Contrasting Root System Structure and Belowground Interactions between Black Spruce (Picea mariana (Mill.) B.S.P) and Trembling Aspen (Populus tremuloides Michx) in Boreal Mixedwoods of Eastern Canada


Abstract
1. Introduction
2. Materials and Methods
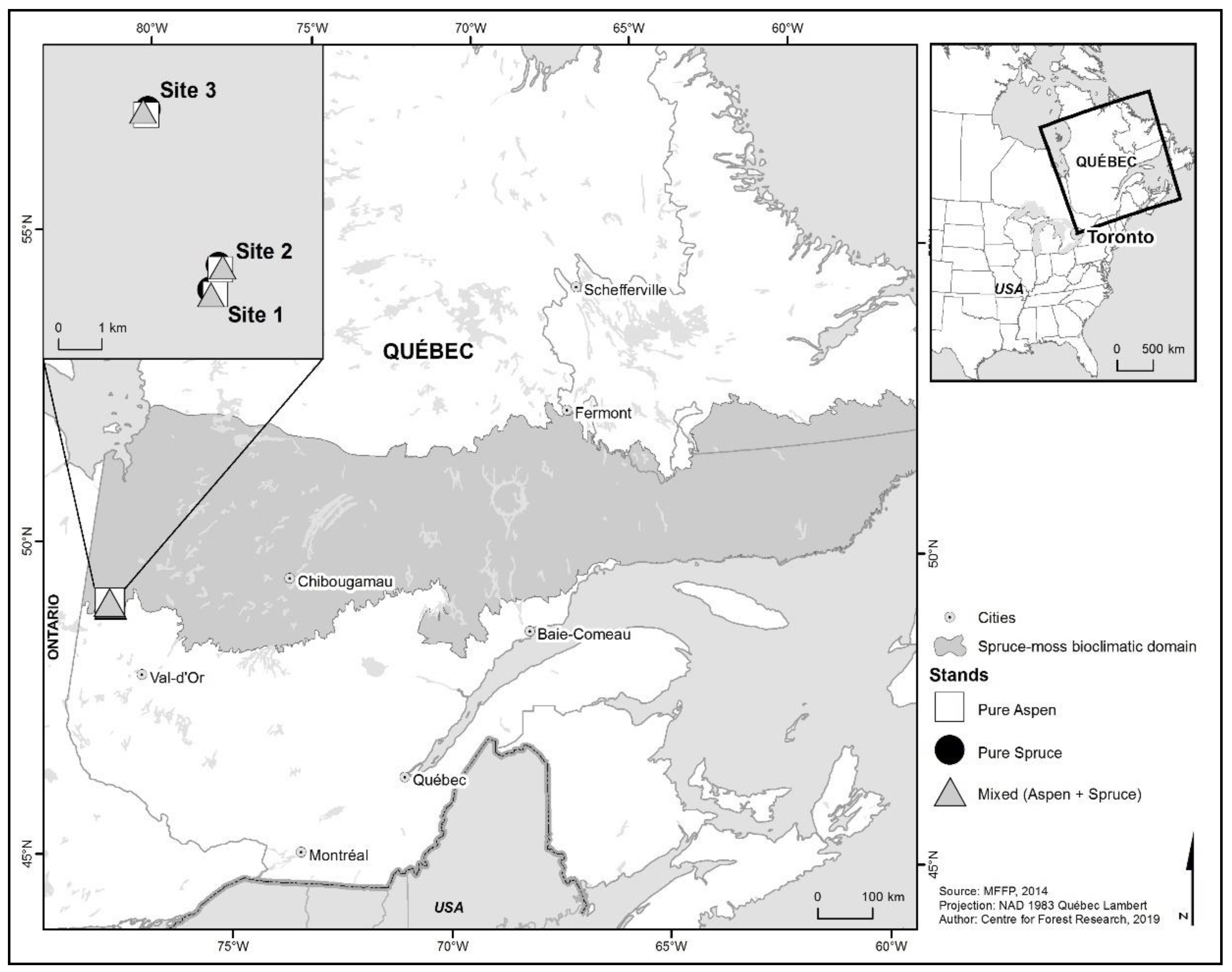
2.1. Study Area
2.2. Sample Collection and Preparation
2.3. Chemical Analysis
2.4. Data Analysis
2.4.1. Metrics Calculation
2.4.2. Statistical Analysis
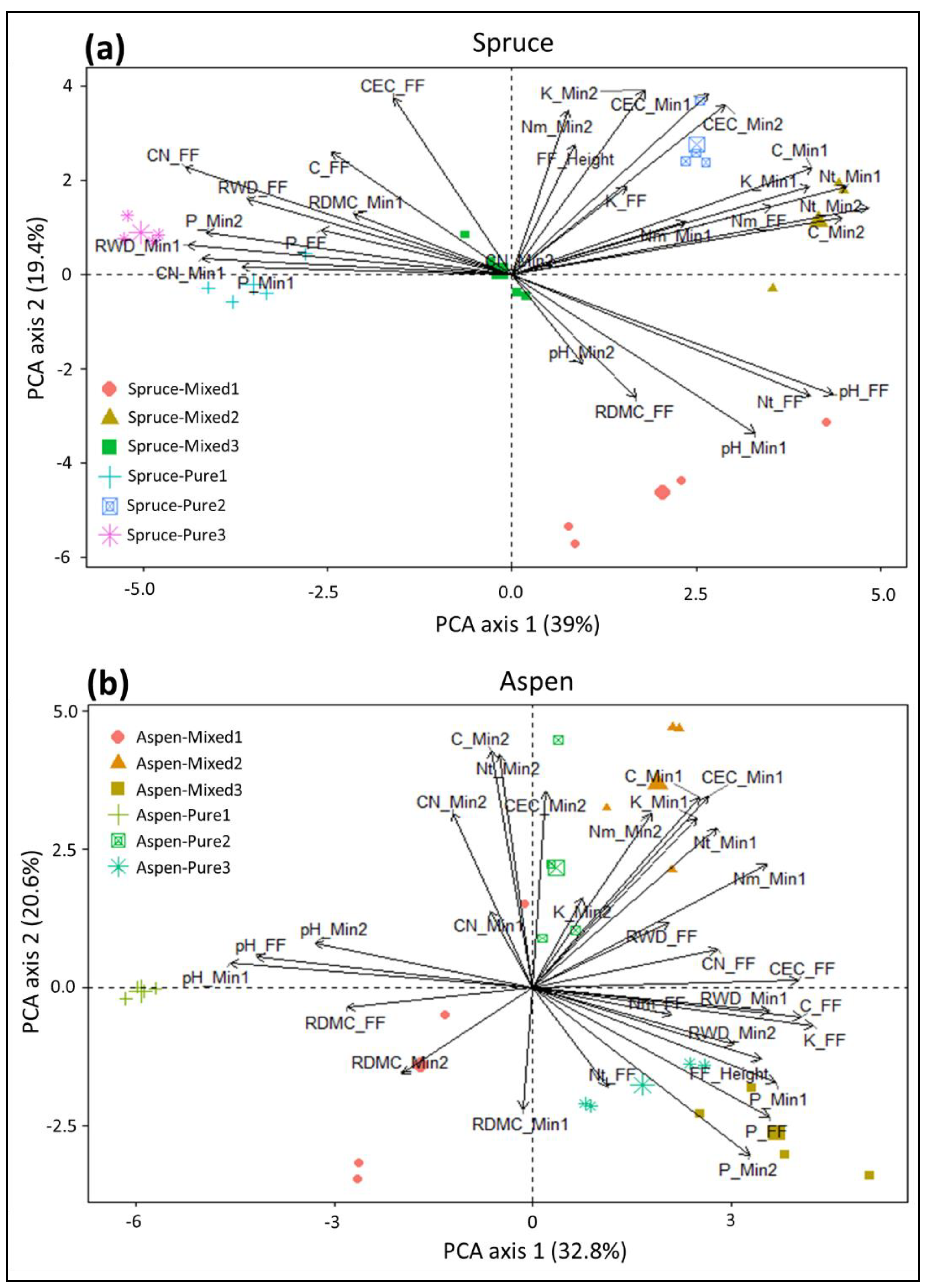
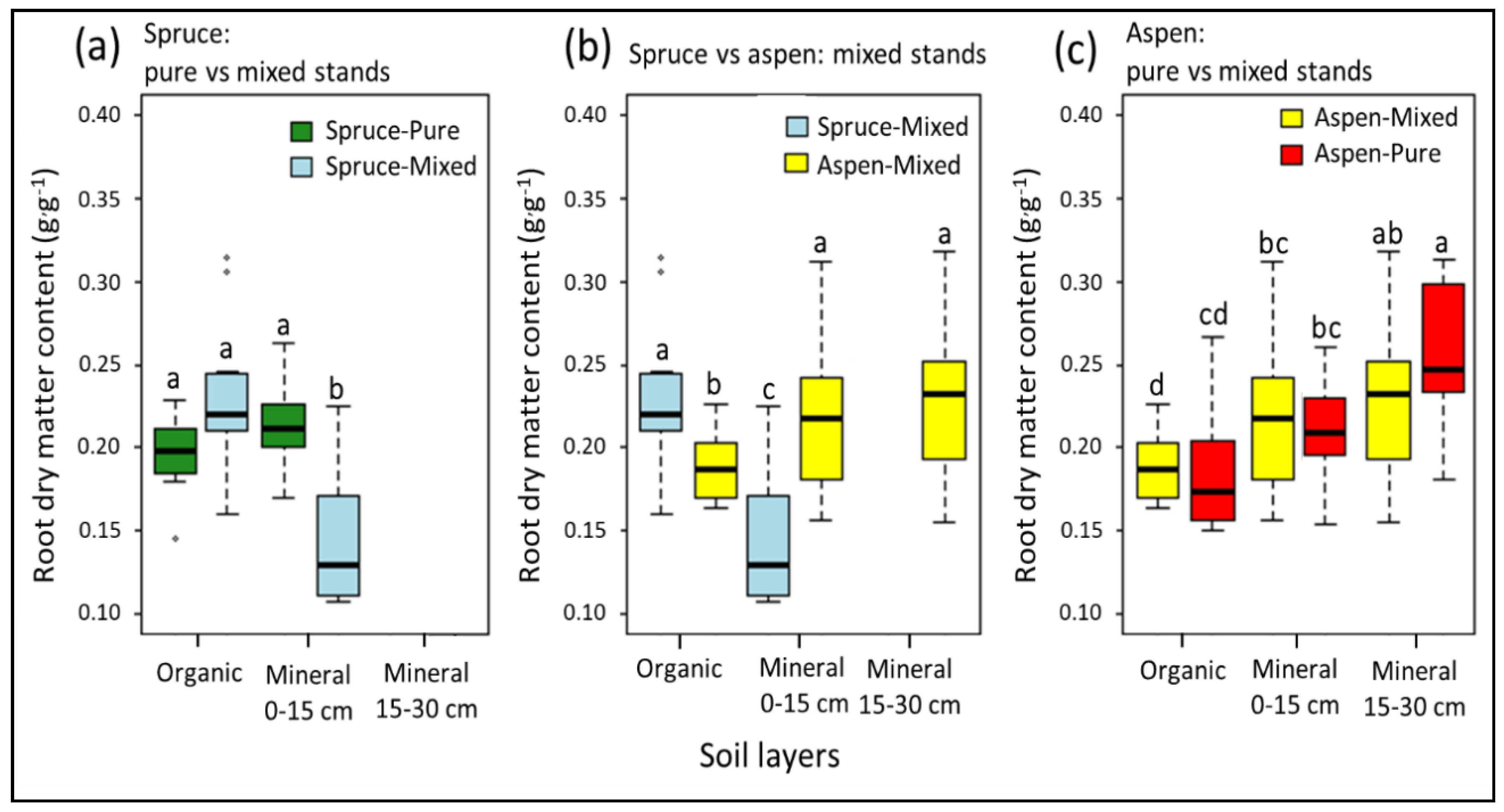
3. Results
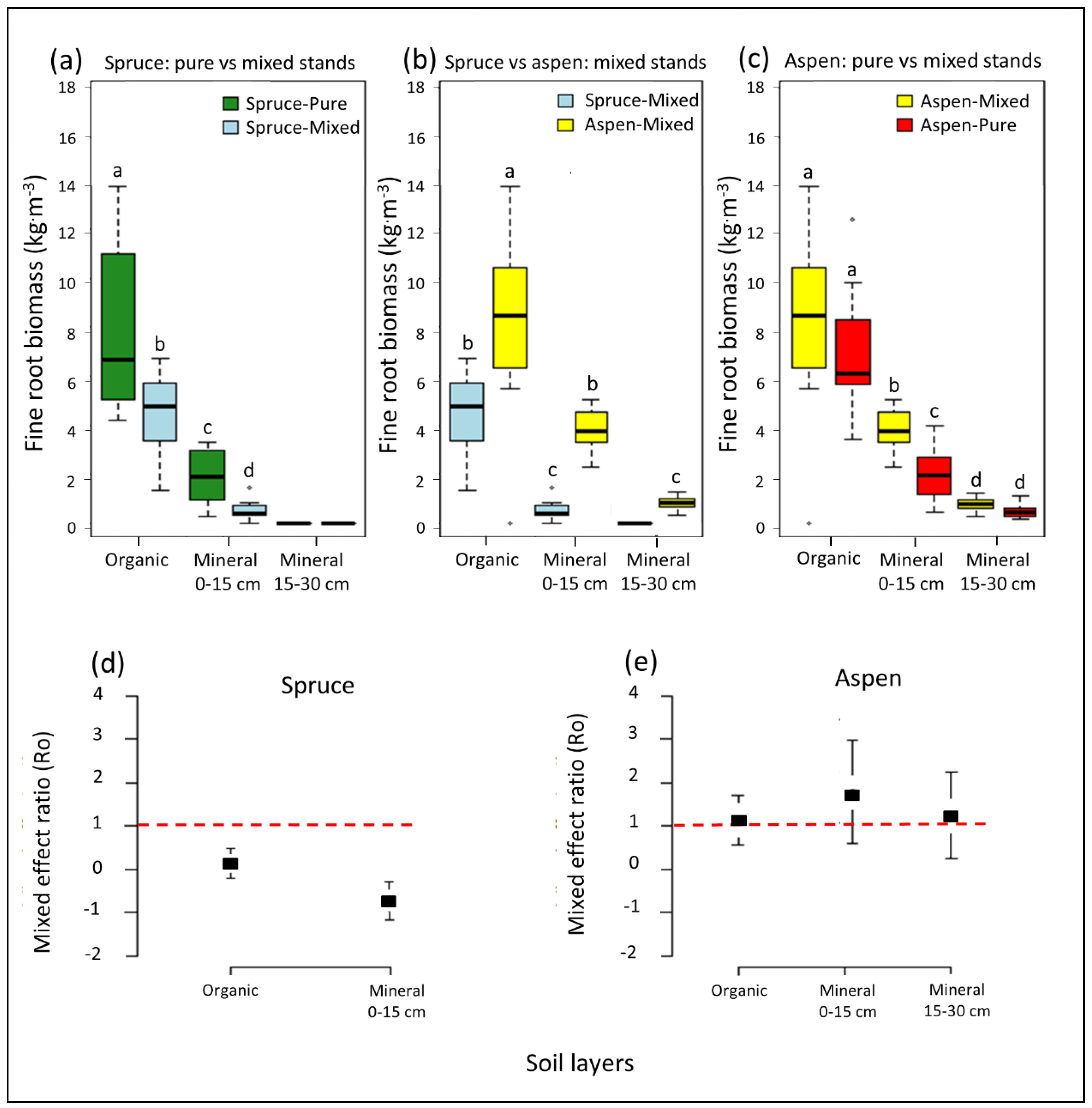
3.1. Fine Root Biomass
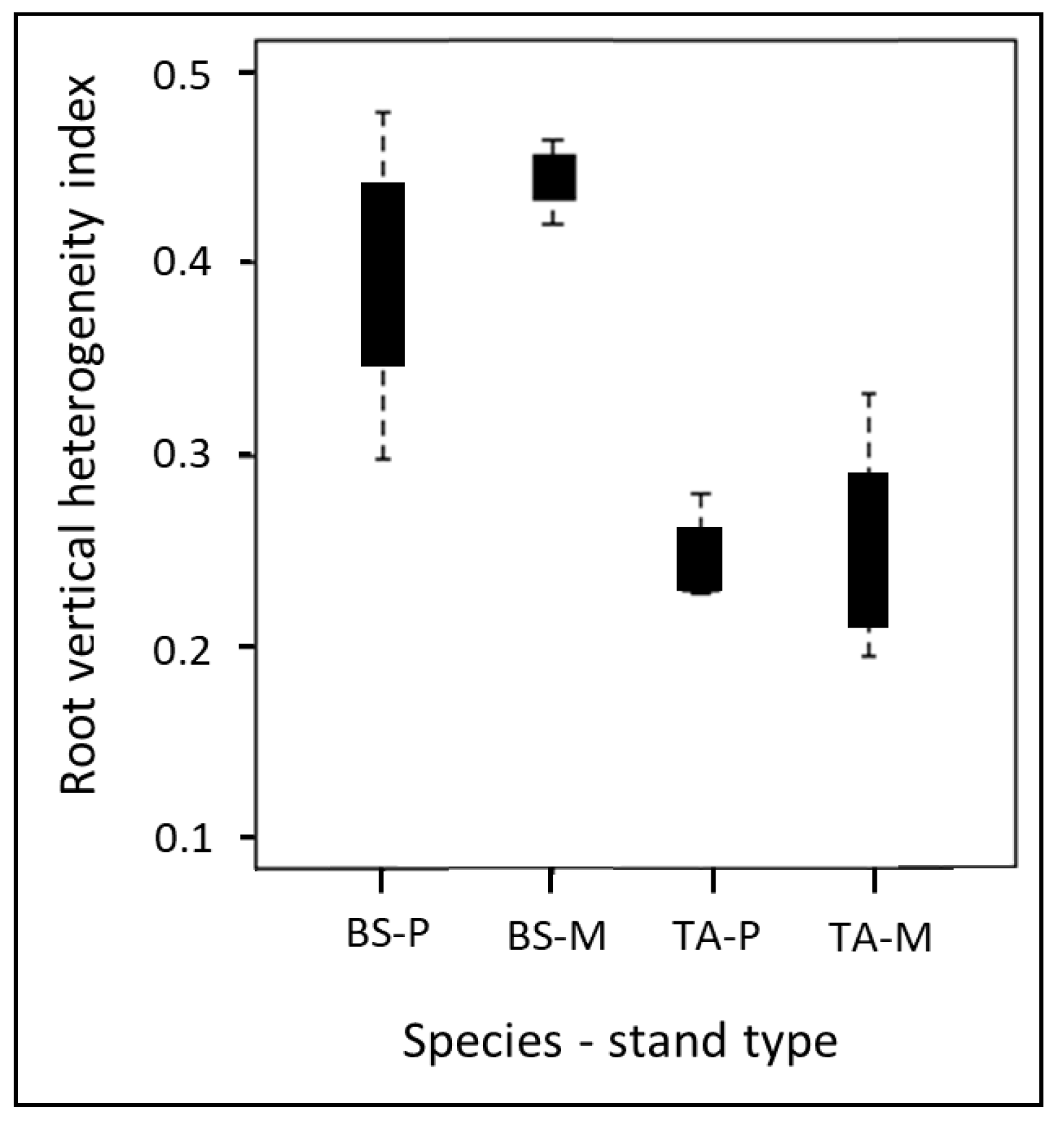
3.2. Root Dry Matter Content
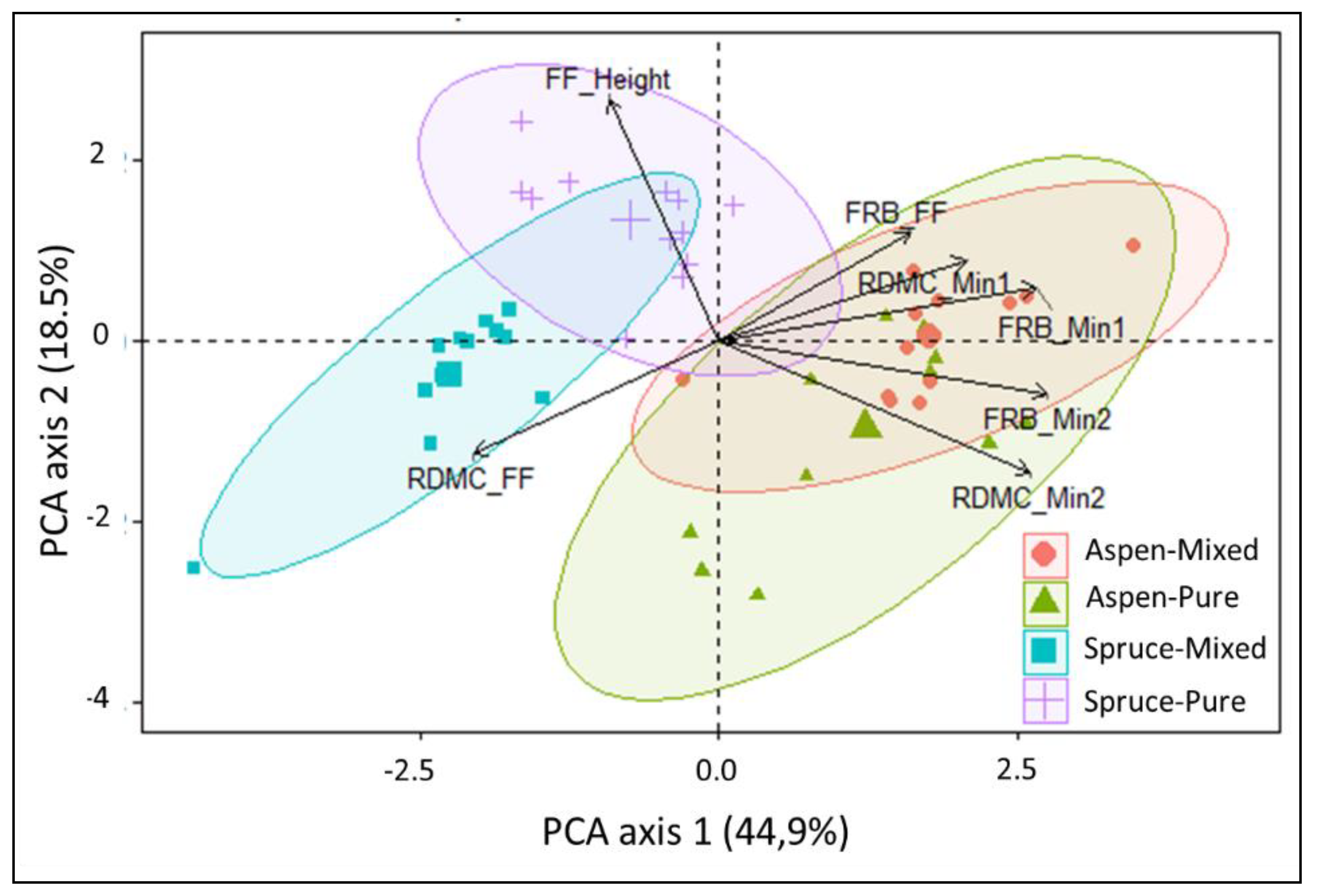
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liang, J.; Crowther, T.W.; Picard, N.; Wiser, S.; Zhou, M.; Alberti, G.; Schulze, E.D.; McGuire, A.D.; Bozzato, F.; Pretzsch, H.; et al. Positive biodiversity-productivity relationship predominant in global forests. Science 2016, 354, aaf8957. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.Y.H.; Taylor, A.R. Positive species diversity and above-ground biomass relationships are ubiquitous across forest strata despite interference from overstorey trees. Funct. Ecol. 2016, 31, 419–426. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.Y.H.; Reich, P.B. Forest productivity increases with evenness, species richness and trait variation: A global meta-analysis. J. Ecol. 2012, 100, 742–749. [Google Scholar] [CrossRef]
- Morin, X.; Fahse, L.; Scherer-Lorenzen, M.; Bugmann, H. Tree species richness promotes productivity in temperate forests through strong complementarity between species: Species richness promotes forest productivity. Ecol. Lett. 2011, 14, 1211–1219. [Google Scholar] [CrossRef] [PubMed]
- Ratcliffe, S.; Holzwarth, F.; Nadrowski, K.; Levick, S.; Wirth, C. Tree neighbourhood matters—Tree species composition drives diversity–productivity patterns in a near-natural beech forest. For. Ecol. Manag. 2015, 335, 225–234. [Google Scholar] [CrossRef]
