Assessment of the Response of a Scots Pine Tree to Effective Wind Loading


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
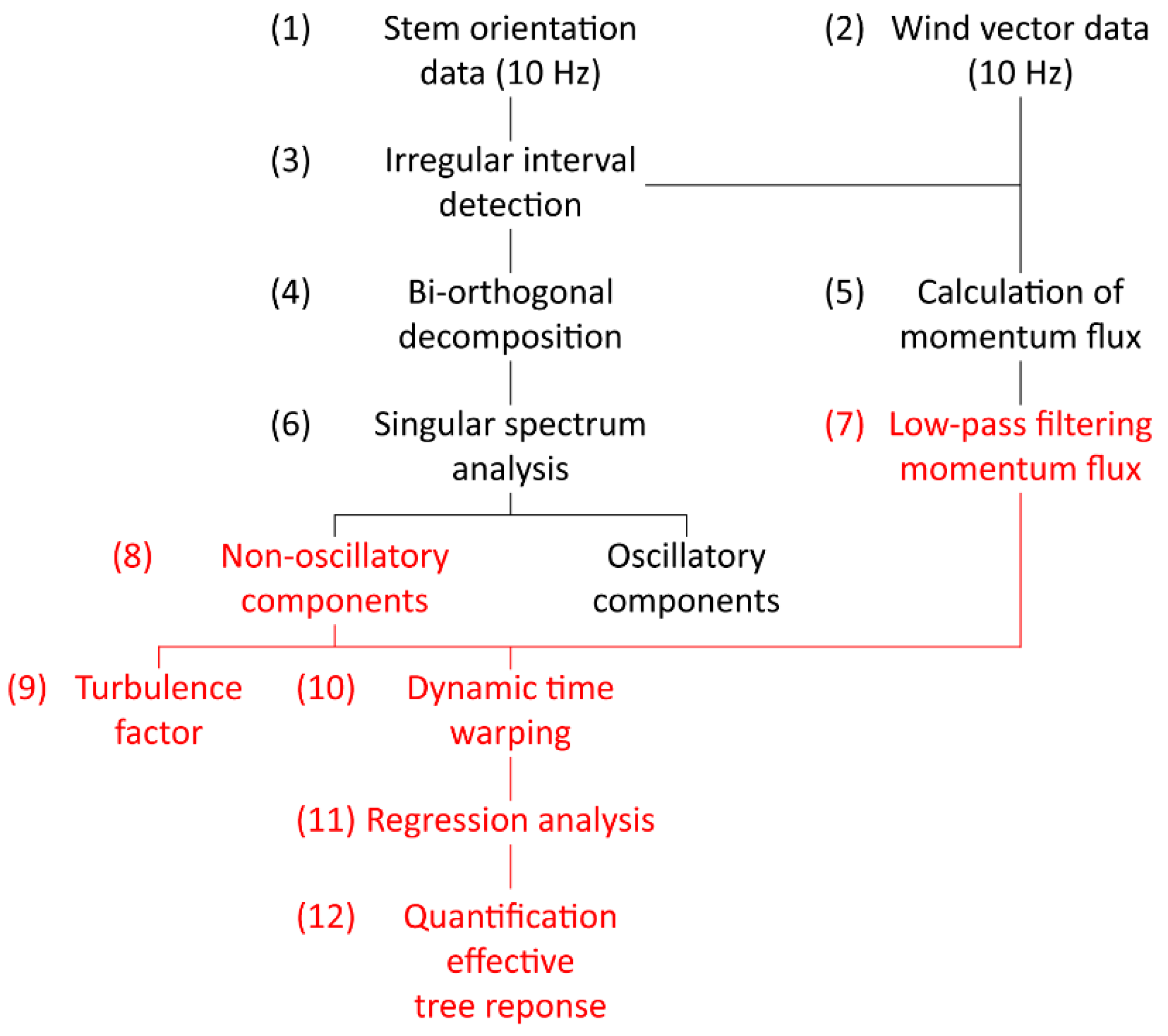
2.1. Workflow
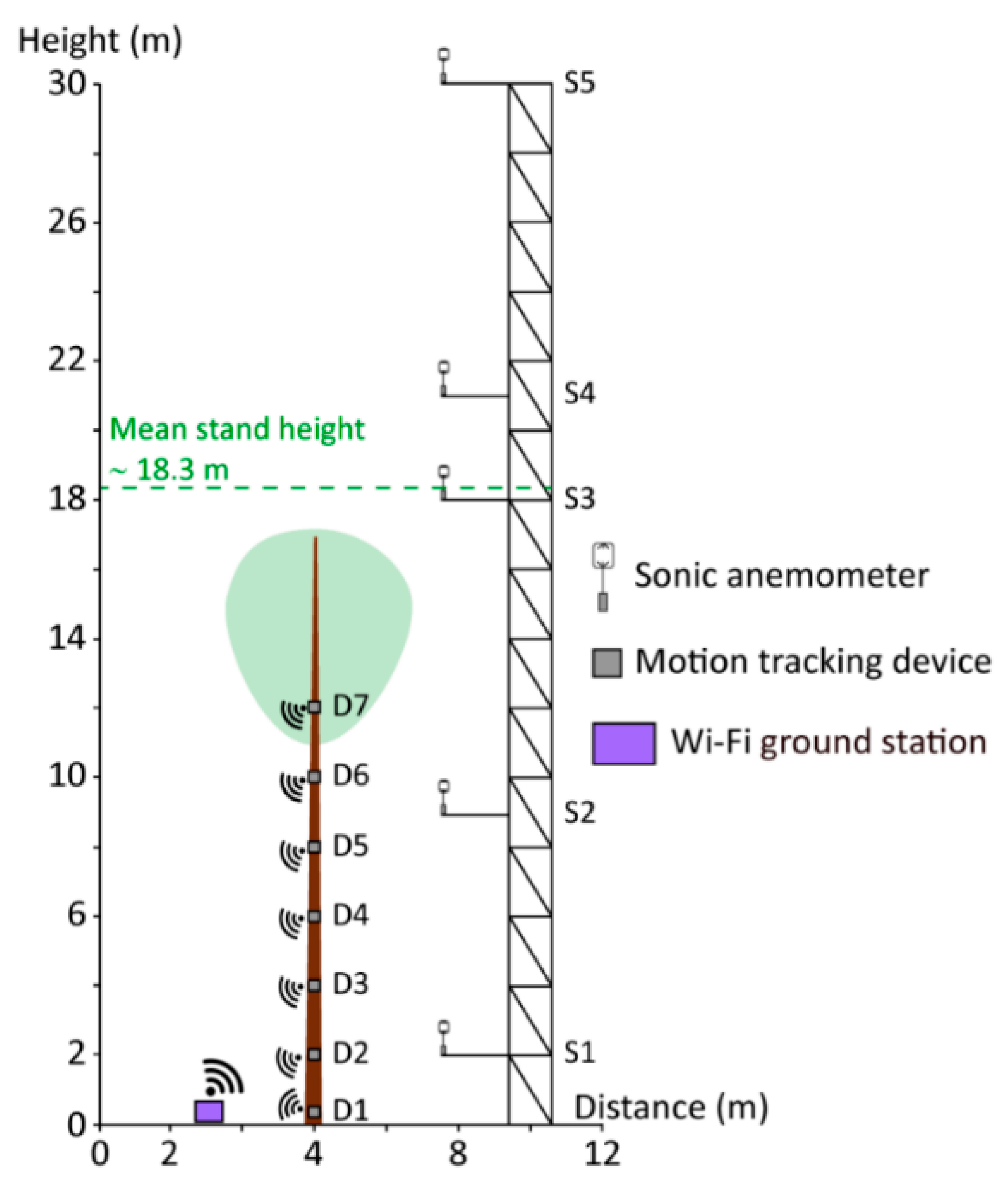
2.2. Airflow and Stem Orientation Measurements
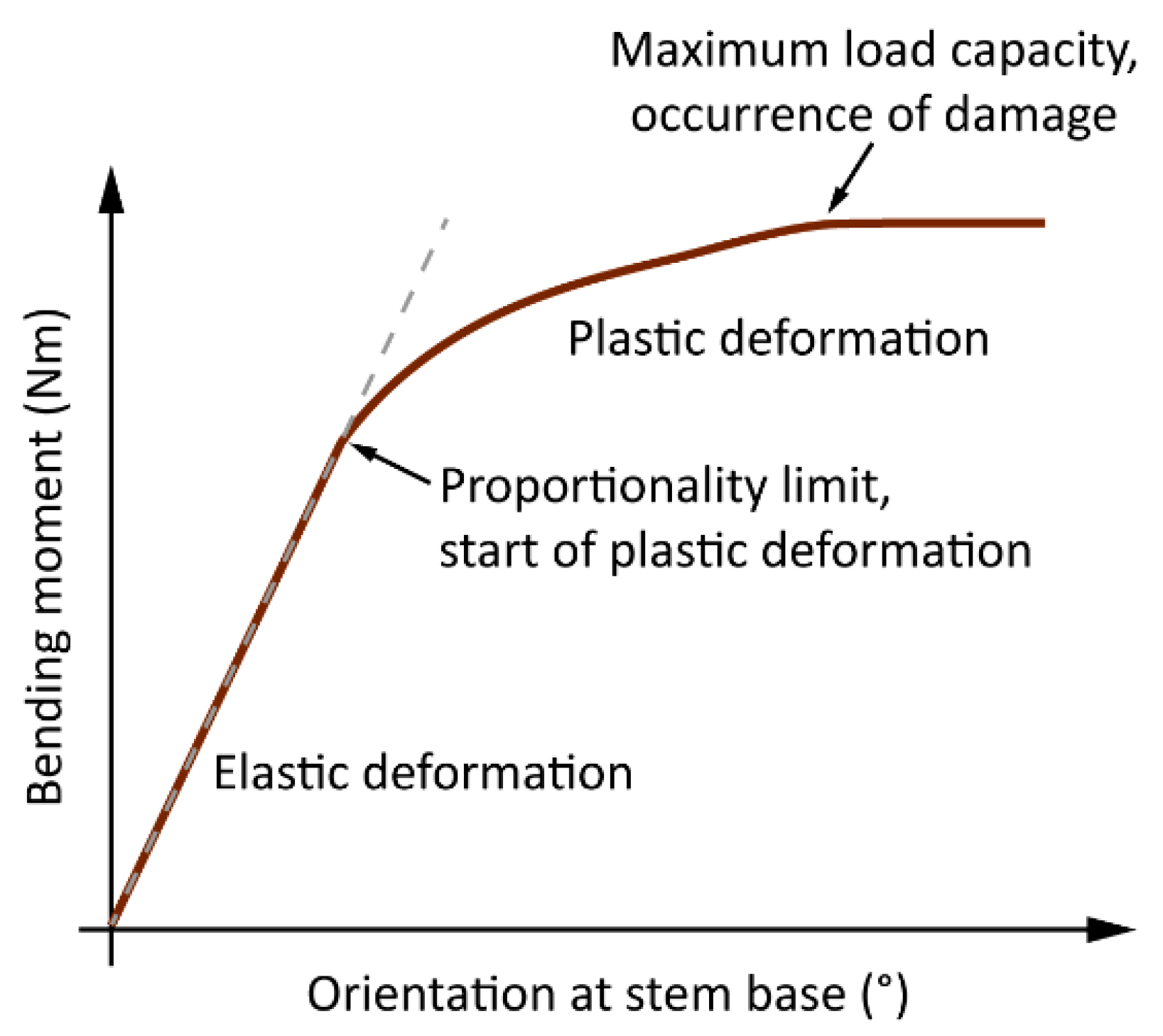
2.3. Analysis of Wind-Induced Tree Response
2.4. Bi-Orthogonal Decomposition
2.5. Singular Spectrum Analysis
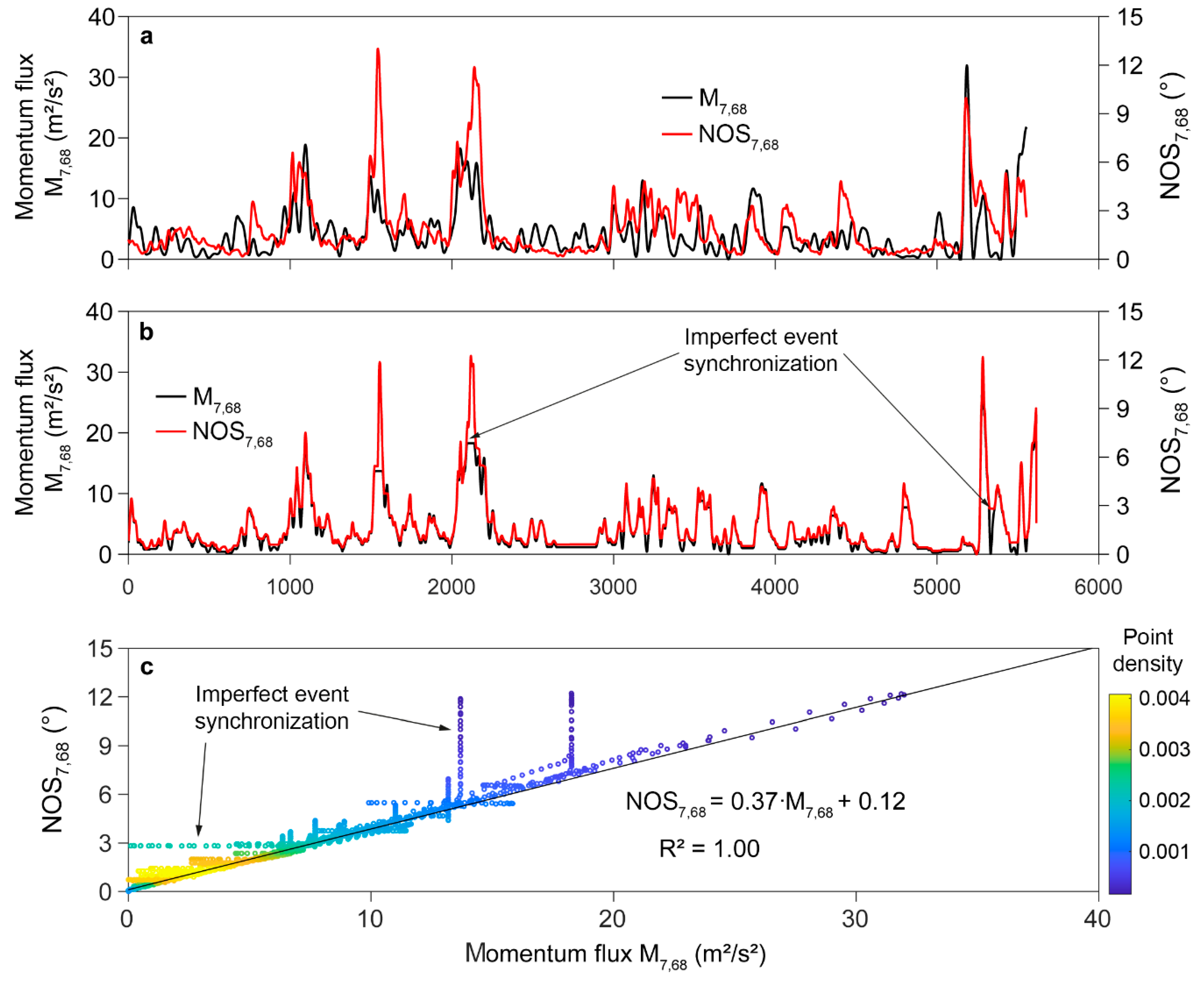
2.6. Dynamic Time Warping
2.7. Turbulence Factor Calculation
3. Results and Discussion
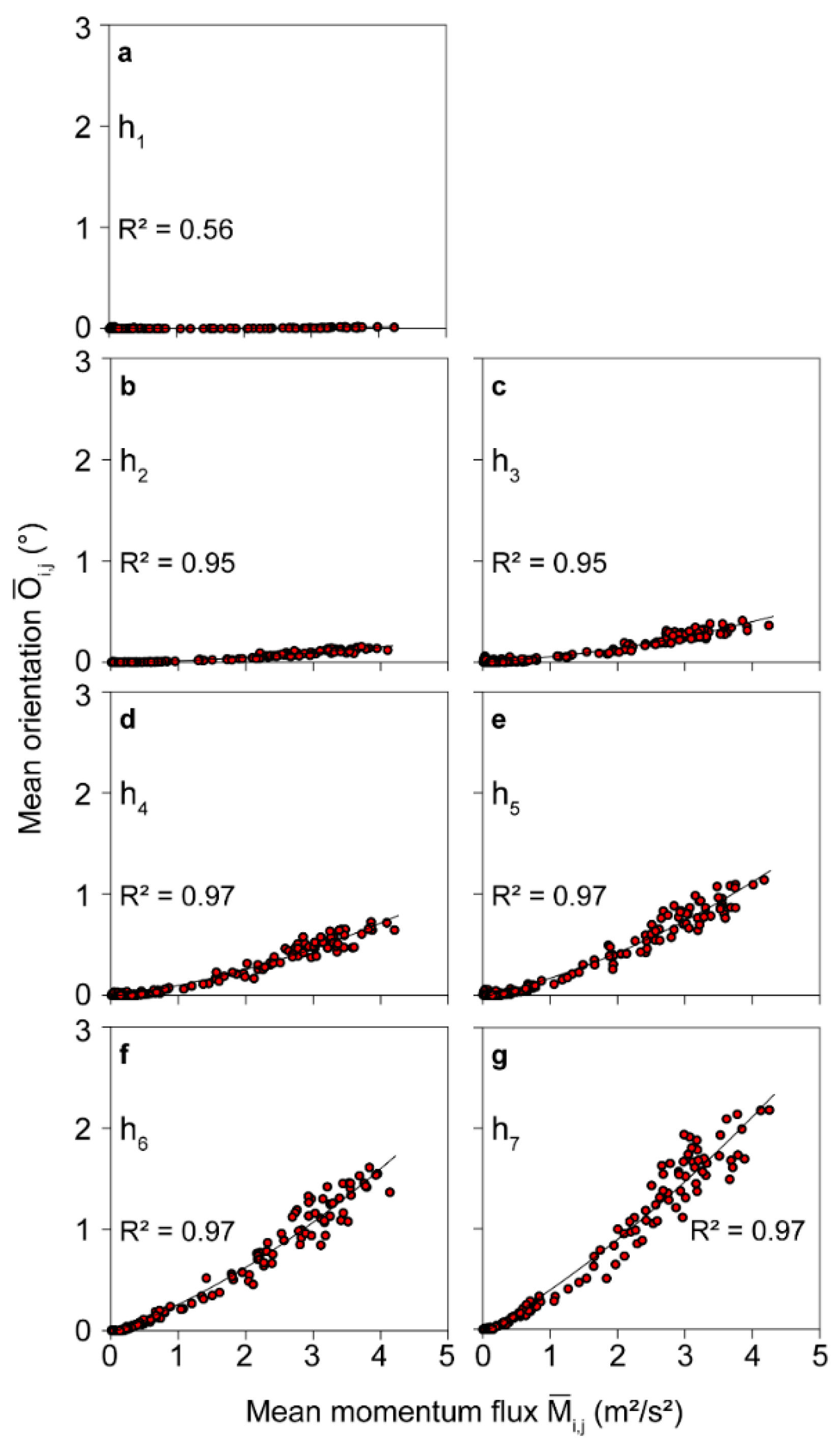
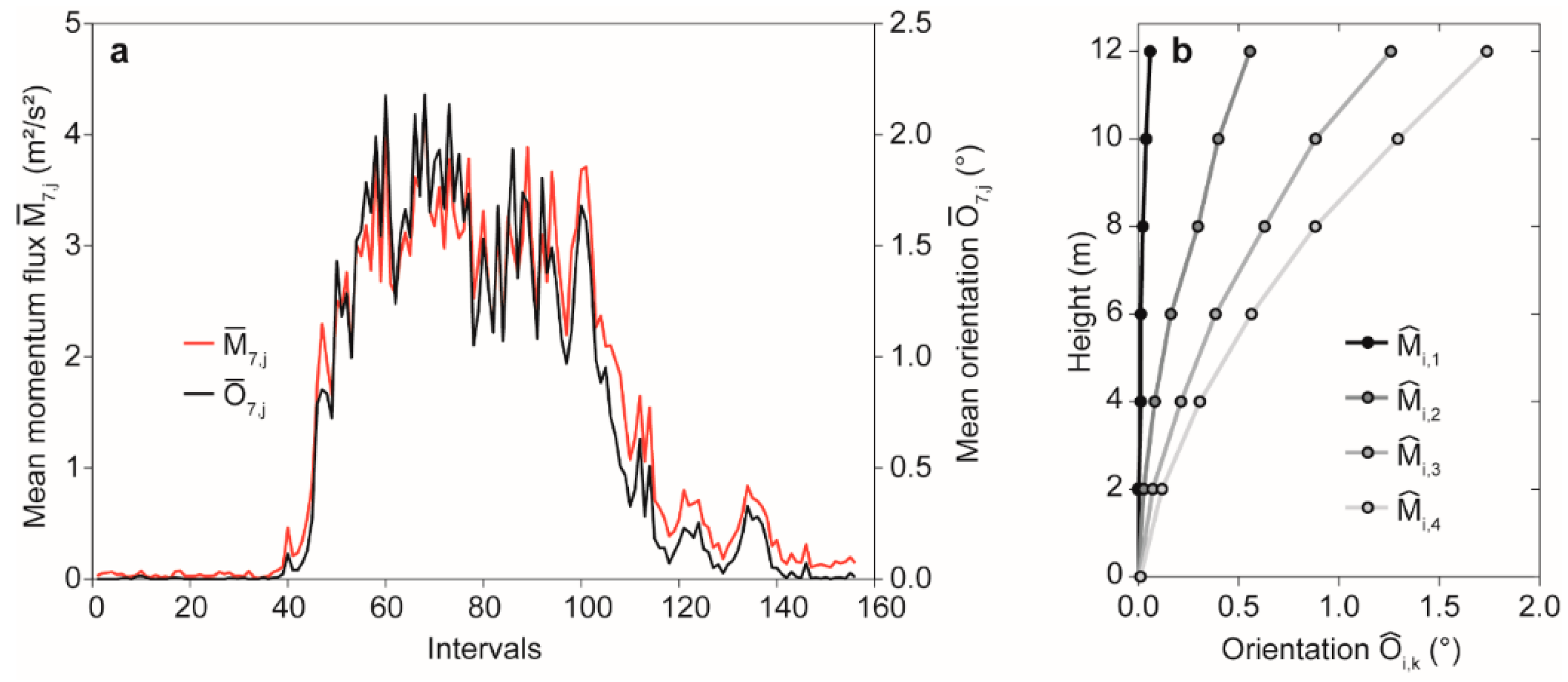
