Deciphering S-RNase Allele Patterns in Cultivated and Wild Accessions of Italian Pear Germplasm


 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and DNA Extraction
2.2. S-Genotyping Assay
2.3. DNA Sequencing and Allele Identification
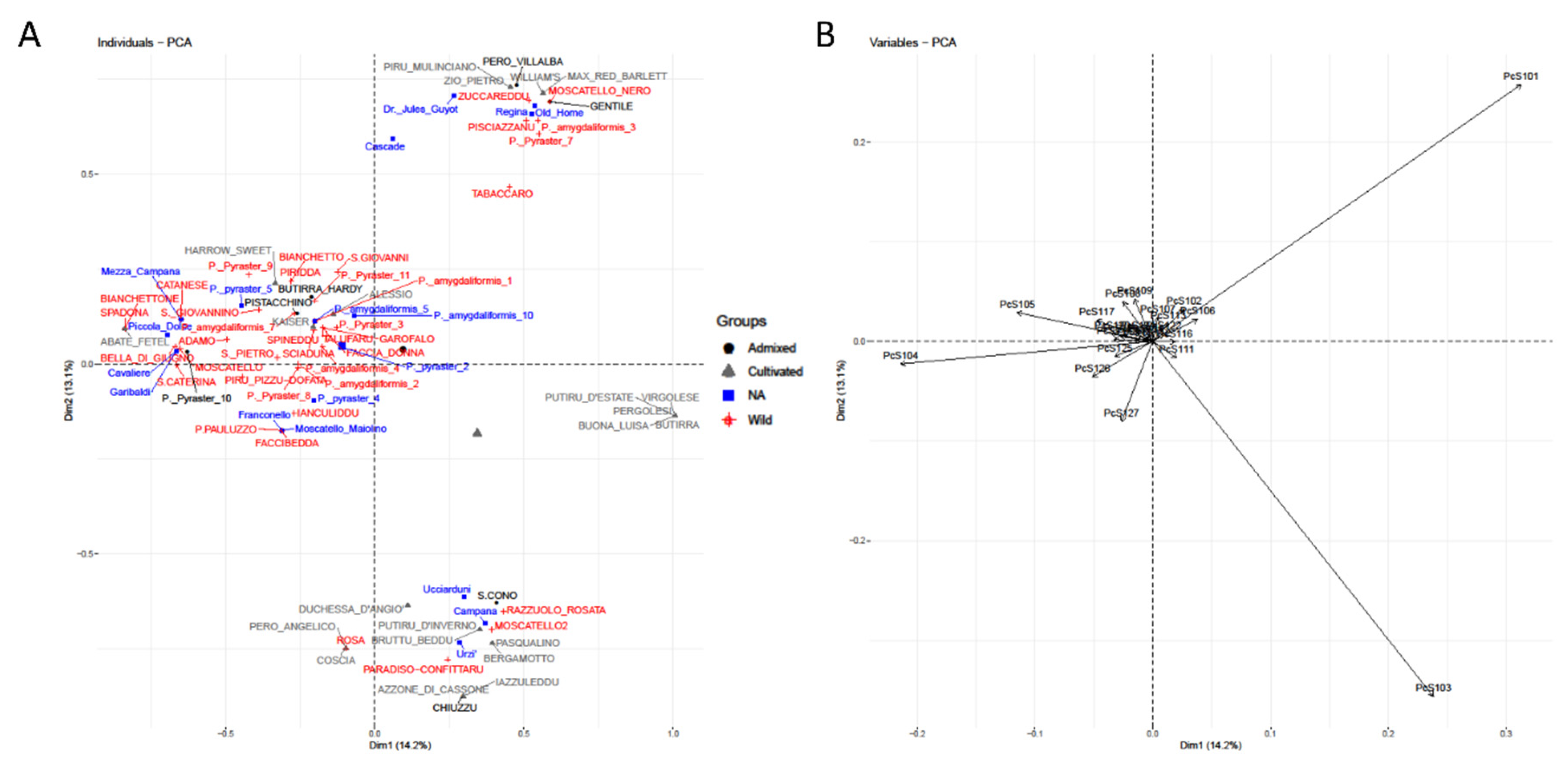
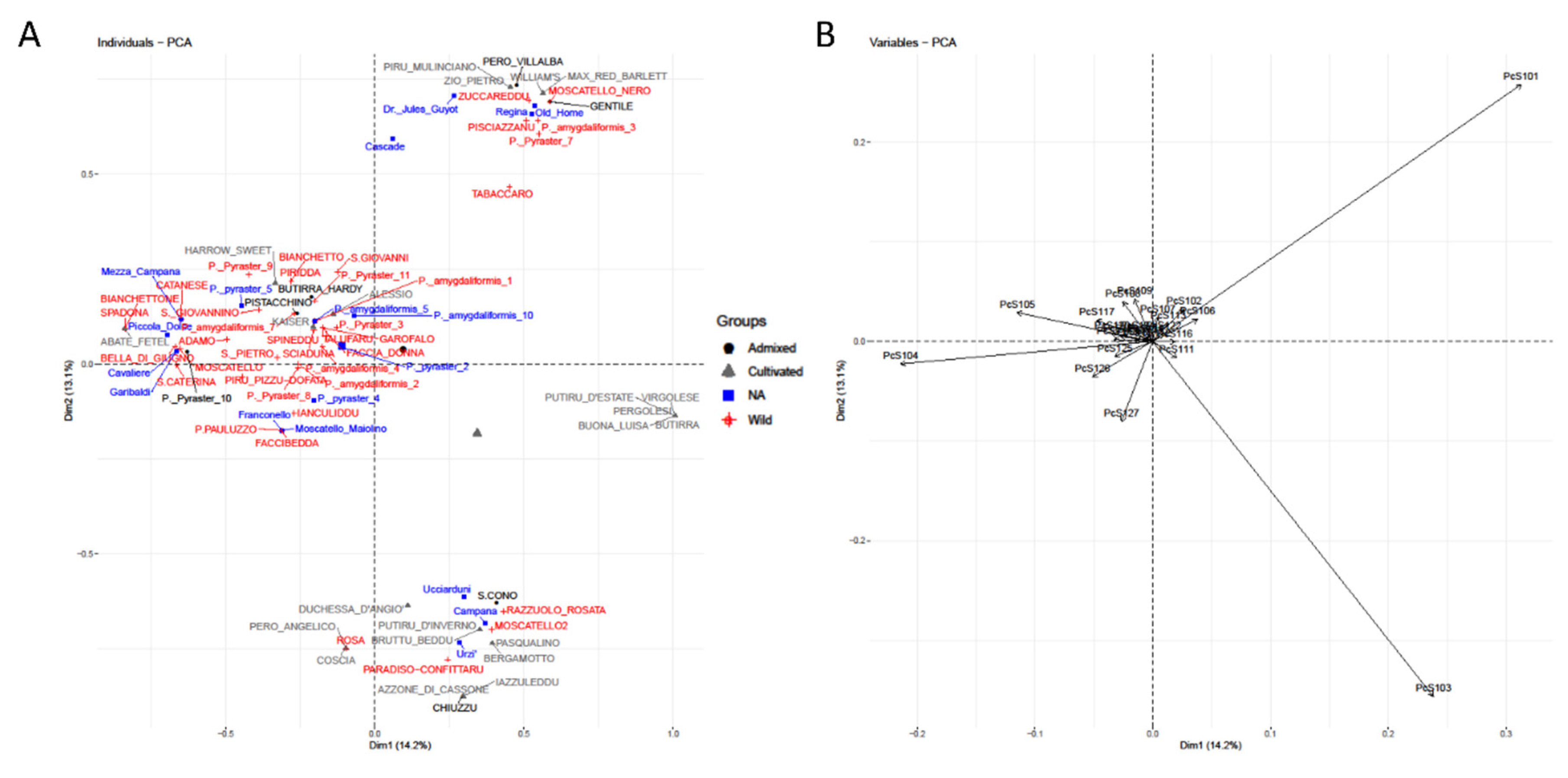
2.4. Clustering
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- McClure, B.; Haring, V.; Ebert, P.; Anderson, M.; Simpson, R.; Sakiyama, F.; Clarke, A. Style self-incompatibility gene products of Nicotiana alata are ribonucleases. Nature 1989, 342, 955–957. [Google Scholar] [CrossRef] [PubMed]
- De Nettancourt, D. Incompatibility and Incongruity in Wild and Cultivated Plants; Springer: Berlin, Germany, 2001; Volume 3. [Google Scholar]
- Broothaerts, W.; Janssens, G.A.; Proost, P.; Broekaert, W.F. cDNA cloning and molecular analysis of two self-incompatibility alleles from apple. Plant Mol. Biol. 1995, 27, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Sassa, H.; Kakui, H.; Miyamoto, M.; Suzuki, Y.; Hanada, T.; Ushijima, K.; Kusaba, M.; Hirano, H.; Koba, T. S Locus F-Box Brothers: Multiple and Pollen-Specific F-Box Genes With S Haplotype-Specific Polymorphisms in Apple and Japanese Pear. Genetics 2007, 175, 1869–1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Franceschi, P.; Dondini, L.; Sanzol, J. Molecular bases and evolutionary dynamics of self-incompatibility in the Pyrinae (Rosaceae). J. Exp. Bot. 2012, 63, 4015–4032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuccherelli, S.; Tassinari, P.; Broothaerts, W.; Tartarini, S.; Dondini, L.; Sansavini, S. S-Allele characterization in self-incompatible pear (Pyrus communis L.). Sex. Plant Reprod. 2002, 15, 153–158. [Google Scholar] [CrossRef]
- Sanzol, J.; Herrero, M.B. Identification of self-incompatibility alleles in pear cultivars (Pyrus communis L.). Euphytica 2002, 128, 325–331. [Google Scholar] [CrossRef]
- Zisovich, A.; Stern, R.; Sapir, G.; Shafir, S.; Goldway, M. The RHV region of S-RNase in the European pear (Pyrus communis) is not required for the determination of specific pollen rejection. Sex. Plant Reprod. 2004, 17, 151–156. [Google Scholar] [CrossRef]
- Sanzol, J.; Sutherland, B.G.; Robbins, T.P. Identification and characterization of genomic DNA sequences of the S-ribonuclease gene associated with self-incompatibility alleles S1 to S5 in European pear. Plant Breed. 2006, 125, 513–518. [Google Scholar] [CrossRef]