- Roscher, C.; Schumacher, J.; Gubsch, M.; Lipowsky, A.; Weigelt, A.; Buchmann, N.; Schmid, B.; Schulze, E.-D. Using Plant Functional Traits to Explain Diversity–Productivity Relationships. PLoS ONE 2012, 7, e36760. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Benito, P.; Gómez-Aparicio, L.; Paquette, A.; Messier, C.; Kattge, J.; Zavala, M.A. Diversity increases carbon storage and tree productivity in Spanish forests: Diversity effects on forest carbon storage and productivity. Glob. Ecol. Biogeogr. 2014, 23, 311–322. [Google Scholar] [CrossRef]
- Franklin, J.F.; Van Pelt, R. Spatial aspects of structural complexity in old-growth forests. J. For. 2004, 102, 22–28. [Google Scholar]
- Hunter, M.L. Maintaining Biodiversity in Forest Ecosystems, 1st ed.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 1999; ISBN 0-521-63104-1. [Google Scholar]
- Légaré, S.; Paré, D.; Bergeron, Y. Influence of Aspen on Forest Floor Properties in Black Spruce-dominated Stands. Plant Soil 2005, 275, 207–220. [Google Scholar] [CrossRef]
- Lei, X.; Wang, W.; Peng, C. Relationships between stand growth and structural diversity in spruce-dominated forests in New Brunswick, Canada. Can. J. For. Res. 2009, 39, 1835–1847. [Google Scholar] [CrossRef]
- Paquette, A.; Messier, C. The effect of biodiversity on tree productivity: From temperate to boreal forests. Glob. Ecol. Biogeogr. 2011, 20, 170–180. [Google Scholar] [CrossRef]
- Wang, W.; Lei, X.; Ma, Z.; Kneeshaw, D.D.; Peng, C. Positive relationship between aboveground carbon stocks and structural diversity in spruce-dominated forest stands in New Brunswick, Canada. For. Sci. 2011, 57, 506–515. [Google Scholar]
- Aussenac, R.; Bergeron, Y.; Ghotsa Mekontchou, C.; Gravel, D.; Pilch, K.; Drobyshev, I. Intraspecific variability in growth response to environmental fluctuations modulates the stabilizing effect of species diversity on forest growth. J. Ecol. 2016, 105, 1010–1020. [Google Scholar] [CrossRef]
- Morin, X.; Fahse, L.; de Mazancourt, C.; Scherer-Lorenzen, M.; Bugmann, H. Temporal stability in forest productivity increases with tree diversity due to asynchrony in species dynamics. Ecol. Lett. 2014, 17, 1526–1535. [Google Scholar] [CrossRef] [PubMed]
- Yachi, S.; Loreau, M. Biodiversity and ecosystem productivity in a fluctuating environment: The insurance hypothesis. Proc. Natl. Acad. Sci. USA 1999, 96, 1463–1468. [Google Scholar] [CrossRef] [PubMed]