3.1. Mean Momentum Flux-Induced Tree Response
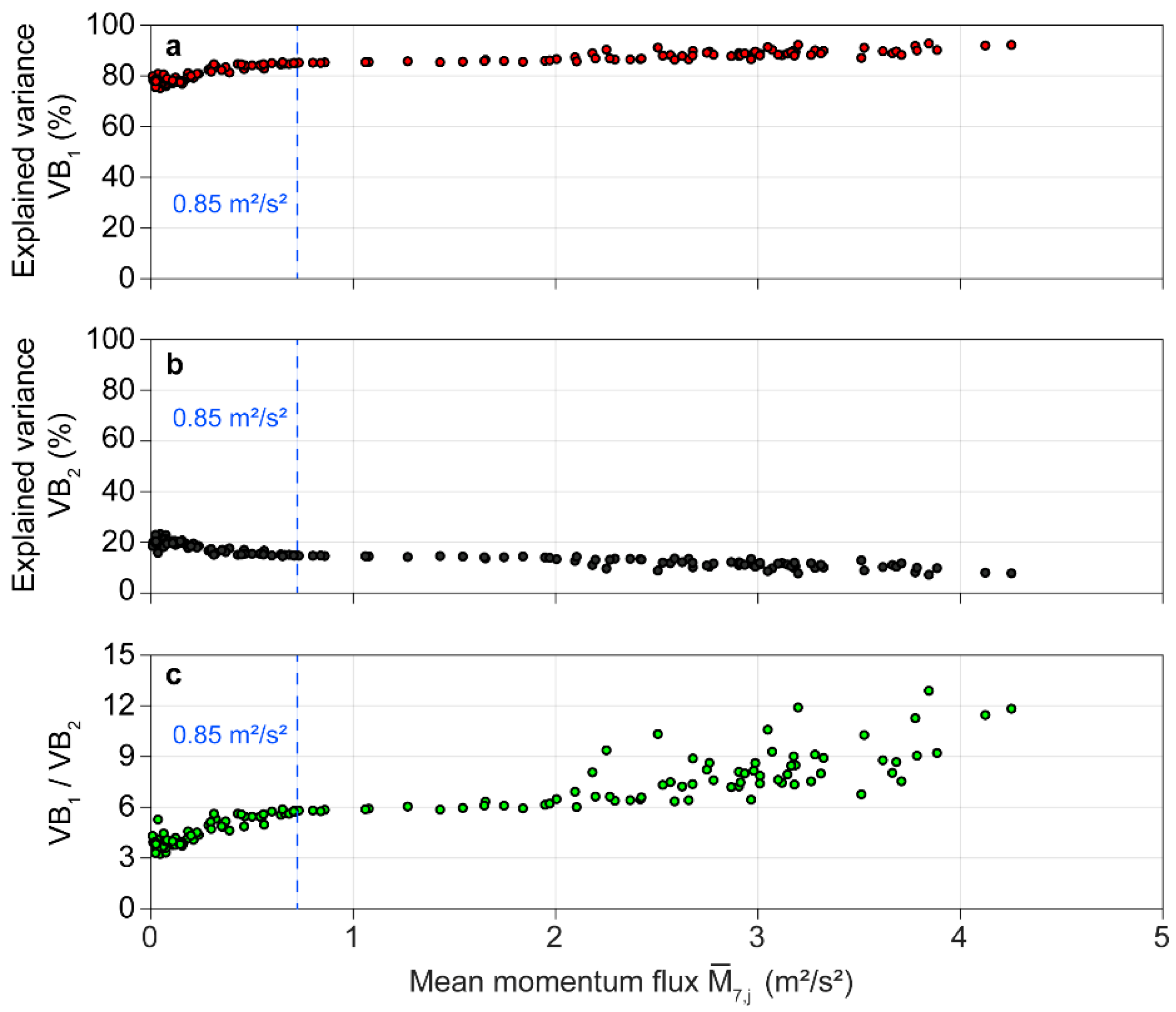
3.2. Bi-Orthogonal Decomposition
3.3. Singular Spectrum Analysis
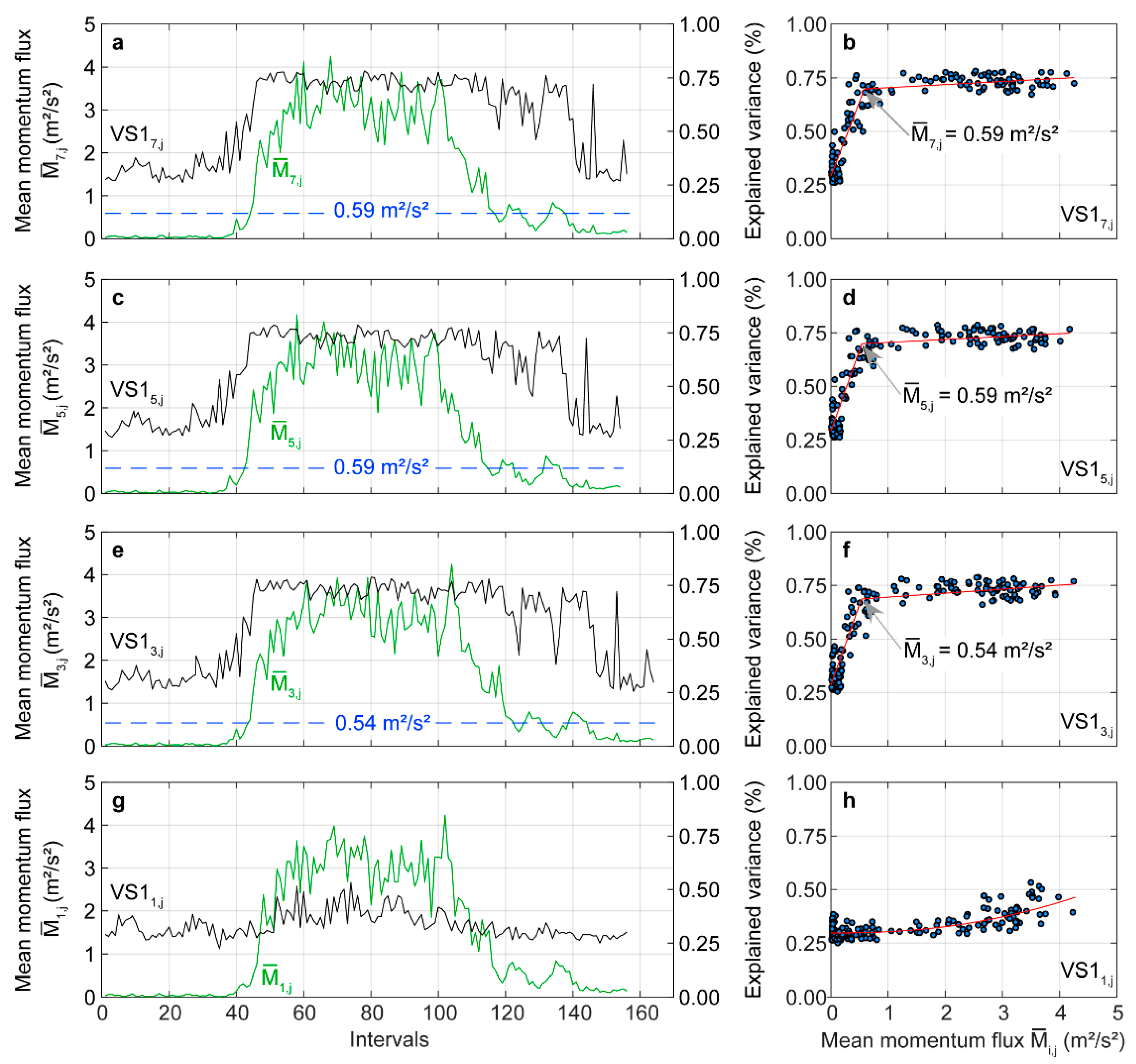
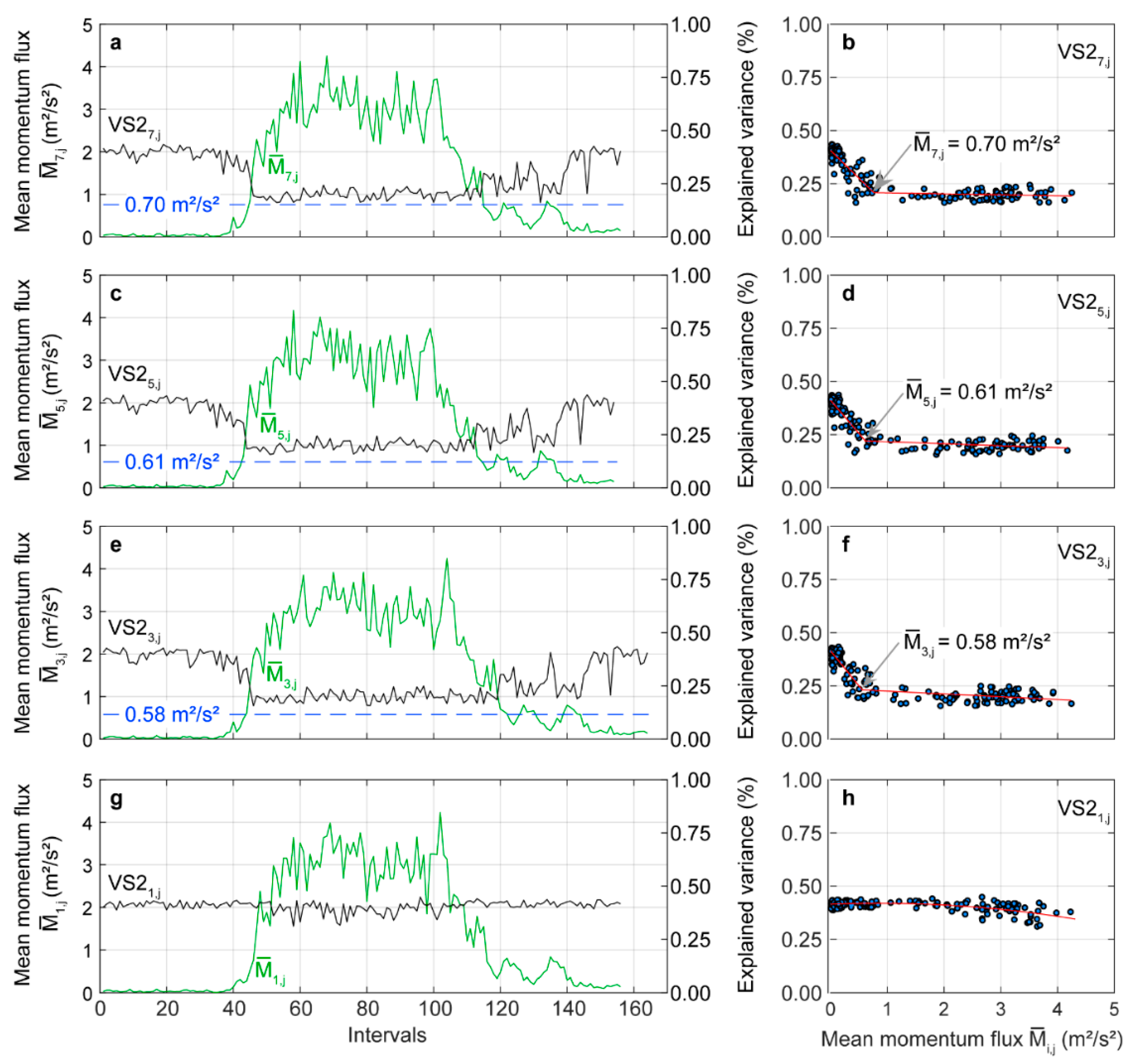
3.4. Dynamic Time Warping
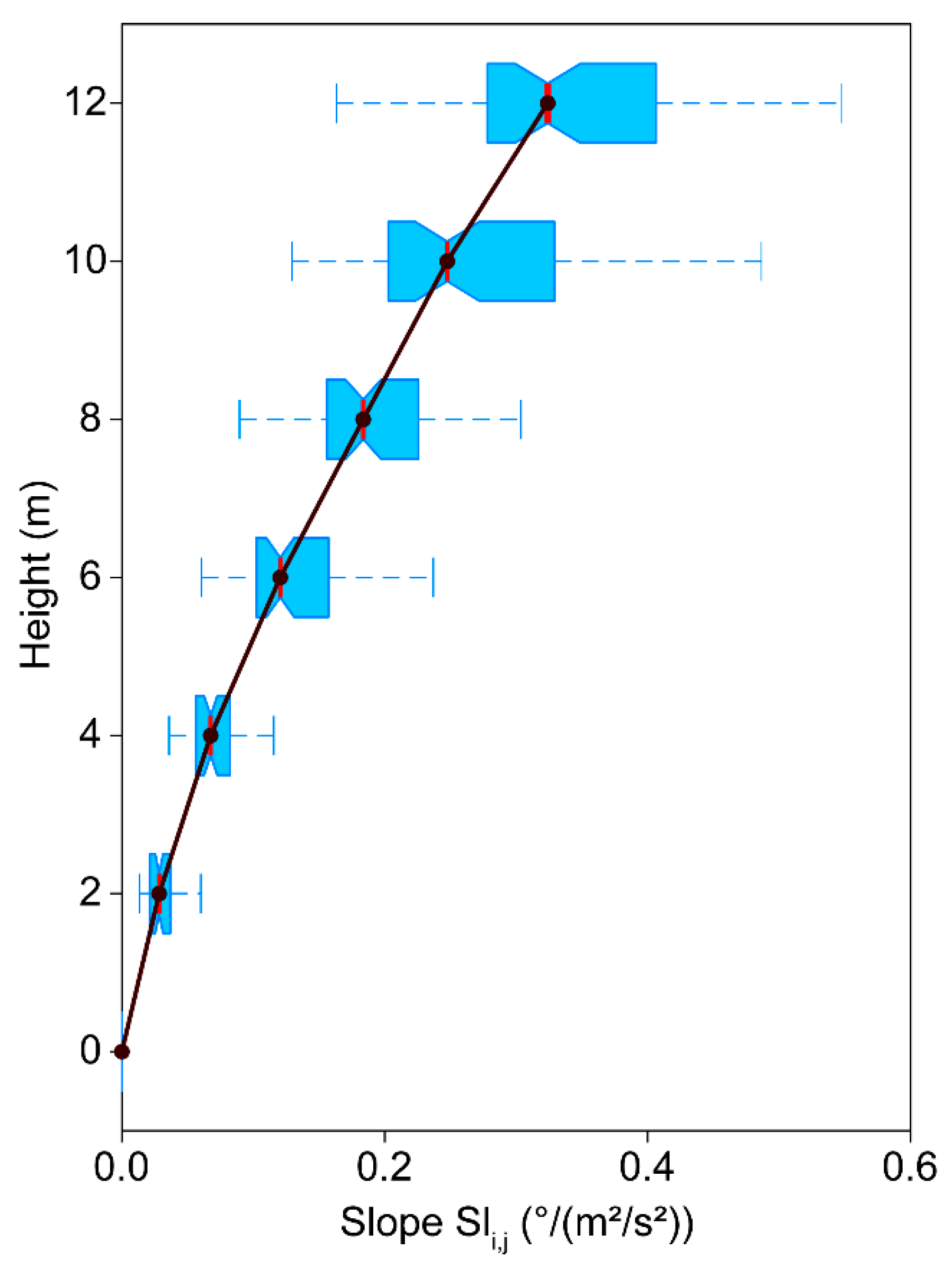
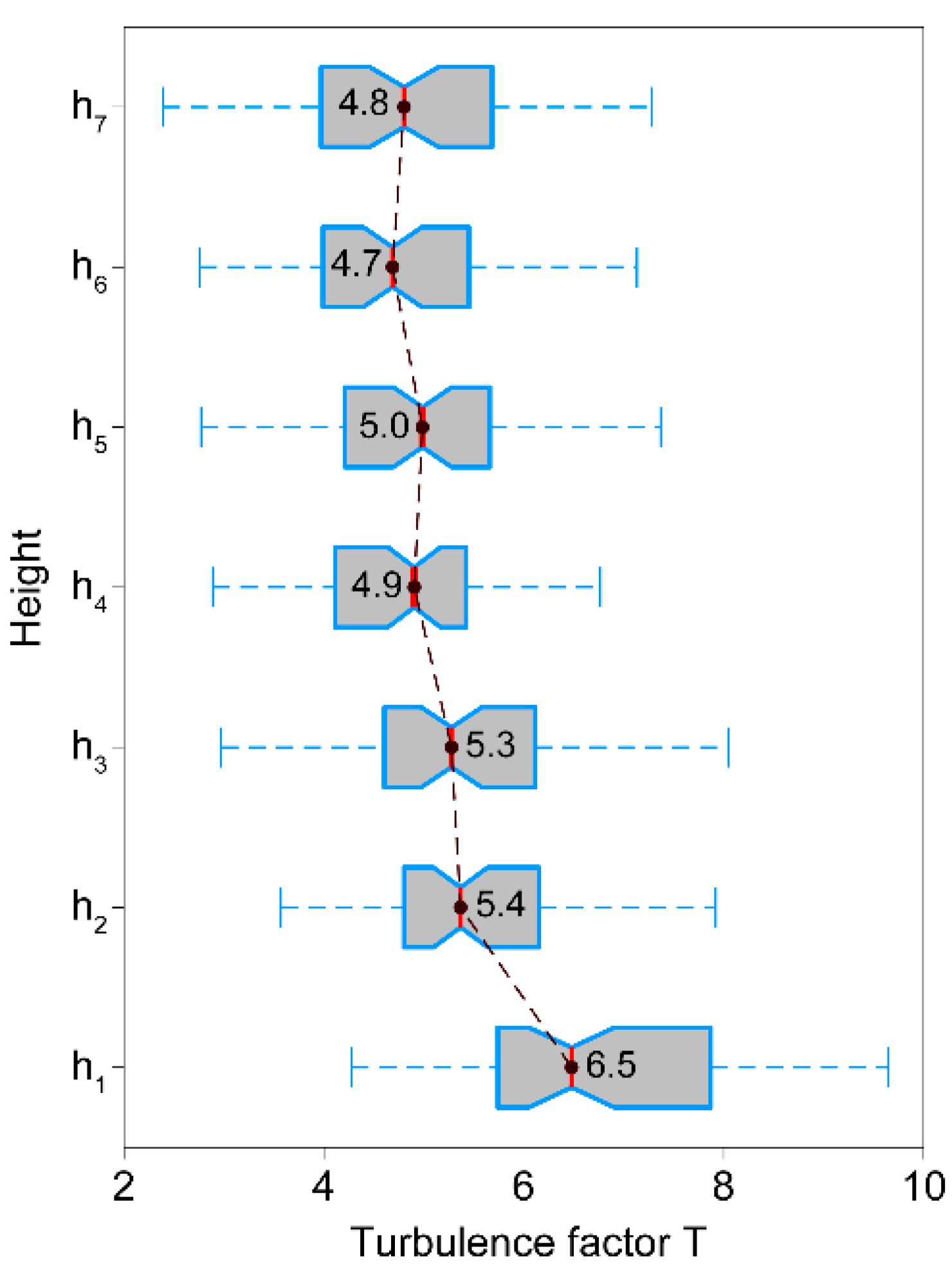
3.5. Turbulence Factor
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Nomenclature
| Acronyms | |
| BOD | bi-orthogonal decomposition |
| CWS | critical wind speed |
| D1-D7 | seven MEMS motion-tracking devices installed at to |
| MEMS | micro electro-mechanical systems |