- Takasaki, T.; Moriya, Y.; Okada, K.; Yamamoto, K.; Iwanami, H.; Bessho, H.; Nakanishi, T. cDNA cloning of nine S alleles and establishment of a PCR-RFLP system for genotyping European pear cultivars. Theor. Appl. Genet. 2006, 112, 1543–1552. [Google Scholar] [CrossRef]
- Moriya, Y.; Yamamoto, K.; Okada, K.; Iwanami, H.; Bessho, H.; Nakanishi, T.; Takasaki, T. Development of a CAPS marker system for genotyping European pear cultivars harboring 17 S alleles. Plant Cell Rep. 2007, 26, 345–354. [Google Scholar] [CrossRef]
- Sanzol, J.; Robbins, T.P. Combined Analysis of S-Alleles in European Pear by Pollinations and PCR-based S-Genotyping; Correlation between S-Phenotypes and S-RNase Genotypes. J. Am. Soc. Hortic. Sci. 2008, 133, 213–224. [Google Scholar] [CrossRef] [Green Version]
- Goldway, M.; Takasaki-Yasuda, T.; Sanzol, J.; Mota, M.; Zisovich, A.; Stern, R.A.; Sansavini, S. Renumbering the S-RNase alleles of European pears (Pyrus communis L.) and cloning the S109 RNase allele. Sci. Hortic. 2009, 119, 417–422. [Google Scholar] [CrossRef]
- Sanzol, J. Genomic characterization of self-incompatibility ribonucleases (S-RNases) in European pear cultivars and development of PCR detection for 20 alleles. Tree Genet. Genomes 2009, 5, 393–405. [Google Scholar] [CrossRef]
- Sanzol, J. Pistil-function breakdown in a new S-allele of European pear, S21°, confers self-compatibility. Plant Cell Rep. 2008, 28, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Gharehaghaji, A.N.; Arzani, K.; Abdollahi, H.; Shojaeiyan, A.; Dondini, L.; De Franceschi, P. Genomic characterization of self-incompatibility ribonucleases in the Central Asian pear germplasm and introgression of new alleles from other species of the genus Pyrus. Tree Genet. Genomes 2014, 10, 411–428. [Google Scholar] [CrossRef]
- Bennici, S.; Casas, G.L.; Distefano, G.; Di Guardo, M.; Continella, A.; Ferlito, F.; Gentile, A.; La Malfa, S. Elucidating the contribution of wild related species on autochthonous pear germplasm: A case study from Mount Etna. PLoS ONE 2018, 13, e0198512. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, K.; Tamura, F.; Chun, J.-P.; Tanabe, K. Native Mediterranean Pyrus Rootstock, P. amygdaliformis and P. elaeagrifolia, Present Higher Tolerance to Salinity Stress Compared with Asian Natives. J. Jpn. Soc. Hortic. Sci. 2006, 75, 450–457. [Google Scholar] [CrossRef] [Green Version]