- Cavard, X.; Bergeron, Y.; Chen, H.Y.H.; Paré, D. Mixed-species effect on tree aboveground carbon pools in the east-central boreal forests. Can. J. For. Res. 2010, 40, 37–47. [Google Scholar] [CrossRef]
- Chen, H.Y.; Klinka, K.; Mathey, A.H.; Wang, X.; Varga, P.; Chourmouzis, C. Are mixed-species stands more productive than single-species stands: An empirical test of three forest types in British Columbia and Alberta. Can. J. For. Res. 2003, 33, 1227–1237. [Google Scholar] [CrossRef]
- Drobyshev, I.; Gewehr, S.; Berninger, F.; Bergeron, Y. Species specific growth responses of black spruce and trembling aspen may enhance resilience of boreal forest to climate change. J. Ecol. 2013, 101, 231–242. [Google Scholar] [CrossRef]
- Cavard, X.; Bergeron, Y.; Chen, H.Y.H.; Paré, D.; Laganière, J.; Brassard, B. Competition and facilitation between tree species change with stand development. Oikos 2011, 120, 1683–1695. [Google Scholar] [CrossRef]
- Filipescu, C.N.; Comeau, P.G. Competitive interactions between aspen and white spruce vary with stand age in boreal mixedwoods. For. Ecol. Manag. 2007, 247, 175–184. [Google Scholar] [CrossRef]
- Armstrong, G.W. Considerations for boreal mixedwood silviculture: A view from the dismal science. For. Chron. 2014, 90, 44–49. [Google Scholar] [CrossRef]
- Zhang, D.; Pearse, P.H. Forest Economics; UBC Press: Vancouver, BC, Canada, 2012; ISBN 978-0-7748-2153-7. [Google Scholar]
- Bergeron, Y.; Chen, H.Y.; Kenkel, N.C.; Leduc, A.L.; Macdonald, S.E. Boreal mixedwood stand dynamics: Ecological processes underlying multiple pathways. For. Chron. 2014, 90, 202–213. [Google Scholar] [CrossRef]
- Légaré, S.; Paré, D.; Bergeron, Y. The responses of black spruce growth to an increased proportion of aspen in mixed stands. Can. J. For. Res. 2004, 34, 405–416. [Google Scholar] [CrossRef]
- Statistics Canada. Human Activity and the Environment 2017. Can. Public Policy Anal. Polit. 2018, 4, 587. [Google Scholar] [CrossRef]
- Wirth, C. Fire Regime and Tree Diversity in Boreal Forests: Implications for the Carbon Cycle. In Forest Diversity and Function; Scherer-Lorenzen, M., Körner, C., Schulze, E.D., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; Volume 176, pp. 309–344. ISBN 3-540-22191-3. [Google Scholar]
