Identification of an Emaravirus in a Common Oak (Quercus robur L.) Conservation Seed Orchard in Germany: Implications for Oak Health


Abstract
:1. Introduction
2. Materials and Methods
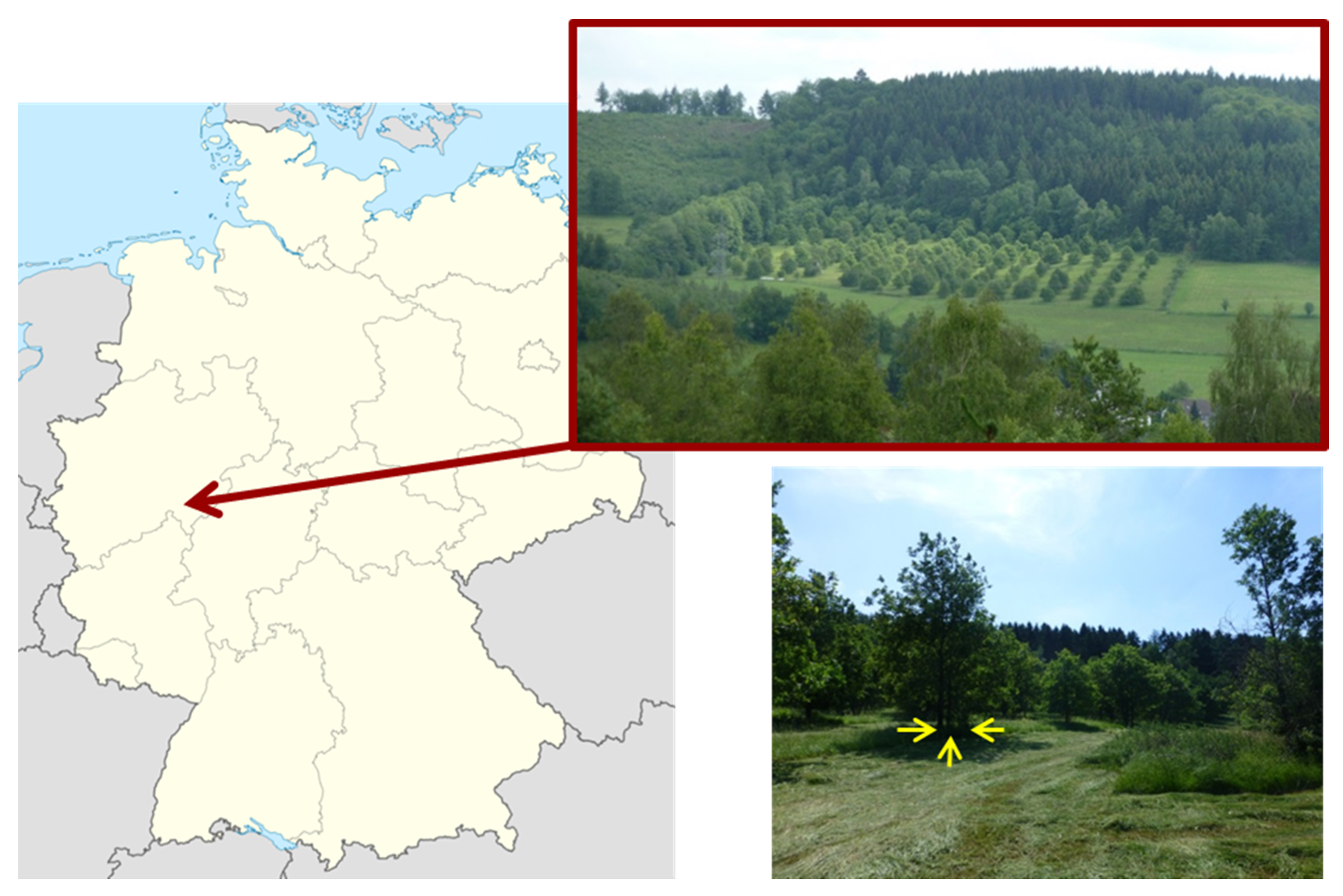
2.1. Conservation Seed Orchard
2.2. RNASeq of Diseased Oak Leaves and Data Analyses
2.3. RT-PCR for Plant Virus Diagnosis
3. Results
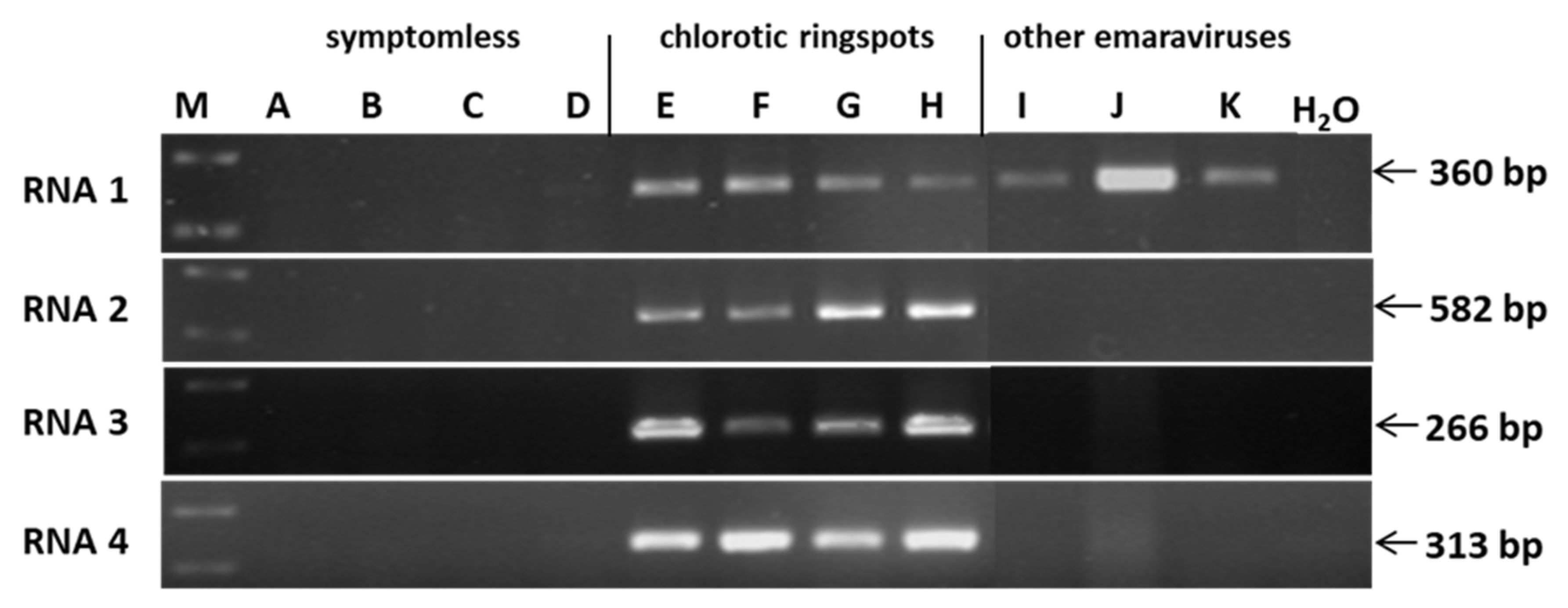
3.1. Identification of a Novel Virus in Oak
3.2. Detection of the Novel Virus in Ringspot Diseased oak Trees
3.3. Monitoring
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Eaton, E.; Caudullo, G.; Oliveira, S.; de Rigo, D. Quercus robur and Quercus petraea in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the EU: Luxembourg, 2016; p. e01c6df+. [Google Scholar]
- Hemery, G.E. Forest management and silvicultural responses to projected climate change impacts on European broadleaved trees and forests. Int. For. Rev. 2008, 10, 591–607. [Google Scholar] [CrossRef]
- BMEL. The Forests in Germany-Selected results of the Third National Forest Inventory. 2015. Available online: www.bmel.de/SharedDocs/Downloads/EN/Publications/ForestsInGermany-BWI.pdf?__blob=publicationFile (accessed on 24 September 2020).
