PK177, PK132, PK146, PK540, PK139, PK46, PK207, PK118, PK431, PK105, PK170, PK296, PK246, PK537, PK377, PK288, PK101, PK543 | [19] |
| Family | 68 | Growth and fruit traits | 50, 112, 107, 95 | [20] |
| Clone | 50 | Growth traits | PK11, PK19, PK04, PK14, PK28 | [21] |
| Clone | 170 | Growth and fruit traits | 14, 84, 83, 138, 85, 82, 1110, 88, 123, 126, 820, 137, 81, 86, 11, 12, 810 | [22] |
| Clone | 50 | Growth and wood characteristics | PK6, PK47, PK15, PK37, PK27, PK6, PK47, PK15, PK37, PK27 | [23] |
| Codes | Establishment Time of Base | Location | Longitude | Latitude | Altitude (m) | Improvement Base |
|---|---|---|---|---|---|---|
| L1 | 1974 | Benxi, Liaoning | 124°12′33.80″ | 41°26′59.87″ | 276 | Primary |
| L2 | 1968 | Benxi, Liaoning | 124°20′57.16″ | 40°10′50.76″ | 616 | Primary |
| J1 | 1972 | Wangqing, Jilin | 129°45′37.57″ | 43°19′47.93″ | 320 | Primary |
| J2 | 1958 | Tonghua, Jilin | 125°33′41.71″ | 41°52′57.50″ | 543 | Primary |
| J3 | 1985 | Longjin, Jilin | 129°45′20.18″ | 42°42′11.96″ | 164 | Primary |
| J4 | 1984 | Baishan, Jilin | 127°48′30.16″ | 42°31′36.87″ | 745 | Primary, 2 generation |
| J5 | 1978 | Linjiang, Jilin | 126°50′55.32″ | 41°45′12.02″ | 324 | Primary |
| J6 | 1989 | Baishan, Jilin | 126°49′06.24″ | 42°36′12.41″ | 606 | Primary |
| H1 | 1958 | Ningan, Heilongjiang | 128°42′29.24″ | 44°10′33.05″ | 585 | Primary |
| H2 | 1965 | Hegang, Heilongjiang | 130°13′59.02″ | 47°24′25.03″ | 197 | Primary |
| H3 | 1978 | Jiamusi, Heilongjiang | 130°40′47.06″ | 46°25′46.80″ | 201 | Primary |
| H4 | 1973 | Yichun, Heilongjiang | 130°13′61.05″ | 46°50′08.12″ | 465 | Primary, 2 generation |
| H5 | 1981 | Shangzhi, Heilongjiang | 130°13′62.21″ | 44°40′21.72″ | 398 | Primary, 1.5 generation |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Liu, X.-T.; Wei, J.-T.; Li, Y.; Tigabu, M.; Zhao, X.-Y. Genetic Improvement of Pinus koraiensis in China: Current Situation and Future Prospects. Forests 2020, 11, 148. https://doi.org/10.3390/f11020148
Li X, Liu X-T, Wei J-T, Li Y, Tigabu M, Zhao X-Y. Genetic Improvement of Pinus koraiensis in China: Current Situation and Future Prospects. Forests. 2020; 11(2):148. https://doi.org/10.3390/f11020148
Chicago/Turabian StyleLi, Xiang, Xiao-Ting Liu, Jia-Tong Wei, Yan Li, Mulualem Tigabu, and Xi-Yang Zhao. 2020. "Genetic Improvement of Pinus koraiensis in China: Current Situation and Future Prospects" Forests 11, no. 2: 148. https://doi.org/10.3390/f11020148
APA StyleLi, X., Liu, X.-T., Wei, J.-T., Li, Y., Tigabu, M., & Zhao, X.-Y. (2020). Genetic Improvement of Pinus koraiensis in China: Current Situation and Future Prospects. Forests, 11(2), 148. https://doi.org/10.3390/f11020148