- Bergeron, Y.; Leduc, A.; Joyal, C.; Morin, H. Balsam fir mortality following the last spruce budworm outbreak in northwestern Quebec. Can. J. For. Res. 1995, 25, 1375–1384. [Google Scholar] [CrossRef]
- Bouchard, M.; Kneeshaw, D.; Bergeron, Y. Forest dynamics after successive spruce budworm outbreaks in mixedwood forests. Ecology 2006, 87, 2319–2329. [Google Scholar] [CrossRef]
- McKenney, D.W.; Pedlar, J.H.; Lawrence, K.; Campbell, K.; Hutchinson, M.F. Potential Impacts of Climate Change on the Distribution of North American Trees. BioScience 2007, 57, 939–948. [Google Scholar] [CrossRef]
- McKenney, D.W.; Pedlar, J.H.; Rood, R.B.; Price, D. Revisiting projected shifts in the climate envelopes of North American trees using updated general circulation models. Glob. Chang. Biol. 2011, 17, 2720–2730. [Google Scholar] [CrossRef]
- Callaway, R.M. The detection of neighbors by plants. Trends Ecol. Evol. 2002, 17, 104–105. [Google Scholar] [CrossRef]
- Kuebbing, S.E.; Nuñez, M.A. Negative, neutral, and positive interactions among nonnative plants: Patterns, processes, and management implications. Glob. Chang. Biol. 2015, 21, 926–934. [Google Scholar] [CrossRef]
- Callaway, R.M. Positive interactions among plants. Bot. Rev. 1995, 61, 306–349. [Google Scholar] [CrossRef]
- Callaway, R.M.; Brooker, R.W.; Choler, P.; Kikvidze, Z.; Lortie, C.J.; Michalet, R.; Paolini, L.; Pugnaire, F.I.; Newingham, B.; Aschehoug, E.T.; et al. Positive interactions among alpine plants increase with stress. Nature 2002, 417, 844–848. [Google Scholar] [CrossRef] [PubMed]
- De Kroon, H.; Hendriks, M.; van Ruijven, J.; Ravenek, J.; Padilla, F.M.; Jongejans, E.; Visser, E.J.W.; Mommer, L. Root responses to nutrients and soil biota: Drivers of species coexistence and ecosystem productivity. J. Ecol. 2012, 100, 6–15. [Google Scholar] [CrossRef]
- De Kroon, H.; Mommer, L.; Nishiwaki, A. Root Competition: Towards a Mechanistic Understanding. In Root Ecology; de Kroon, H., Visser, E.J.W., Eds.; Springer: Berlin/Heidelberg, Germany, 2003; Volume 168, pp. 215–234. ISBN 978-3-642-05520-1. [Google Scholar] [CrossRef]
- Goldberg, D.E. Components of resource competition in plant communities. Perspect. Plant Compet. 1990, 27–49. [Google Scholar]