- Gibbs, J.N.; Greig, B.J.W. Biotic and abiotic factors affecting the dying back of pedunculate oak Quercus robur L. Forestry 1997, 70, 399–406. [Google Scholar] [CrossRef] [Green Version]
- Haavik, L.J.; Billings, S.A.; Guldin, J.M.; Stephen, F.M. Emergent insects, pathogens and drought shape changing patterns in oak decline in North America and Europe. For. Ecol. Manag. 2015, 354, 190–205. [Google Scholar] [CrossRef]
- Nienhaus, F. Infectious diseases in forest trees caused by viruses, mycoplasma-likeorganisms and primitive bacteria. Experientia 1985, 41, 597–603. [Google Scholar] [CrossRef]
- Samara, R.; Hunter, D.M.; Stobbs, L.W.; Greig, N.; Lowery, D.T.; Delury, C. Impact of Plum pox virus (PPV-D) infection on peach tree growth, productivity and bud cold hardiness. Can. J. Plant Pathol. 2017, 39, 218–228. [Google Scholar] [CrossRef]
- Nemeth, M.V. On the distribution and economic significance of fruit trees in Hungary. Növényvédelem 1992, 28, 26–32. [Google Scholar]
- Büttner, C.; Führling, M. Studies on virus infection of diseased Quercus robur (L.) from forest stands in northern Germany. Ann. Sci. For. 1996, 53, 383–388. [Google Scholar]
- Nienhaus, F. Viren und virusverdächtige Erkrankungen in Eichen (Quercus robur und Quercus sessiliflora). Z. Pflkrankh. Pflschutz 1975, 82, 739–749. [Google Scholar]
- Nienhaus, F.; Castello, J.D. Viruses in forest trees. Ann. Rev. Phytopath 1989, 27, 165–186. [Google Scholar] [CrossRef]
- Büttner, C.; Führling, M. Beobachtungen zu virusbedingten Symptomen an erkrankten Stieleichen (Quercus robur L.)–eine Abgrenzung zu ähnlichen, nicht virusbedingten Krankheitsbildern. Nachr. Dtsch. Pflanzenschutzd. 1993, 45, 110–115. [Google Scholar]
- Adams, I.P.; Fox, A.; Boonham, N.; Massart, S.; De Jonghe, K. The impact of high throughput sequencing on plant health diagnostics. Eur. J. Plant Pathol. 2018, 152, 909–919. [Google Scholar] [CrossRef]
- Roossinck, M.J.; Martin, D.P.; Roumagnac, P. Plant virus metagenomics: Advances in virus discovery. Phytopathology 2015, 105, 716–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huson, D.H.; Auch, A.F.; Qi, J.; Schuster, S.C. MEGAN analysis of metagenomic data. Genome Res. 2007, 17, 377–386. [Google Scholar] [CrossRef] [Green Version]
- Di Bello, P.L.; Ho, T.; Tzanetakis, I.E. The evolution of emaraviruses is becoming more complex: Seven segments identified in the causal agent of Rose rosette disease. Virus Res. 2015, 210, 241–244. [Google Scholar] [CrossRef]
- von Bargen, S.; Dieckmann, H.L.; Candresse, T.; Mühlbach, H.P.; Roßbach, J.; Büttner, C. Determination of the complete genome sequence of European mountain ash ringspot-associated emaravirus from Sorbus intermedia reveals two additional genome segments. Arch. Virol. 2019, 164, 1937–1941. [Google Scholar] [CrossRef]
- Boom, R.C.J.A.; Sol, C.J.; Salimans, M.M.; Jansen, C.L.; Wertheim-van Dillen, P.M.; Van der Noordaa, J.P.M.E. Rapid and simple method for purification of nucleic acids. J. Clin. Microbiol. 1990, 28, 495–503. [Google Scholar] [CrossRef] [Green Version]