- Łukasz, W.; Antkowiak, W.; Lenartowicz, E.; Bocianowski, J. Genetic diversity of European pear cultivars (Pyrus communis L.) and wild pear (Pyrus pyraster (L.) Burgsd.) inferred from microsatellite markers analysis. Genet. Resour. Crop. Evol. 2010, 57, 801–806. [Google Scholar] [CrossRef] [Green Version]
- Łukasz, W.; Antkowiak, W.; Sips, M.; Slomski, R. Self-incompatibility alleles in Polish wild pear (Pyrus pyraster (L.) Burgsd.): A preliminary analysis. J. Appl. Genet. 2010, 51, 33–35. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.G.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Tassinari, P. Analisi molecolari ed agronomiche dell’incompatibilita’ gemetofitica in pero (Pyrus communis) e ciliegio (Prunus avium e P. cerasus): Ricerche sul locus S. Ph.D. Thesis, University of Bologna, Bologna, Italy, 2005. [Google Scholar]
- Mota, M.; Tavares, L.; Oliveira, C.M. Identification of S-alleles in pear (Pyrus communis L.) cv. ‘Rocha’ and other European cultivars. Sci. Hortic. 2007, 113, 13–19. [Google Scholar] [CrossRef]
- Takasaki-Yasuda, T.; Nomura, N.; Moriya-Tanaka, Y.; Okada, K.; Iwanami, H.; Bessho, H. Cloning anS-RNaseallele, including the longest intron, from cultivars of European pear (Pyrus communis L.). J. Hortic. Sci. Biotechnol. 2013, 88, 427–432. [Google Scholar] [CrossRef]
- Rehder, A. A Manual of Cultivated Trees and Shrubs Hardy in North American Exclusive of the Subtropical and Warmer Temperate Regions, 2nd ed.; Macmillan: New York, NY, USA, 1940; p. 403. [Google Scholar]
- Katayama, H.; Tachibana, M.; Iketani, H.; Zhang, S.-L.; Uematsu, C. Phylogenetic utility of structural alterations found in the chloroplast genome of pear: Hypervariable regions in a highly conserved genome. Tree Genet. Genomes 2011, 8, 313–326. [Google Scholar] [CrossRef]
- Volk, G.M.; Henk, A.D.; Richards, C.M.; Bassil, N.; Postman, J. Chloroplast sequence data differentiate Maleae, and specifically Pyrus, species in the USDA-ARS National Plant Germplasm System. Genet. Resour. Crop. Evol. 2018, 66, 5–15. [Google Scholar] [CrossRef]
- Liu, Q.; Song, Y.; Liu, L.; Zhang, M.; Sun, J.; Zhang, S.; Wu, J. Genetic diversity and population structure of pear (Pyrus spp.) collections revealed by a set of core genome-wide SSR markers. Tree Genet. Genomes 2015, 11, 128. [Google Scholar] [CrossRef]