- Holzapfel, C.; Mahall, B.E. Bidirectional facilitation and interference between shrubs and annuals in the Mojave Desert. Ecology 1999, 80, 1747–1761. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; DeForest, J.L.; Burton, A.J.; Allen, M.F.; Ruess, R.W.; Hendrick, R.L. Fine root architecture of nine North American trees. Ecol. Monogr. 2002, 72, 293–309. [Google Scholar] [CrossRef]
- Brassard, B.W.; Chen, H.Y.H.; Bergeron, Y. Influence of Environmental Variability on Root Dynamics in Northern Forests. Crit. Rev. Plant Sci. 2009, 28, 179–197. [Google Scholar] [CrossRef]
- Craine, J.M. Reconciling plant strategy theories of Grime and Tilman. J. Ecol. 2005, 93, 1041–1052. [Google Scholar] [CrossRef]
- Lõhmus, K.; Truu, J.; Truu, M.; Kaar, E.; Ostonen, I.; Alama, S.; Kuznetsova, T.; Rosenvald, K.; Vares, A.; Uri, V.; et al. Black alder as a promising deciduous species for the reclaiming of oil shale mining areas. In Proceedings of the Brownfields III: Prevention, Assessment, Rehabilitation and Development of Brownfield Sites, Tallin, Estonia, 2006; WIT Press: Southampton, UK, 2006; Volume 94, pp. 87–98. [Google Scholar] [CrossRef]
- Ostonen, I.; Lohmus, K.; Helmisaari, H.S.; Truu, J.; Meel, S. Fine root morphological adaptations in Scots pine, Norway spruce and silver birch along a latitudinal gradient in boreal forests. Tree Physiol. 2007, 27, 1627–1634. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; George, T.S.; Dupuy, L.X.; Karley, A.J.; Valentine, T.A.; Wiesel, L.; Wishart, J. Root traits for infertile soils. Front. Plant Sci. 2013, 4. [Google Scholar] [CrossRef]
- Ostonen, I.; Püttsepp, Ü.; Biel, C.; Alberton, O.; Bakker, M.R.; Lõhmus, K.; Majdi, H.; Metcalfe, D.; Olsthoorn, A.F.M.; Pronk, A.; et al. Specific root length as an indicator of environmental change. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2007, 141, 426–442. [Google Scholar] [CrossRef]
- Brassard, B.W.; Chen, H.Y.H.; Cavard, X.; Laganière, J.; Reich, P.B.; Bergeron, Y.; Paré, D.; Yuan, Z. Tree species diversity increases fine root productivity through increased soil volume filling. J. Ecol. 2013, 101, 210–219. [Google Scholar] [CrossRef]
- Tilman, D. Resource competition and plant traits: A response to Craine et al. 2005. J. Ecol. 2007, 95, 231–234. [Google Scholar] [CrossRef]