- Menzel, W.; Jelkmann, W.; Maiss, E. Detection of four apple viruses by multiplex RT-PCR assays with coamplification of plant mRNA as internal control. J. Virol. Methods 2002, 99, 81–92. [Google Scholar] [CrossRef]
- Elbeaino, T.; Whitfield, A.; Sharma, M.; Digiaro, M. Emaravirus-specific degenerate PCR primers allowed the identification of partial RNA-dependent RNA polymerase sequences of Maize red stripe virus and Pigeonpea sterility mosaic virus. J. Virol. Methods 2013, 188, 37–40. [Google Scholar] [CrossRef] [Green Version]
- Löf, M.; Rydberg, D.; Bolte, A. Mounding site preparation for forest restoration: Survival and short-term growth response in Quercus robur L. seedlings. For. Ecol. Manag. 2006, 232, 19–25. [Google Scholar] [CrossRef]
- Saha, S.; Kuehne, C.; Bauhus, J. Lessons learned from oak cluster planting trials in central Europe. Can. J. For. Res. 2017, 47, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.; Kuehne, C.; Kohnle, U.; Brang, P.; Ehring, A.; Geisel, J.; Leder, B.; Muth, M.; Petersen, R.; Peter, J.; et al. Growth and quality of young oaks (Quercus robur and Q. petraea) grown in cluster plantings in central Europe: A weighted meta-analysis. For. Ecol. Manag. 2012, 283, 106–118. [Google Scholar] [CrossRef]
- Thomas, F.M.; Blank, R.; Hartmann, G. Abiotic and biotic factors and their interactions as causes of oak decline in Central Europe. For. Pathol. 2002, 32, 277–307. [Google Scholar] [CrossRef]
- Dong, P.H.; Eder, W.; Muth, M. Eichen-Nesterpflanzungsversuche in Rheinland-Pfalz- Ergebnisse eines 15jährigen Beobachtungszeitraums. In Eiche im Pfälzerwald. Struktur- und Genehmigungsdirektion Süd, Forschungsanstalt für Waldökologie und Forstwirtschaft Rheinland-Pfalz; Trippstadt, Germany, 2007; pp. 4–22. Available online: https://docplayer.org/40677562-Eichen-nesterpflanzungsversuche-in-rheinland-pfalz-ergebnisse-eines-15jaehrigen-beobachtungszeitraums-p-h-dong-1-m-muth-1-und-w.html (accessed on 25 September 2020).
- Petersen, R. Eichen-Trupp-Pflanzung–erste Ergebnisse einer Versuchsfläche im NFA Neuhaus. Forst Holz 2007, 62, 19–25. [Google Scholar]
- Marçais, B.; Desprez-Loustau, M. European oak powdery mildew: Impact on trees, effects of environmental factors, and potential effects of climate change. Ann. For. Sci. 2014, 71, 633–642. [Google Scholar] [CrossRef] [Green Version]
- Hajji, M.; Dreyer, E.; Marçais, B. Impact of Erysiphe alphitoides on transpiration and photosynthesis in Quercus robur leaves. Eur. J. Plant Pathol. 2009, 125, 63–72. [Google Scholar] [CrossRef] [Green Version]
- García, J.A.; Glasa, M.; Cambra, M.; Candresse, T. Plum pox virus and sharka: A model potyvirus and a major disease. Mol. Plant Pathol. 2014, 15, 226–241. [Google Scholar] [CrossRef]
- Clemente-Moreno, M.J.; Hernández, J.A.; Díaz-Vivancos, P. Sharka: How do plants respond to Plum pox virus infection? J. Exp. Bot. 2015, 66, 25–35. [Google Scholar] [CrossRef] [Green Version]