- Teng, Y.; Tanabe, K.; Tamura, F.; Itai, A. Genetic Relationships of Pyrus Species and Cultivars Native to East Asia Revealed by Randomly Amplified Polymorphic DNA Markers. J. Am. Soc. Hortic. Sci. 2002, 127, 262–270. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Kirk, C.; Deng, C.; Wiedow, C.; Knaebel, M.; Brewer, L. Genotyping-by-sequencing of pear (Pyrus spp.) accessions unravels novel patterns of genetic diversity and selection footprints. Hortic. Res. 2017, 4, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Dolatowski, J.; Podyma, W.; Szymañska, M.; Zych, M. Molecular studies on the variability of Polish semi-wild pears (Pyrus) using AFLP. J. Fruit Ornam. Plant Res. 2004, 12, 331–337. [Google Scholar]
- Volk, G.M.; Cornille, A. Genetic Diversity and Domestication History in Pyrus. In The Pear Genome; Korban, S.S., Ed.; Springer: Cham, Germany, 2019; pp. 51–62. [Google Scholar]
- Wright, S. The Distribution of Self-Incompatibility Alleles in Populations. Evolution 1964, 18, 609. [Google Scholar] [CrossRef]
- Ioerger, T.R.; Clark, A.G.; Kao, T.H. Polymorphism at the self-incompatibility locus in Solanaceae predates speciation. Proc. Natl. Acad. Sci. USA 1990, 87, 9732–9735. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
| Species | Accession Name | Origin | Status a | Collection b | Q1 | Q2 | Subpopulation c | S-Genotype |
|---|---|---|---|---|---|---|---|---|
| Pyrus communis | Adamo | Italy (Sicily) | LV | UNICT | 0.053 | 0.947 | Wild | PcS105/PcS126 |
| P. communis | Alessio | Italy (Sicily) | LV | Mt Etna | 0.97 | 0.03 | Cultivated | PcS107/- |
| P. communis | Angelico | Italy (Sicily) | NCV | Mt Etna | 0.989 | 0.011 | Cultivated | PcS104/PcS103 |
| P. communis | Azzone di Cassone | Italy (Sicily) | LV | UNICT | 0.972 | 0.028 | Cultivated | PcS103/PcS127 |
| P. communis | Bella di Giugno | Italy (Sicily) | NCV | Mt Etna | 0.054 | 0.946 | Wild | PcS104/PcS120 * |
| P. communis | Bergamotto | Italy (Sicily) | NCV | UNICT | 0.984 | 0.016 | Cultivated | PcS103/PcS111 |
| P. communis | Bianchetto | Italy (Sicily) | NCV | UNICT | 0.012 | 0.988 | Wild | PcS109/PcS117 |
| P. communis | Bianchettone | Italy (Sicily) | LV | Mt Etna | 0.032 | 0.968 | Wild | PcS104/PcS105 |
| P. communis | Bruttu Beddu | Italy (Sicily) | LV | Mt Etna | 0.929 | 0.071 | Cultivated | PcS103/- |
| P. communis | Buona Luisa | Italy (Sicily) | NCV | Mt Etna | 0.993 | 0.007 | Cultivated | PcS101/PcS103 |