- Tilman, D. Resource Competition and Community Structure; Monographs in population biology; Princeton University Press: Princeton, NJ, USA, 1982; ISBN 0-691-08301-0. [Google Scholar]
- Grime, J.P. Evidence for the Existence of Three Primary Strategies in Plants and Its Relevance to Ecological and Evolutionary Theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Turner, B.L. Resource partitioning for soil phosphorus: A hypothesis. J. Ecol. 2008, 96, 698–702. [Google Scholar] [CrossRef]
- Iwasa, Y.; Roughgarden, J. Shoot/root balance of plants: Optimal growth of a system with many vegetative organs. Theor. Popul. Biol. 1984, 25, 78–105. [Google Scholar] [CrossRef]
- Grime, J.P.; Crick, J.; Rincon, J. The ecological significance of plasticity. Symp. Soc. Exp. Biol. 1986, 40, 5–29. [Google Scholar]
- Lõhmus, K.; Oja, T.; Lasn, R. Specific root area: A soil characteristic. Plant Soil 1989, 119, 245–249. [Google Scholar] [CrossRef]
- Addo-Danso, S.D.; Prescott, C.E.; Adu-Bredu, S.; Duah-Gyamfi, A.; Moore, S.; Guy, R.D.; Forrester, D.I.; Owusu-Afriyie, K.; Marshall, P.L.; Malhi, Y. Fine-root exploitation strategies differ in tropical old growth and logged-over forests in Ghana. Biotropica 2018, 50, 606–615. [Google Scholar] [CrossRef]
- Veillette, J.J. Evolution and paleohydrology of glacial Lakes Barlow and Ojibway. Quat. Sci. Rev. 1994, 13, 945–971. [Google Scholar] [CrossRef]
- Environment Canada. Canadian Climate Normals 1971–2000. Available online: http://climate.wetheroffice.gc.ca/climate_normals/index_e.html (accessed on 12 January 2019).
- Brais, S.; Camiré, C. Keys for soil moisture regime evaluation for northwestern Quebec. Can. J. For. Res. 1992, 22, 718–724. [Google Scholar] [CrossRef]
- Ma, Z.; Chen, H.Y.H. Effects of species diversity on fine root productivity increase with stand development and associated mechanisms in a boreal forest. J. Ecol. 2017, 105, 237–245. [Google Scholar] [CrossRef]
- Ministère des Forêts de la Faune et des Parcs (MFFP). Norme de Stratification Ecoforestière du Quatrième Inventaire Ecoforestier du Québec Méridional. 2015, 111. Available online: http://collections.banq.qc.ca/ark:/52327/2748275 (accessed on 11 June 2019).