| S1-S5 | five ultrasonic anemometers installed on the meteorological measurement tower |
| S3 | ultrasonic anemometer installed close to canopy top |
| SSA | singular spectrum analysis |
| Symbols | |
| Euclidean distance | |
| fundamental sway frequency of the stem (Hz) | |
| gravity acceleration (m/s2) | |
| gust factor | |
| mounting heights ( = 1, …, 7) of D1-D7 along the stem of the Scots pine tree (m) | |
| embedding dimension | |
| momentum flux in height i and interval j (m2/s2) | |
| interval mean of (m2/s2) | |
| mean of assigned to four classes (m2/s2) | |
| non-oscillatory sway component determined with SSA in height i and interval j (°) | |
| 50th percentile (median) of (°) | |
| 98th percentile of (°) | |
| orientation in height i and interval j (°) | |
| interval mean of (°) | |
| mean of assigned to four classes (°) | |
| oscillatory sway component determined with SSA in height i and interval j (°) | |
| coefficient of determination | |
| slope of regression line calculated between and low-pass filtered (°/(m2/s2)) | |
| turbulence factor calculated in height i and interval j | |
| wind vector component in east-west direction in height i and interval j (m/s) | |
| fluctuation of (m/s) | |
| wind vector component in north-south direction in height i and interval j (m/s) | |
| fluctuation of (m/s) | |
| variance explained by BOD components in interval j (%) | |
| variance explained by SSA components in height i and interval j (%) | |
| variance explained by (%) | |
| variance explained by (%) | |
| vertical wind vector component in height i and interval j (m/s) | |
| fluctuation of (m/s) | |
| orientation component in east-west direction in height i and interval j (°) | |
| fluctuation (°) | |
| orientation component in north-south direction in height i and interval j (°) | |
| fluctuation of (°) | |
| Subscripts | |
| index for MEMS motion tracking devices | |
| index for horizontal moves in dynamic time warping | |
| index for vertical moves in dynamic time warping | |
| index for irregular, device-specific analysis intervals | |
| index for mean momentum flux class | |
| index for momentum flux sample in dynamic time warping | |
| maximum of | |
| index for orientation sample in dynamic time warping | |
| maximum of | |
References
- Gardiner, B.; Blennow, K.; Carnus, J.-M.; Fleischer, P.; Ingemarson, F.; Landmann, G.; Lindner, M.; Marzano, M.; Nicoll, B.; Orazio, C.; et al. Destructive storms in European forests: Past and forthcoming impacts. Available online: https://ec.europa.eu/environment/forests/pdf/STORMS%20Final_Report.pdf (accessed on 19 June 2019).