| P. communis | Butirra | Italy (Sicily) | NCV | UNICT | 0.991 | 0.009 | Cultivated | PcS101/PcS103 |
| P. communis | Campana | Italy (Sicily) | NCV | UNICT | NA | PcS103/PcS122 | ||
| P. communis | Catanese | Italy (Sicily) | LV | Mt Etna | 0.019 | 0.981 | Wild | PcS104/PcS108 |
| P. communis | Cavaliere | Italy (Sicily) | LV | UNICT | NA | PcS104/PcS115 | ||
| P. communis | Chiuzzu | Italy (Sicily) | LV | UNICT | 0.215 | 0.785 | Admixed | PcS103/PcS127 |
| P. communis | Coscia | Italy (Sicily) | NCV | UNICT | 0.99 | 0.01 | Cultivated | PcS104/PcS103 * |
| P. communis | Duchessa d’Angio’ | Italy (Sicily) | LV | Mt Etna | 0.98 | 0.02 | Cultivated | PcS103/PcS105 |
| P. communis | Faccia Donna | Italy (Sicily) | LV | UNICT | 0.009 | 0.991 | Wild | PcS124/- |
| P. communis | Faccibedda | Italy (Sicily) | LV | UNICT | 0.018 | 0.982 | Wild | PcS126/PcS127 |
| P. communis | Franconello | Italy (Sicily) | LV | Mt Etna | NA | PcS126/PcS127 | ||
| P. communis | Garibaldi | Italy (Sicily) | LV | Mt Etna | NA | PcS104/PcS115 | ||
| P. communis | Garofalo | Italy (Sicily) | NCV | Mt Etna | 0.009 | 0.991 | Wild | PcS124/- |
| P. communis | Gentile | Italy (Sicily) | NCV | UNICT | 0.644 | 0.356 | Admixed | PcS101/PcS106 * |
| P. communis | Ialufaru | Italy (Sicily) | LV | Mt Etna | 0.014 | 0.986 | Wild | PcS124/- |
| P. communis | Ianculiddu | Italy (Sicily) | LV | Mt Etna | 0.012 | 0.988 | Wild | PcS125/PcS127 |
| P. communis | Iazzuleddu | Italy (Sicily) | LV | UNICT | 0.984 | 0.016 | Cultivated | PcS103/PcS127 |
| P. communis | Mezza Campana | Italy (Sicily) | LV | Mt Etna | NA | PcS104/PcS108 | ||
| P. communis | Moscatello 1 | Italy (Sicily) | NCV | Mt Etna | 0.018 | 0.982 | Wild | PcS104/PcS123 |
| P. communis | Moscatello 2 | Italy (Sicily) | NCV | UNICT | 0.027 | 0.973 | Wild | PcS103/PcS116 |
| P. communis | Moscatello Maiolino | Italy (Sicily) | LV | Mt Etna | NA | PcS126/PcS127 | ||
| P. communis | Moscatello Nero | Italy (Sicily) | LV | Mt Etna | 0.014 | 0.986 | Cultivated | PcS101/PcS106 |
| P. communis | Paradiso/Confittaru | Italy (Sicily) | LV | Mt Etna | 0.081 | 0.919 | Wild | PcS103/PcS126 |
| P. communis | Pasqualino | Italy (Sicily) | LV | UNICT | 0.968 | 0.032 | Cultivated | PcS103/PcS111 |
| P. communis | Pauluzzo | Italy (Sicily) | LV | Mt Etna | 0.018 | 0.982 | Wild | PcS126/PcS127 |