- Pacé, M.; Fenton, N.J.; Paré, D.; Bergeron, Y. Ground-layer composition affects tree fine root biomass and soil nutrient availability in jack pine and black spruce forests under extreme drainage conditions. Can. J. For. Res. 2017, 47, 433–444. [Google Scholar] [CrossRef]
- Houle, D.; Moore, J.D.; Ouimet, R.; Marty, C. Tree species partition N uptake by soil depth in boreal forests. Ecology 2014, 95, 1127–1133. [Google Scholar] [CrossRef]
- Birouste, M.; Zamora-Ledezma, E.; Bossard, C.; Pérez-Ramos, I.M.; Roumet, C. Measurement of fine root tissue density: A comparison of three methods reveals the potential of root dry matter content. Plant Soil 2014, 374, 299–313. [Google Scholar] [CrossRef]
- Carter, M.R.; Gregorich, E.G. Soil Sampling and Methods of Analysis, 2nd ed.; Canadian Society of Soil Science; CRC Press: Pinawa, MB, Canada; Boca Raton, FL, USA, 2008; ISBN 978-0-8493-3586-0. [Google Scholar]
- Mehlich, A. Mehlich 3 soil test extractant: A modification of Mehlich 2 extractant. Commun. Soil Sci. Plant Anal. 1984, 15, 1409–1416. [Google Scholar] [CrossRef]
- Brassard, B.W.; Chen, H.Y.H.; Bergeron, Y.; Paré, D. Differences in fine root productivity between mixed- and single-species stands: Fine root productivity in boreal forest. Funct. Ecol. 2011, 25, 238–246. [Google Scholar] [CrossRef]
- Loreau, M.; Hector, A. Partitioning selection and complementarity in biodiversity experiments. Nature 2001, 412, 72–76. [Google Scholar] [CrossRef]
- Zobel, M.; Zobel, K. Studying plant competition: From root biomass to general aims. Studying plant competition. J. Ecol. 2002, 90, 578–580. [Google Scholar] [CrossRef]
- Pierret, A.; Maeght, J.L.; Clément, C.; Montoroi, J.P.; Hartmann, C.; Gonkhamdee, S. Understanding deep roots and their functions in ecosystems: An advocacy for more unconventional research. Ann. Bot. 2016, 118, 621–635. [Google Scholar] [CrossRef]
- Roumet, C.; Birouste, M.; Picon-Cochard, C.; Ghestem, M.; Osman, N.; Vrignon-Brenas, S.; Cao, K.; Stokes, A. Root structure-function relationships in 74 species: Evidence of a root economics spectrum related to carbon economy. New Phytol. 2016, 210, 815–826. [Google Scholar] [CrossRef] [PubMed]
- McDonald, J.H. Handbook of Biological Statistics, 3rd ed.; Sparky House Publishing: Baltimore, MD, USA, 2014; Volume 2014. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Steele, S.J.; Gower, S.T.; Vogel, J.G.; Norman, J.M. Root mass, net primary production and turnover in aspen, jack pine and black spruce forests in Saskatchewan and Manitoba, Canada. Tree Physiol. 1997, 17, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Tamm, C.O. Nitrogen in Terrestrial Ecosystems: Questions of Productivity, Vegetational Changes, and Ecosystem Stability; Springer: Berlin/Heidelberg, Germany, 1991; ISBN 978-3-642-75168-4. [Google Scholar]
- Zackrisson, O.; Nilsson, M.-C.; Dahlberg, A.; Jäderlund, A.; Jaderlund, A. Interference Mechanisms in Conifer-Ericaceae-Feathermoss Communities. Oikos 1997, 78, 209. [Google Scholar] [CrossRef]
- Schenk, H.J. Root competition: Beyond resource depletion. Root competition: Beyond resource depletion. J. Ecol. 2006, 94, 725–739. [Google Scholar] [CrossRef]
- Pellegrini, A.F.A.; Ahlström, A.; Hobbie, S.E.; Reich, P.B.; Nieradzik, L.P.; Staver, A.C.; Scharenbroch, B.C.; Jumpponen, A.; Anderegg, W.R.L.; Randerson, J.T.; et al. Fire frequency drives decadal changes in soil carbon and nitrogen and ecosystem productivity. Nature 2017, 553, 194–198. [Google Scholar] [CrossRef]
- Cavard, X.; Macdonald, S.E.; Bergeron, Y.; Chen, H.Y.H. Importance of mixedwoods for biodiversity conservation: Evidence for understory plants, songbirds, soil fauna, and ectomycorrhizae in northern forests. Environ. Rev. 2011, 19, 142–161. [Google Scholar] [CrossRef]
- Légaré, S.; Bergeron, Y.; Leduc, A.; Paré, D. Comparison of the understory vegetation in boreal forest types of southwest Quebec. Can. J. Bot. 2001, 79, 1019–1027. [Google Scholar] [CrossRef]
- McClaugherty, C.A.; Aber, J.D.; Melillo, J.M. The Role of Fine Roots in the Organic Matter and Nitrogen Budgets of Two Forested Ecosystems. Ecology 1982, 63, 1481–1490. [Google Scholar] [CrossRef]
- Vogt, K.A.; Vogt, D.J.; Bloomfield, J. Input of Organic Matter to the Soil by Tree Roots. In Developments in Agricultural and Managed Forest Ecology; Elsevier: Amsterdam, The Netherlands, 1991; Volume 24, pp. 171–190. ISBN 978-0-444-89104-4. [Google Scholar] [CrossRef]
- Morrow, R.R. Periodicity and growth of sugar maple surface layer roots. J. For. 1950, 48, 875–881. [Google Scholar]
- Paula, S.; Pausas, J.G. Root traits explain different foraging strategies between resprouting life histories. Oecologia 2011, 165, 321–331. [Google Scholar] [CrossRef][Green Version]
- Bradshaw, A.D. Evolutionary Significance of Phenotypic Plasticity in Plants. In Advances in Genetics; Caspari, E.W., Thoday, J.M., Eds.; Academic Press: New York, NY, USA, 1965; Volume 13, pp. 115–155. ISBN 978-0-12-017613-7. [Google Scholar] [CrossRef]
- Bauhus, J.; Messier, C. Soil exploitation strategies of fine roots in different tree species of the southern boreal forest of eastern Canada. Can. J. For. Res. 1999, 29, 260–273. [Google Scholar] [CrossRef]
- Eissenstat, D.M.; Kucharski, J.M.; Zadworny, M.; Adams, T.S.; Koide, R.T. Linking root traits to nutrient foraging in arbuscular mycorrhizal trees in a temperate forest. New Phytol. 2015, 208, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Schmid, I. The influence of soil type and interspecific competition on the fine root system of Norway spruce and European beech. Basic Appl. Ecol. 2002, 3, 339–346. [Google Scholar] [CrossRef]
- Peltzer, D.A.; Wilson, S.D.; Gerry, A.K. Competition Intensity along a Productivity Gradient in a Low-Diversity Grassland. Am. Nat. 1998, 151, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.D.; Grime, J.P.; Mackey, J.M.L. A trade-off between scale and precision in resource foraging. Oecologia 1991, 87, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Grams, T.E.E.; Andersen, C.P. Competition for Resources in Trees: Physiological Versus Morphological Plasticity. In Progress in Botany; Esser, K., Löttge, U., Beyschlag, W., Murata, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; Volume 68, pp. 356–381. ISBN 978-3-540-36830-4. [Google Scholar] [CrossRef]