- Mielke, N.; Mühlbach, H.P. A novel, multipartite, negative-strand RNA virus is associated with the ringspot disease of European mountain ash (Sorbus aucuparia L.). J. Gen. Virol. 2007, 88, 1337–1346. [Google Scholar] [CrossRef]
- Ilyas, M.; Avelar, S.; Schuch, U.K.; Brown, J.K. First Report of an Emaravirus Associated with Witches’ Broom Disease and Eriophyid Mite Infestations of the Blue Palo Verde Tree in Arizona. Plant Dis. 2018, 102, 1863. [Google Scholar] [CrossRef]
- Yang, C.; Zhang, S.; Han, T.; Fu, J.; Di Serio, F.; Cao, M. Identification and Characterization of a novel Emaravirus associated with Jujube (Ziziphus jujuba Mill.) Yellow Mottle Disease. Front. Microbiol. 2019, 10, 1417. [Google Scholar] [CrossRef] [Green Version]
- Olmedo-Velarde, A.; Park, A.C.; Sugano, J.; Uchida, J.Y.; Kawate, M.; Borth, W.B.; Hu, J.S.; Melzer, M.J. Characterization of Ti Ringspot-Associated Virus, a Novel Emaravirus Associated with an Emerging Ringspot Disease of Cordyline fruticosa. Plant Dis. 2019, 103, 2345–2352. [Google Scholar] [CrossRef]
- Elbeaino, T.; Digiaro, M.; Mielke-Ehret, N.; Muehlbach, H.P.; Martelli, G.P. ICTV Report Consortium. ICTV virus taxonomy profile: Fimoviridae. J. Gen. Virol. 2018, 99, 1478–1479. [Google Scholar] [CrossRef]
- Bostock, R.M.; Pye, M.F.; Roubtsova, T.V. Predisposition in plant disease: Exploiting the nexus in abiotic and biotic stress perception and response. Annu. Rev. Phytopathol. 2014, 52, 517–549. [Google Scholar] [CrossRef] [Green Version]
- Bao, X.; Roossinck, M.J. A life history view of mutualistic viral symbioses: Quantity or quality for cooperation? Curr. Opin. Microbiol. 2013, 16, 514–518. [Google Scholar] [CrossRef]
- van Molken, T.; de Caluwe, H.; Hordijk, C.A.; Leon-Reyes, A.; Snoeren, T.A.; van Dam, N.M.; Stuefer, J.F. Virus infection decreases the attractiveness of white clover plants for a non-vectoring herbivore. Oecologia 2012, 170, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, L.R.; Salvaudon, L.; Mauck, K.E.; Pulido, H.; De Moraes, C.M.; Stephenson, A.G.; Mescher, M.C. Disease interactions in a shared host plant: Effects of pre-existing viral infection on cucurbit plant defense responses and resistance to bacterial wilt disease. PLoS ONE 2013, 8, e77393. [Google Scholar] [CrossRef] [Green Version]
- Pechinger, K.; Chooi, K.M.; MacDiarmid, R.M.; Harper, S.J.; Ziebell, H. A new era for mild strain cross-protection. Viruses 2019, 11, 670. [Google Scholar] [CrossRef] [Green Version]
- Xu, P.; Chen, F.; Mannas, J.P.; Feldman, T.; Sumner, L.W.; Roossinck, M.J. Virus infection improves drought tolerance. New Phytol. 2008, 180, 911–921. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.S.; Bosque-Pérez, N.A.; Foote, N.E.; Magney, T.; Eigenbrode, S.D. Environmentally dependent host–pathogen and vector–pathogen interactions in the Barley yellow dwarf virus pathosystem. J. Appl. Ecol. 2015, 52, 1392–1401. [Google Scholar] [CrossRef]