| P. communis | Pergolesi | Italy (Sicily) | LV | Mt Etna | 0.989 | 0.011 | Cultivated | PcS101/PcS103 |
| P. communis | Piccola Dolce | Italy (Sicily) | LV | Mt Etna | NA | PcS104/PcS117 | ||
| P. communis | Piridda | Italy (Sicily) | LV | Mt Etna | 0.007 | 0.993 | Wild | PcS109/PcS117 |
| P. communis | Piru Mulinciana | Italy (Sicily) | LV | Mt Etna | 0.99 | 0.01 | Cultivated | PcS101/PcS108 |
| P. communis | Piru Pizzu | Italy (Sicily) | LV | Mt Etna | 0.139 | 0.861 | Wild | PcS105/PcS127 |
| P. communis | Pisciazzanu | Italy (Sicily) | LV | Mt Etna | 0.056 | 0.944 | Wild | PcS101/- |
| P. communis | Pistacchino | Italy (Sicily) | LV | Mt Etna | 0.699 | 0.301 | Admixed | PcS117/PcS121 |
| P. communis | Putiru d’Estate | Italy (Sicily) | LV | Mt Etna | 0.985 | 0.015 | Cultivated | PcS101/PcS103 |
| P. communis | Putiru d’Inverno | Italy (Sicily) | LV | UNICT | 0.992 | 0.008 | Cultivated | PcS103/- |
| P. communis | Razzuolo Rosata | Italy (Sicily) | LV | Mt Etna | 0.017 | 0.983 | Wild | PcS103/PcS106 |
| P. communis | Regina | Italy (Sicily) | LV | UNICT | NA | PcS101/PcS122 | ||
| P. communis | Rosa | Italy (Sicily) | LV | Mt Etna | 0.01 | 0.99 | Wild | PcS104/PcS103 |
| P. communis | San Cono | Italy (Sicily) | LV | Mt Etna | 0.651 | 0.349 | Admixed | PcS102/PcS103 |
| P. communis | San Giovanni | Italy (Sicily) | NCV | UNICT | 0.009 | 0.991 | Wild | PcS102/PcS109 |
| P. communis | San Giovannino | Italy (Sicily) | LV | Mt Etna | 0.01 | 0.99 | Wild | PcS105/- |
| P. communis | San Pietro | Italy (Sicily) | NCV | UNICT | 0.015 | 0.985 | Wild | PcS120/PcS126 |
| P. communis | Santa Caterina | Italy (Sicily) | LV | UNICT | 0.01 | 0.99 | Wild | PcS104/PcS125 |
| P. communis | Sciaduna | Italy (Sicily) | LV | Mt Etna | 0.05 | 0.95 | Wild | PcS111/PcS117 |
| P. communis | Spadona | Italy (Sicily) | NCV | Mt Etna | 0.029 | 0.971 | Wild | PcS104/PcS105 |
| P. communis | Spineddu | Italy (Sicily) | NCV | UNICT | 0.039 | 0.961 | Wild | PcS111/PcS117 |
| P. communis | Tabaccaro | Italy (Sicily) | LV | Mt Etna | 0.029 | 0.971 | Wild | PcS101/PcS127 |
| P. communis | Ucciarduni | Italy (Sicily) | NCV | UNICT | NA | PcS103/PcS108 | ||
| P. communis | Urzi’ | Italy (Sicily) | LV | UNICT | NA | PcS103/PcS125 | ||
| P. communis | Villalba | Italy (Sicily) | LV | Mt Etna | 0.276 | 0.724 | Admixed | PcS101/PcS109 |
| P. communis | Virgolese | Italy (Sicily) | NCV | UNICT | 0.99 | 0.01 | Cultivated | PcS101/PcS103 |