- Gregow, H.; Laaksonen, A.; Alper, M.E. Increasing large scale windstorm damage in Western, Central and Northern European forests, 1951–2010. Sci. Rep. 2017, 7, 46397. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.; Schindler, D. Historical winter storm atlas for Germany (GeWiSA). Atmosphere 2019, 10, 387. [Google Scholar] [CrossRef]
- Schelhaas, M.-J.; Nabuurs, G.-J.; Schuck, A. Natural disturbances in the European forests in the 19th and 20th centuries. Glob. Change Biol. 2003, 9, 1620–1633. [Google Scholar] [CrossRef]
- Gardiner, B.; Schuck, A.; Schelhaas, M.-J.; Orazio, C.; Blennow, K.; Nicoll, B. What Science Can Tell Us. In Living with Storm Damage to Forests; European Forest Institute: Barcelona, Spain, 2013. [Google Scholar]
- Mayer, H.; Schindler, D. Forest meteorological fundamentals of storm damage in forests in connection with the extreme storm “Lothar”. Allg. Forst Jgdztg. 2002, 173, 200–208. [Google Scholar]
- Lindroth, A.; Lagergren, F.; Grelle, A.; Klemendtsson, L.; Langvall, O.; Weslien, P.; Tuulik, J. Storms can cause Europe-wide reduction in forest carbon sink. Glob. Chang. Biol. 2009, 15, 346–355. [Google Scholar] [CrossRef]
- Dahle, A.G.; James, K.R.; Kane, B.; Grabosky, J.C.; Detter, A. A review of factors that affect the static load-bearing capacity of urban trees. Arboricul. Urban. For. 2017, 43, 89–106. [Google Scholar]
- Gardiner, B.; Byrne, K.; Hale, S.; Kamimura, S.; Mitchell, S.J.; Peltola, H.; Ruel, J.-C. A review of mechanistic modelling of wind damage risk to forests. Forestry 2008, 81, 447–463. [Google Scholar] [CrossRef]
- Peltola, H.; Kellomäki, S.; Vaisanen, H.; Ikonen, V.-P. A mechanistic model for assessing the risk of wind and snow damage to single trees and stands of Scots pine, Norway spruce, and birch. Can. J. For. Res. 1999, 29, 647–661. [Google Scholar] [CrossRef]
- Gardiner, B.; Peltola, H.; Kellomäki, S. Comparison of two models for predicting the critical winds required to damage coniferous trees. Ecol. Model. 2000, 129, 1–23. [Google Scholar] [CrossRef]
- Ancelin, P.; Courbaud, B.; Fourcaud, T. Development of an individual tree-based mechanical model to predict wind damage within forest stands. For. Ecol. Manag. 2004, 203, 101–121. [Google Scholar] [CrossRef]
- Byrne, K.E.; Mitchell, S.J. Testing of WindFIRM/ForestGALES_BC: A hybrid-mechanistic model for predicting windthrow in partially harvested stands. Forestry 2013, 86, 185–199. [Google Scholar] [CrossRef]
- Dupont, S.; Ikonen, V.-P.; Väisänen, H.; Peltola, H. Predicting tree damage in fragmented landscapes using a wind risk model coupled with an airflow model. Can. J. For. Res. 2015, 45, 1065–1076. [Google Scholar] [CrossRef]
- Blennow, K.; Sallnäs, O. WINDA—A system of models for assessing the probability of wind damage to forest stands within a landscape. Ecol. Model. 2004, 175, 87–99. [Google Scholar] [CrossRef]
- Schelhaas, M.-J.; Kramer, K.; Peltola, H.; van der Werf, D.C.; Wijdeven, S.M.J. Introducing tree interactions in wind damage simulation. Ecol. Model. 2007, 207, 197–209. [Google Scholar] [CrossRef]
- Kamimura, K.; Gardiner, B.; Kato, A.; Hiroshima, T.; Shiraishi, N. Developing a decision support approach to reduce wind damage risk – a case study on sugi (Cryptomeria japonica (L.f.) D.Don) forests in Japan. Forestry 2008, 81, 429–445. [Google Scholar] [CrossRef]
- Seidl, R.; Rammer, W.; Blennow, K. Simulating wind disturbance impacts on forest landscapes: Tree-level heterogeneity matters. Environ. Modell. Softw. 2014, 51, 1–11. [Google Scholar] [CrossRef]
- Kamimura, K.; Gardiner, B.; Dupont, S.; Finnigan, J. Agent-based modelling of wind damage processes and patterns in forests. Agr. Forest Meteorol. 2019, 268, 279–288. [Google Scholar] [CrossRef]
- Moore, J. 2000: Differences in maximum resistive bending moments of Pinus radiata trees grown on a range of soil types. Forest Ecol. Manag. 2000, 135, 63–71. [Google Scholar] [CrossRef]
- Nicoll, B.C.; Gardiner, B.A.; Rayner, B.; Peace, A.J. Anchorage of coniferous trees in relation to species, soil type, and rooting depth. Can. J. For. Res. 2006, 36, 1871–1883. [Google Scholar] [CrossRef]
- Lundström, T.; Jonas, T.; Stöckli, V.; Ammann, W. Anchorage of mature conifers: Resistive turning moment, root–soil plate geometry and root growth orientation. Tree Physiol. 2007, 27, 1217–1227. [Google Scholar] [CrossRef]
- Kane, B.; Clouston, P. Tree pulling tests of large shade trees in the genus Acer. Arboricul. Urban. For. 2008, 34, 101–109. [Google Scholar]
- Rahardjo, H.; Harnas, F.R.; Indrawan, I.G.B.; Leong, E.C.; Tan, P.Y.; Fong, Y.K.; Ow, L.F. Understanding the stability of Samanea saman trees through tree pulling, analytical calculations and numerical models. Urban. For. Urban. Gree. 2014, 13, 355–364. [Google Scholar] [CrossRef]
- Peltola, H. Mechanical stability of trees under static loads. Am. J. Bot. 2006, 93, 1501–1511. [Google Scholar] [CrossRef] [PubMed]
- Hale, S.E.; Gardiner, B.A.; Wellpott, A.; Nicoll, B.C.; Alexis, A. Wind loading of trees: Influence of tree size and competition. Eur. J. Forest Res. 2012, 131, 203–217. [Google Scholar] [CrossRef]
- Sani, L.; Lisci, R.; Moschi, M.; Sarri, D.; Rimediotti, M.; Vieri, M.; Tofanelli, S. Preliminary experiments and verification of controlled pulling tests for tree stability assessments in Mediterranean urban areas. Biosyst. Eng. 2012, 112, 218–226. [Google Scholar] [CrossRef]
- Detter, A.; Rust, S. Findings of recent research on the pulling test method. In Jahrbuch der Baumpflege; Dujesiefken, D., Ed.; Haymarket Media: Twickenham, Germany, 2013; pp. 87–100. [Google Scholar]
- Gross, G. A windthrow model for urban trees with application to storm “Xavier”. Meteorl. Z. 2018, 27, 299–308. [Google Scholar] [CrossRef]
- Holbo, H.R.; Corbett, T.C.; Horton, P.J. Aeromechanical behavior of selected Douglas-fir. Agr. Meteorol. 1980, 21, 81–91. [Google Scholar] [CrossRef]
- Mayer, H. Wind-induced tree sways. Trees 1987, 1, 195–206. [Google Scholar] [CrossRef]
- Gardiner, B.A. The interactions of wind and tree movement in forest canopies. In Wind and Trees; Coutts, M.P., Grace, J., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 41–59. [Google Scholar]
- Peltola, H. Swaying of trees in response to wind and thinning in a stand of Scots pine. Bound.-Lay. Meteorol. 1996, 77, 285–304. [Google Scholar] [CrossRef]
- Schindler, D.; Vogt, R.; Fugmann, H.; Rodriguez, M.; Schönborn, J.; Mayer, H. Vibration behavior of plantation-grown Scots pine trees in response to wind excitation. Agr. Forest Meteorol. 2010, 150, 984–993. [Google Scholar] [CrossRef]
- Schindler, D.; Fugmann, H.; Mayer, H. Analysis and simulation of dynamic response behaviour of Scots pine trees to wind loading. Int. J. Biometeorol. 2013, 57, 819–833. [Google Scholar] [CrossRef]
- Schindler, D.; Mohr, M. Non-oscillatory response to wind loading dominates movement of Scots pine trees. Agr. Forest Meteorol. 2018, 250-251, 209–216. [Google Scholar] [CrossRef]