- Hutchings, M.J.; Wijesinghe, D.K. Patchy habitats, division of labour and growth dividends in clonal plants. Trends Ecol. Evol. 1997, 12, 390–394. [Google Scholar] [CrossRef]
- Stuefer, J.F. Two types of division of labour in clonal plants: Benefits, costs and constraints. Perspect. Plant Ecol. Evol. Syst. 1998, 1, 47–60. [Google Scholar] [CrossRef]
- Stuefer, J.F.; de Kroon, H.; During, H.J. Exploitation of environmental Hetergeneity by Spatial Division of Labor in a Clonal Plant. Funct. Ecol. 1996, 10, 328. [Google Scholar] [CrossRef]
- Hutchings, M.J.; John, E.A.; Wijesinghe, D.K. Toward understanding the consequences of soil heterogeneity for plant populations and communities. Ecology 2003, 84, 2322–2334. [Google Scholar] [CrossRef]
- Robertson, S.J.; Tackaberry, L.E.; Egger, K.N.; Massicotte, H.B. Ectomycorrhizal fungal communities of black spruce differ between wetland and upland forests. Can. J. For. Res. 2006, 36, 972–985. [Google Scholar] [CrossRef]
- Neville, J.; Tessier, J.L.; Morrison, I.; Scarratt, J.; Canning, B.; Klironomos, J.N. Soil depth distribution of ecto- and arbuscular mycorrhizal fungi associated with Populus tremuloides within a 3-year-old boreal forest clear-cut. Appl. Soil Ecol. 2002, 19, 209–216. [Google Scholar] [CrossRef]
- Kalliokoski, T.; Pennanen, T.; Nygren, P.; Sievänen, R.; Helmisaari, H.S. Belowground interspecific competition in mixed boreal forests: Fine root and ectomycorrhiza characteristics along stand developmental stage and soil fertility gradients. Plant Soil 2010, 330, 73–89. [Google Scholar] [CrossRef]
- Berger, T.W.; Sun, B.; Glatzel, G. Soil seed banks of pure spruce (Picea abies) and adjacent mixed species stands. Plant Soil 2004, 264, 53–67. [Google Scholar] [CrossRef]
- Berger, T.W.; Untersteiner, H.; Toplitzer, M.; Neubauer, C. Nutrient fluxes in pure and mixed stands of spruce (Picea abies) and beech (Fagus sylvatica). Plant Soil 2009, 322, 317–342. [Google Scholar] [CrossRef]
- Bhatti, J.S.; Foster, N.W.; Hazlett, P.W. Fine root biomass and nutrient content in a black spruce peat soil with and without alder. Can. J. Soil Sci. 1998, 78, 163–169. [Google Scholar] [CrossRef]
- Rosengren, U.; Göransson, H.; Jönsson, U.; Stjernquist, I.; Thelin, G.; Wallander, H. Functional Biodiversity Aspects on the Nutrient Sustainability in Forests-Importance of Root Distribution. J. Sustain. For. 2006, 21, 77–100. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site 1 | Site2 | Site 3 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| BS | TA | M | BS | TA | M | BS | TA | M | ||
| Species proportion (% number of stems) | Populus tremuloides Michx. | 2 | 60 | 34 | 2 | 70 | 38 | 2 | 74 | 43 |
| Picea mariana Mill. | 83 | 33 | 50 | 89 | 22 | 51 | 97 | 20 | 54 | |
| Abies balsamea (L.) Mill. | 6 | 3 | 2 | 6 | 1 | |||||
| Pinus banksiana Lamb. | 15 | 1 | 10 | 9 | 5 | 11 | 1 | 1 | ||
| Betula papyrifera Marshall | 1 | 1 | ||||||||
| Populus balsamifera L. | 2 | 1 | ||||||||
| Density (stems ha−1) | 3100 | 1410 | 1160 | 3000 | 1220 | 1610 | 2382 | 900 | 1680 | |
| Organic soil layer properties | pH | 3.8 | 4.0 | 4.9 | 4.2 | 5.0 | 4.7 | 3.6 | 5.01 | 4.4 |
| Total C (g kg−1) | 311 | 194.2 | 301.2 | 337.7 | 321.5 | 321.2 | 376.2 | 287.2 | 348.7 | |
| Total N (g kg−1) | 7.8 | 10.5 | 13.7 | 10.67 | 11.5 | 11.8 | 7.7 | 17.3 | 11 | |
| Mineral N (mg kg−1) | 29.2 | 31.7 | 27.5 | 34.2 | 43.5 | 42.0 | 22.5 | 72.0 | 37.2 | |
| C:N ratio | 39.9 | 18.3 | 21.8 | 31.6 | 26.6 | 27.1 | 48.8 | 16.6 | 31.7 | |
| P (mg kg−1) | 81.5 | 36.2 | 59.7 | 69.5 | 79.0 | 61.5 | 129.7 | 90.5 | 161.2 | |
| K (mg kg−1) | 407 | 331 | 444.7 | 552.2 | 585.7 | 480.7 | 460.7 | 524.2 | 635.7 | |
| Basal area (BA, m2 ha−1) | 61.42 | 113.46 | 61.63 | 66.6 | 91.6 | 78.55 | 65.48 | 78.07 | 74.53 | |
| Basal area percentage per species (%) | Populus tremuloides | 3.32 | 81.93 | 58.82 | 5.50 | 87.05 | 63.52 | 2.86 | 90.47 | 68.93 |
| Picea mariana | 72.9 | 16.11 | 26.83 | 80.00 | 7.50 | 24.26 | 96.02 | 8.47 | 29.14 | |
| Abies balsamea | 0.94 | 0.78 | 0.27 | 1.06 | 0.20 | |||||
| Pinus banksiana | 23.78 | 1.02 | 12.21 | 14.50 | 4.35 | 12.22 | 1.12 | 0.78 | ||
| Betula papyrifera | 0.46 | 0.25 | ||||||||
| Populus balsamifera | 0.90 | 0.83 | 0.70 | |||||||
| Organic layer depth (cm) | 17.3 | 6.2 | 10.16 | 20.0 | 7.5 | 11.5 | 14.2 | 9.76 | 13 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghotsa Mekontchou, C.; Houle, D.; Bergeron, Y.; Drobyshev, I. Contrasting Root System Structure and Belowground Interactions between Black Spruce (Picea mariana (Mill.) B.S.P) and Trembling Aspen (Populus tremuloides Michx) in Boreal Mixedwoods of Eastern Canada. Forests 2020, 11, 127. https://doi.org/10.3390/f11020127
Ghotsa Mekontchou C, Houle D, Bergeron Y, Drobyshev I. Contrasting Root System Structure and Belowground Interactions between Black Spruce (Picea mariana (Mill.) B.S.P) and Trembling Aspen (Populus tremuloides Michx) in Boreal Mixedwoods of Eastern Canada. Forests. 2020; 11(2):127. https://doi.org/10.3390/f11020127
Chicago/Turabian StyleGhotsa Mekontchou, Claudele, Daniel Houle, Yves Bergeron, and Igor Drobyshev. 2020. "Contrasting Root System Structure and Belowground Interactions between Black Spruce (Picea mariana (Mill.) B.S.P) and Trembling Aspen (Populus tremuloides Michx) in Boreal Mixedwoods of Eastern Canada" Forests 11, no. 2: 127. https://doi.org/10.3390/f11020127
APA StyleGhotsa Mekontchou, C., Houle, D., Bergeron, Y., & Drobyshev, I. (2020). Contrasting Root System Structure and Belowground Interactions between Black Spruce (Picea mariana (Mill.) B.S.P) and Trembling Aspen (Populus tremuloides Michx) in Boreal Mixedwoods of Eastern Canada. Forests, 11(2), 127. https://doi.org/10.3390/f11020127