| P. communis | Zio Pietro | Italy (Sicily) | LV | UNICT | 0.986 | 0.014 | Cultivated | PcS101/PcS108 |
| P. communis | Zuccareddu | Italy (Sicily) | LV | Mt Etna | 0.034 | 0.966 | Wild | PcS101/PcS107 |
| P. amygdaliformis | 1 | Italy (Sicily) | RS | North (N) 37° 47′ 692″ east (E) 14° 50′ 925″ | 0.013 | 0.987 | Wild | PcS120/PcS122 |
| P. amygdaliformis | 2 | Italy (Sicily) | RS | N 37° 51′ 189″ E 14° 50′ 795″ | 0.008 | 0.992 | Wild | PcS126/- |
| P. amygdaliformis | 3 | Italy (Sicily) | RS | N 37° 51′ 147″ E 14° 50′ 739″ | 0.007 | 0.993 | Wild | PcS101/PcS116 |
| P. amygdaliformis | 4 | Italy (Sicily) | RS | N 37° 51′ 109″ E 14°50′ 809″ | 0.007 | 0.993 | Wild | PcS116/PcS125 |
| P. amygdaliformis | 5 | Italy (Sicily) | RS | N 37° 51′ 116″ E 14° 50′ 806″ | NA | PcS120/PcS122 | ||
| P. amygdaliformis | 7 | Italy (Sicily) | RS | N 37° 51′ 139″ E 14° 50′ 765″ | 0.009 | 0.991 | Wild | PcS108/PcS125 |
| P. amygdaliformis | 10 | Italy (Sicily) | RS | N 37° 51′ 137″ E 14° 50′ 748″ | NA | PcS106/- | ||
| P. pyraster | 2 | Italy (Sicily) | RS | N 37° 48′ 143″ E 14° 51′ 203″ | NA | PcS111/- | ||
| P. pyraster | 3 | Italy (Sicily) | RS | N 37° 51′ 796″ E 14° 52′ 409″ | 0.036 | 0.964 | Wild | PcS122/- |
| P. pyraster | 4 | Italy (Sicily) | RS | N 37° 52′ 808″ E 14° 52′ 571″ | NA | PcS127/- | ||
| P. pyraster | 5 | Italy (Sicily) | RS | N 37° 52′ 868″ E 14° 52′ 565″ | NA | PcS105/PcS123 | ||
| P. pyraster | 7 | Italy (Sicily) | RS | N 37° 47′ 691″ E 14° 50′ 901″ | 0.097 | 0.903 | Wild | PcS101/PcS111 |
| P. pyraster | 8 | Italy (Sicily) | RS | N 37° 52′ 006″ E 14° 52′ 969″ | 0.007 | 0.993 | Wild | PcS108/PcS127 |
| P. pyraster | 9 | Italy (Sicily) | RS | N 37° 51′ 865″ E 14° 52′ 672″ | 0.009 | 0.991 | Wild | PcS105/PcS109 |
| P. pyraster | 10 | Italy (Sicily) | RS | N 37° 48′ 276″ E 14° 51′ 269″ | 0.477 | 0.523 | Admixed | PcS104/PcS110 |
| P. pyraster | 11 | Italy (Sicily) | RS | N 37° 53′ 038″ E 14° 52′ 578″ | 0.057 | 0.943 | Wild | PcS108/- |
| P. communis | Abbé Fétel | France | ICV | UNIBO | 0.967 | 0.033 | Cultivated | PcS104/PcS105 * |
| P. communis | Beurre Hardy | USA | ICV | UNIBO | 0.391 | 0.609 | Admixed | PcS108/PcS114 * |
| P. communis | Cascade | USA | ICV | UNIBO | NA | PcS101/PcS104 * | ||
| P. communis | Dr. Jules Guyot | France | ICV | UNIBO | NA | PcS101/PcS105 * | ||
| P. communis | Kaiser | France | ICV | UNIBO | 0.914 | 0.086 | Cultivated | PcS107/PcS125 * |