- Silva, C.C.; Molina, R.O.; Back, L.; Oliveira, C.M.; Frias, A.A.; Zanutto, C.A.; Nunes, W.M. The effect of drought conditions on sweet orange (Citrus sinensis) plants infected with citrus tristeza virus (CTV). Trop. Plant Pathol. 2019, 44, 335–342. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree | RNA Genomic Segment | Contig Number | Size (nt) | Best Match in BlastX (Accession Number) | Query Coverage (%) | E Value | Sequence Identity (%) |
|---|---|---|---|---|---|---|---|
| E53309 | RNA1 | C1727 | 2314 | TiRSaV * (QAB47307.1) | 99 | 1 × 10−139 | 56 |
| C447 | 1290 | TiRSaV (QAB47307.1) | 99 | 0.0 | 48 | ||
| C329 | 1509 | TiRSaV (QAB47307.1) | 84 | 2 × 10−90 | 42 | ||
| RNA2 | C1291 | 2295 | TiRSaV (QAB47308.1) | 76 | 0.0 | 44 | |
| RNA3 | C881 | 1170 | TiRSaV (QAB47309.1) | 71 | 2 × 10−41 | 39 | |
| RNA4 | C755 | 1388 | TiRSaV (QAB47310.1) | 71 | 2 × 10−158 | 65 | |
| E54899 | RNA1 | S71 | 527 | TiRSaV (QAB47307.1) | 88 | 8 × 10−18 | 33 |
| S134 | 346 | RLBV ** (YP_009237274.1) | 97 | 4 × 10−28 | 62 | ||
| S36 | 805 | RLBV (YP_009237274.1) | 90 | 1 × 10−39 | 40 | ||
| RNA2 | S18 | 1138 | TiRSaV (QAB47308.1) | 89 | 4 × 10−105 | 42 | |
| RNA3 | S28 | 990 | TiRSaV (QAB47309.1) | 75 | 2 × 10−47 | 41 | |
| RNA4 | S26 | 1008 | TiRSaV (QAB47310.1) | 64 | 2 × 10−104 | 70 |
| Tree Year | Chlorotic Ringspots [No. Detected/No. Tested] | No Symptoms [No. Detected/no. Tested] |
|---|---|---|
| 2016 | 40/40 | 0/4 |
| 2017 | 20/20 | 0/0 |
| 2018 | 52/53 | 0/0 |
| Total (no.) | 112/113 | 0/4 |
| Total (%) | 99,1 | 0 |
| 1992 | 1998 | 2003 | 2008 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Groups | total | 186 | 175 | 171 | 164 | 162 | 162 | 162 | 162 | 162 | 162 |
| with characteristic symptoms | ne | 14 | 24 | 34 | 17 | 19 | 33 | 38 | 33 | 42 | |
| Seedlings | total | 1302 | 577 | 455 | 440 | 435 | 432 | 432 | 427 | 425 | 422 |
| with characteristic symptoms | ne | 19 | 28 | 43 | 17 | 21 | 38 | 42 | 39 | 51 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bandte, M.; Rehanek, M.; Leder, B.; von Bargen, S.; Büttner, C. Identification of an Emaravirus in a Common Oak (Quercus robur L.) Conservation Seed Orchard in Germany: Implications for Oak Health. Forests 2020, 11, 1174. https://doi.org/10.3390/f11111174
Bandte M, Rehanek M, Leder B, von Bargen S, Büttner C. Identification of an Emaravirus in a Common Oak (Quercus robur L.) Conservation Seed Orchard in Germany: Implications for Oak Health. Forests. 2020; 11(11):1174. https://doi.org/10.3390/f11111174
Chicago/Turabian StyleBandte, Martina, Marius Rehanek, Bertram Leder, Susanne von Bargen, and Carmen Büttner. 2020. "Identification of an Emaravirus in a Common Oak (Quercus robur L.) Conservation Seed Orchard in Germany: Implications for Oak Health" Forests 11, no. 11: 1174. https://doi.org/10.3390/f11111174
APA StyleBandte, M., Rehanek, M., Leder, B., von Bargen, S., & Büttner, C. (2020). Identification of an Emaravirus in a Common Oak (Quercus robur L.) Conservation Seed Orchard in Germany: Implications for Oak Health. Forests, 11(11), 1174. https://doi.org/10.3390/f11111174