- Schindler, D.; Mohr, M. No resonant response of Scots pine trees to wind excitation. Agr. Forest Meteorol. 2019, 265, 227–244. [Google Scholar] [CrossRef]
- Schindler, D.; Schönborn, J.; Fugmann, H.; Mayer, H. Responses of an individual deciduous broadleaved tree to wind excitation. Agr. Forest Meteorol. 2013, 177, 69–82. [Google Scholar] [CrossRef]
- Angelou, N.; Dellwik, E.; Mann, J. Wind load estimation on an open-grown European oak tree. Forestry 2019, 92, 381–392. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.J.; Lexer, M.J. Unraveling the drivers of intensifying forest disturbance regimes in Europe. Glob. Change Biol. 2011, 17, 2842–2852. [Google Scholar] [CrossRef]
- Mölter, T.; Schindler, D.; Albrecht, A.; Kohnle, U. Review on the projections of future storminess over the North Atlantic European Region. Atmosphere 2016, 7, 60. [Google Scholar] [CrossRef]
- Blennow, K.; Andersson, M.; Bergh, J.; Sallnäs, O.; Olofsson, E. Potential climate change impacts on the probability of wind damage in a south Swedish forest. Clim. Change 2010, 99, 261–278. [Google Scholar] [CrossRef]
- Blennow, K.; Andersson, M.; Sallnäs, O.; Olofsson, E. Climate change and the probability of wind damage in two Swedish forests. For. Ecol. Manag. 2010, 259, 818–830. [Google Scholar] [CrossRef]
- Peltola, H.; Ikonen, V.-P.; Gregow, H.; Strandman, H.; Kilpäinen, A.; Venäläinen, A.; Kellomäki, S. Impacts of climate change on timber production and regional risks of wind-induced damage to forests in Finland. Forest Ecol. Manag. 2010, 260, 833–845. [Google Scholar] [CrossRef]
- Schelhaas, M.J.; Hengeveld, G.; Moriondo, M.; Reinds, G.J.; Kundzewicz, Z.W.; ter Maat, H.; Bindi, M. Assessing risk and adaptation options to fires and windstorms in European forestry. Mitig. Adapt. Strat. GL. 2010, 15, 681–701. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.-J.; Rammer, W.; Verkerk, P.J. Increasing forest disturbances in Europe and their impact on carbon storage. Nat. Clim. Change 2014, 4, 806–810. [Google Scholar] [CrossRef] [PubMed]
- Mohr, M.; Schindler, D. Coherent momentum exchange above and within a Scots pine forest. Atmosphere 2016, 7, 61. [Google Scholar] [CrossRef]
- Lavielle, M. Using penalized contrasts for the change-point problem. Signal. Process. 2005, 85, 1501–1510. [Google Scholar] [CrossRef]
- Aubry, N.; Guyonnet, R.; Lima, R. Spatiotemporal analysis of complex signals: Theory and applications. J. Stat. Phys. 1991, 64, 683–739. [Google Scholar] [CrossRef]
- Hémon, P.; Santi, F. Applications of biorthogonal decompositions in fluid structure interactions. J. Fluids Struct. 2003, 17, 1123–1143. [Google Scholar] [CrossRef]
- Kaiser, H.F. The application of electronic computers to factor analysis. Educ. Psychol. Meas. 1960, 20, 141–151. [Google Scholar] [CrossRef]
- Broomhead, D.; King, G. Extracting qualitative dynamics from experimental data. Physica D 1986, 20, 217–236. [Google Scholar] [CrossRef]
- Vautard, R.; Ghil, M. Singular spectrum analysis in nonlinear dynamics with applications to paleoclimatic time series. Physica D 1989, 35, 395–424. [Google Scholar] [CrossRef]
- Schindler, D. Responses of Scots pine trees to dynamic wind loading. Agr. For. Meteorol. 2008, 148, 1733–1742. [Google Scholar] [CrossRef]
- Sellier, D.; Brunet, Y.; Fourcaud, T. A numerical model of tree aerodynamic response to a turbulent airflow. Forestry 2008, 81, 279–297. [Google Scholar] [CrossRef]
- Rudnicki, M.; Mitchell, S.J.; Novak, M.D. Wind tunnel measurements of crown streamlining and drag relationships for three conifer species. Can. J. For. Res. 2004, 34, 666–676. [Google Scholar] [CrossRef]
- Rudnicki, M.; Meyer, T.H.; Lieffers, V.J.; Silins, U.; Webb, V.A. The periodic motion of lodgepole pine trees as affected by collisions with neighbors. Trees 2008, 22, 475–482. [Google Scholar] [CrossRef]
- James, K.R.; Dahle, G.A.; Grabosky, J.; Kane, B.; Detter, A. Tree biomechanics literature review: Dynamics. Arboricul. Urban. For. 2014, 40, 1–15. [Google Scholar]
- Spatz, H.-C.; Theckes, B. Oscillation damping in trees. Plant. Sci. 2013, 207, 66–71. [Google Scholar] [CrossRef]
- Miller, L.A. Structural dynamics and resonance in plants with nonlinear stiffness. J. Theor. Biol. 2005, 234, 511–534. [Google Scholar] [CrossRef]
- Sakoe, H.; Chiba, S. Dynamic programming algorithm optimization for spoken word recognition. IEEE Trans. Acoust. Speech 1978, 26, 43–48. [Google Scholar] [CrossRef]
- Paliwal, K.K.; Agarwal, A.; Sinha, S.S. A modification over Sakoe and Chiba’s dynamic time warping algorithm for isolated word recognition. Signal. Process. 1982, 4, 329–333. [Google Scholar] [CrossRef]
- Matlab. dtw—Distance between Signals Using Dynamic Time Warping. Available online: https://www.mathworks.com/help/signal/ref/dtw.html (accessed on 19 December 2019).
- Stacey, G.R.; Belcher, R.E.; Wood, C.J.; Gardiner, B.A. Wind flows and forces in a model spruce forest. Bound.-Lay. Meteorol. 1994, 69, 311–334. [Google Scholar] [CrossRef]
- Gardiner, B.A.; Stacey, G.R.; Belcher, R.E.; Wood, C.J. Field and wind tunnel assessments of the implications of respacing and thinning for tree stability. Forestry 1997, 70, 233–252. [Google Scholar] [CrossRef]
- Atanasov, D. Two-Phase Linear Regression Model. 2017. Available online: https://de.mathworks.com/matlabcentral/fileexchange/26804-two-phase-linear-regression-model?s_tid=prof_contriblnk (accessed on 19 December 2019).
- Dupont, S.; Défossez, P.; Bonnefond, J.M.; Irvine, M.R.; Garrigou, D. How stand tree motion impacts wind dynamics during windstorms. Agr. For. Meteorol. 2018, 262, 42–58. [Google Scholar] [CrossRef]
- Gardiner, B. Wind and wind forces in a plantation spruce forest. Bound.-Lay. Meteorol. 1994, 67, 161–184. [Google Scholar] [CrossRef]
- Jackson, T.; Shenkin, A.; Wellpott, A.; Calders, K.; Origo, N.; Disney, M.; Burt, A.; Raumonen, P.; Gardiner, B.; Herold, M.; et al. Finite element analysis of trees in the wind based on terrestrial laser scanning data. Agr. For. Meteorol. 2019, 265, 137–144. [Google Scholar] [CrossRef]











© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schindler, D.; Kolbe, S. Assessment of the Response of a Scots Pine Tree to Effective Wind Loading. Forests 2020, 11, 145. https://doi.org/10.3390/f11020145
Schindler D, Kolbe S. Assessment of the Response of a Scots Pine Tree to Effective Wind Loading. Forests. 2020; 11(2):145. https://doi.org/10.3390/f11020145
Chicago/Turabian StyleSchindler, Dirk, and Sven Kolbe. 2020. "Assessment of the Response of a Scots Pine Tree to Effective Wind Loading" Forests 11, no. 2: 145. https://doi.org/10.3390/f11020145
APA StyleSchindler, D., & Kolbe, S. (2020). Assessment of the Response of a Scots Pine Tree to Effective Wind Loading. Forests, 11(2), 145. https://doi.org/10.3390/f11020145