| P. communis | Max Red Barlet | USA | ICV | UNIBO | 0.993 | 0.007 | Cultivated | PcS101/PcS102 * |
| P. communis | Old Home | USA | ICV | UNIBO | NA | PcS101/PcS113 * | ||
| P. communis | Harrow Sweet | Canada | ICV | UNIBO | 0.987 | 0.013 | Cultivated | PcS102/PcS105 * |
| P. communis | Williams | England | ICV | UNIBO | 0.993 | 0.007 | Cultivated | PcS101/PcS102 * |
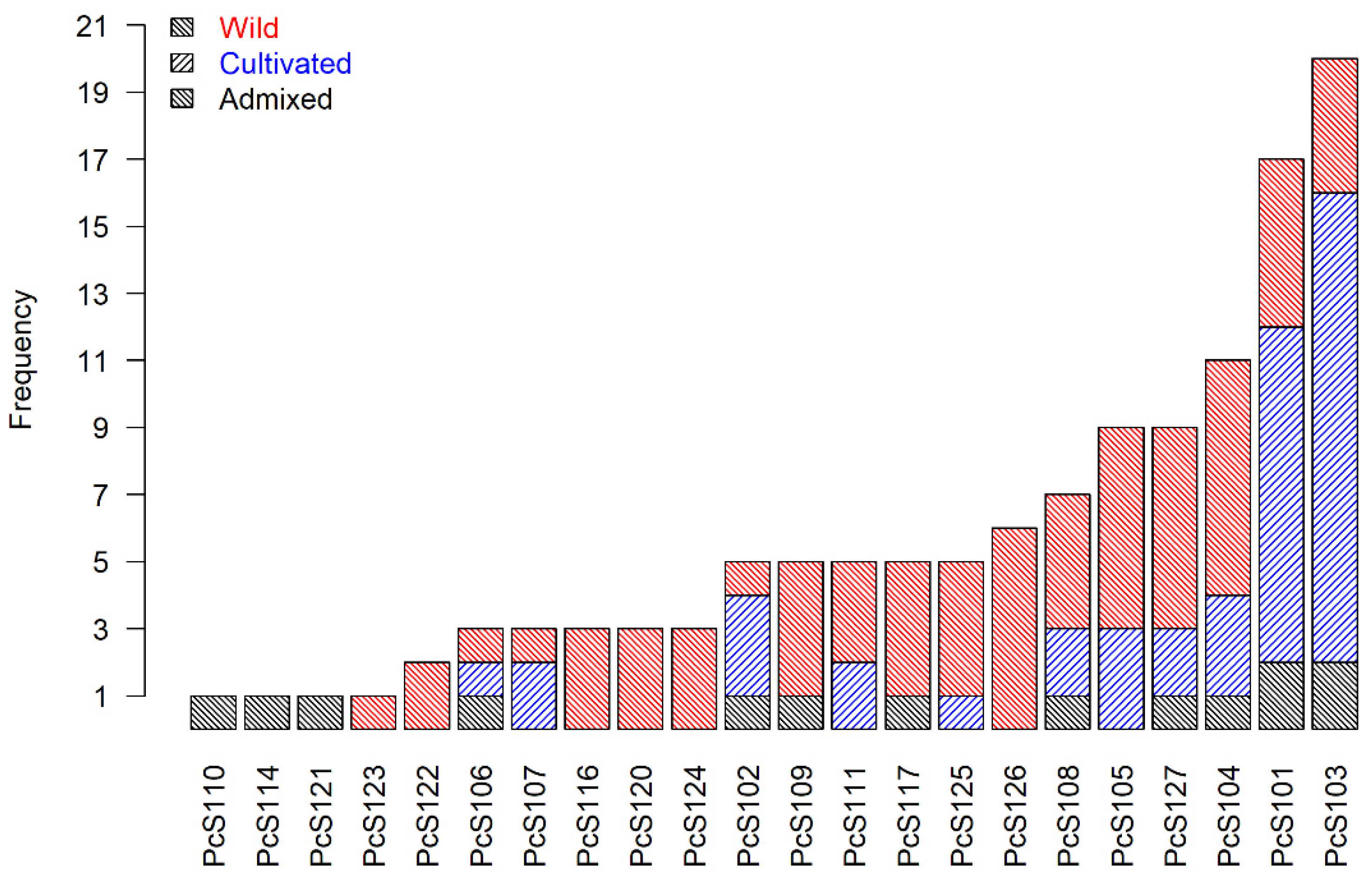
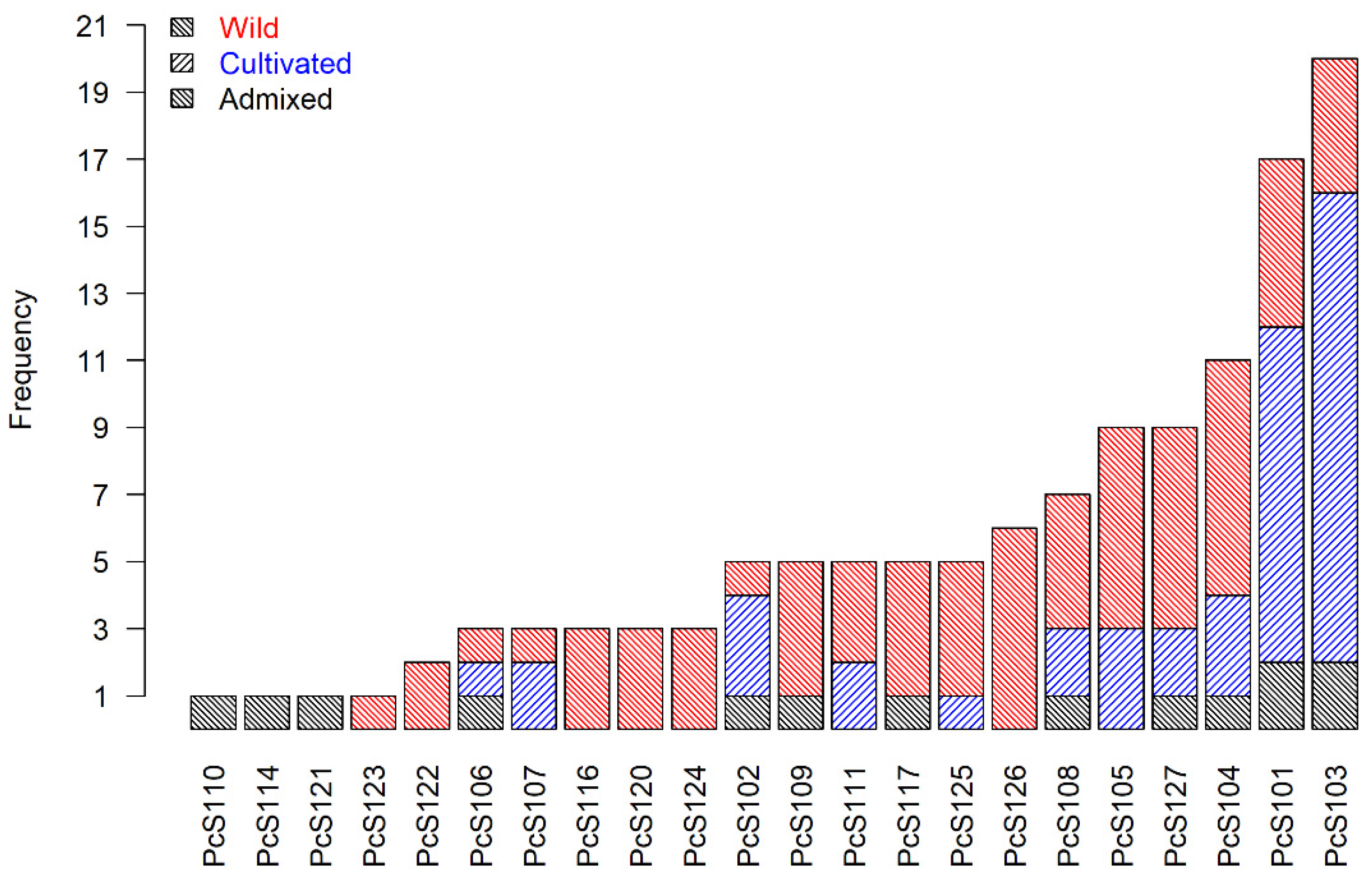
| S-Allele | Count | RS | LV | NCV | ICV |
|---|---|---|---|---|---|
| PcS101 | 21 | 0.1 | 0.48 | 0.19 | 0.24 |
| PcS102 | 5 | 0 | 0.2 | 0.2 | 0.6 |
| PcS103 | 23 | 0 | 0.61 | 0.39 | 0 |
| PcS104 | 16 | 0.06 | 0.5 | 0.31 | 0.13 |
| PcS105 | 11 | 0.18 | 0.45 | 0.09 | 0.27 |
| PcS106 | 4 | 0.25 | 0.5 | 0.25 | 0 |
| PcS107 | 3 | 0 | 0.67 | 0 | 0.33 |
| PcS108 | 9 | 0.33 | 0.44 | 0.11 | 0.11 |
| PcS109 | 5 | 0.2 | 0.4 | 0.4 | 0 |
| PcS110 | 1 | 1 | 0 | 0 | 0 |
| PcS111 | 6 | 0.33 | 0.33 | 0.33 | 0 |
| PcS113 | 1 | 0 | 0 | 0 | 1 |
| PcS114 | 1 | 0 | 0 | 0 | 1 |
| PcS115 | 2 | 0 | 1 | 0 | 0 |
| PcS116 | 3 | 0.67 | 0 | 0.33 | 0 |
| PcS117 | 6 | 0 | 0.67 | 0.33 | 0 |
| PcS120 | 4 | 0.5 | 0 | 0.5 | 0 |
| PcS121 | 1 | 0 | 1 | 0 | 0 |
| PcS122 | 5 | 0.6 | 0.2 | 0.2 | 0 |
| PcS123 | 2 | 0.5 | 0 | 0.5 | 0 |
| PcS124 | 3 | 0 | 0.67 | 0.33 | 0 |
| PcS125 | 6 | 0.33 | 0.5 | 0 | 0.17 |
| PcS126 | 8 | 0.13 | 0.75 | 0.13 | 0 |
| PcS127 | 12 | 0.17 | 0.83 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bennici, S.; Di Guardo, M.; Distefano, G.; Las Casas, G.; Ferlito, F.; De Franceschi, P.; Dondini, L.; Gentile, A.; La Malfa, S. Deciphering S-RNase Allele Patterns in Cultivated and Wild Accessions of Italian Pear Germplasm. Forests 2020, 11, 1228. https://doi.org/10.3390/f11111228
Bennici S, Di Guardo M, Distefano G, Las Casas G, Ferlito F, De Franceschi P, Dondini L, Gentile A, La Malfa S. Deciphering S-RNase Allele Patterns in Cultivated and Wild Accessions of Italian Pear Germplasm. Forests. 2020; 11(11):1228. https://doi.org/10.3390/f11111228
Chicago/Turabian StyleBennici, Stefania, Mario Di Guardo, Gaetano Distefano, Giuseppina Las Casas, Filippo Ferlito, Paolo De Franceschi, Luca Dondini, Alessandra Gentile, and Stefano La Malfa. 2020. "Deciphering S-RNase Allele Patterns in Cultivated and Wild Accessions of Italian Pear Germplasm" Forests 11, no. 11: 1228. https://doi.org/10.3390/f11111228