Tree Growth and Wood Quality in Pure Vs. Mixed-Species Stands of European Beech and Calabrian Pine in Mediterranean Mountain Forests


 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Tree Sampling and BAI Analyses
2.3. Wood Quality
2.4. Relationship between Wood Quality and Tree Growth
2.5. Statistical Analysis
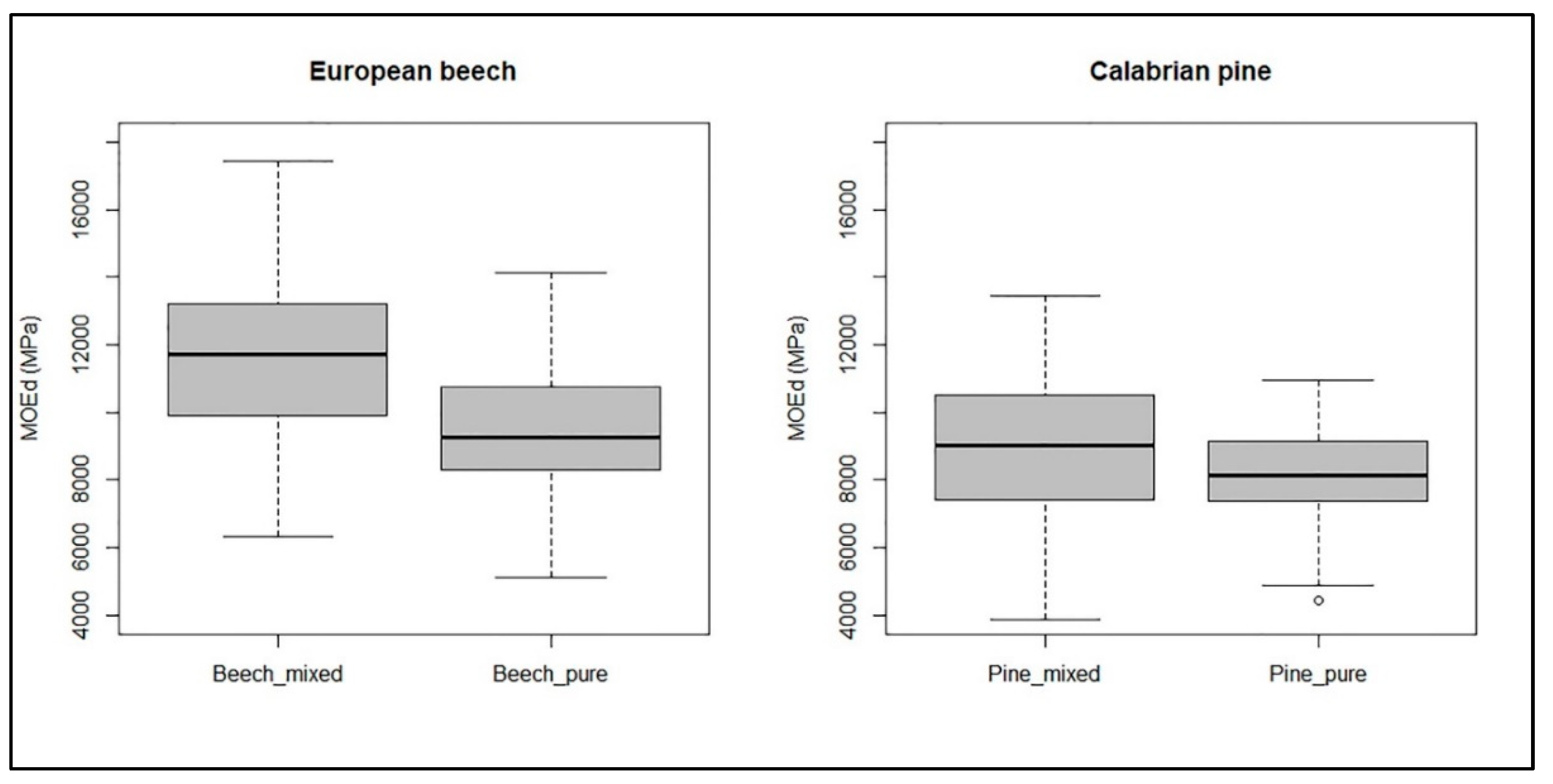
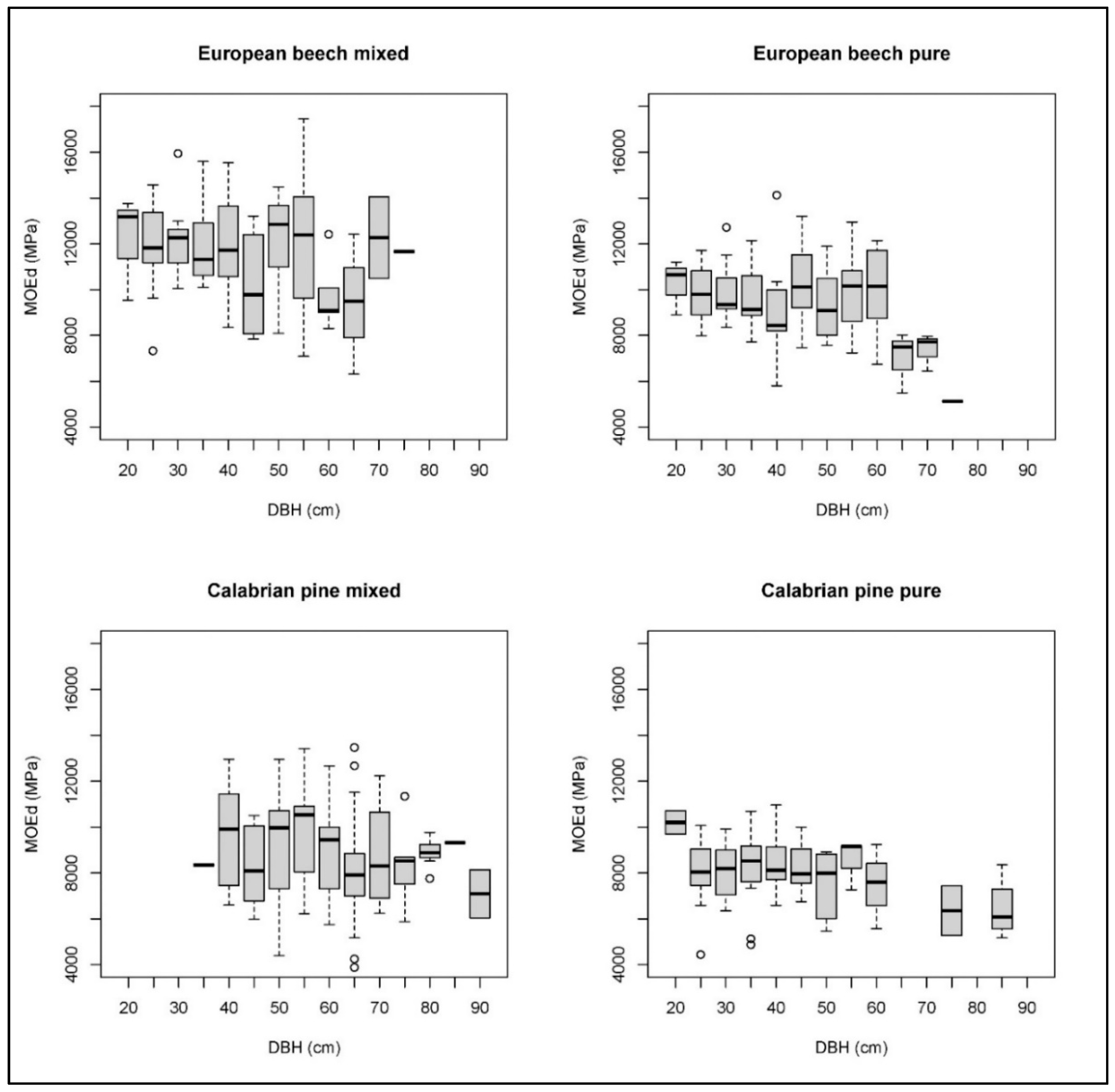
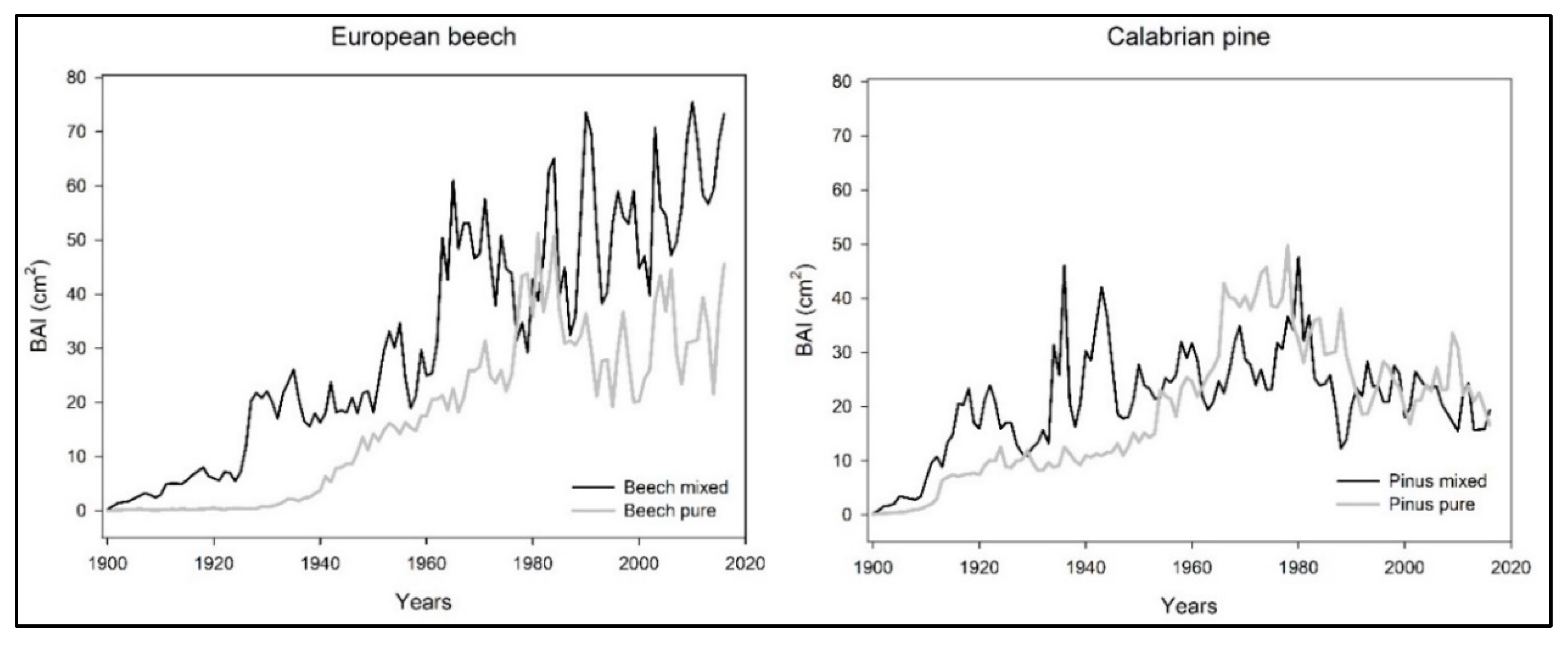
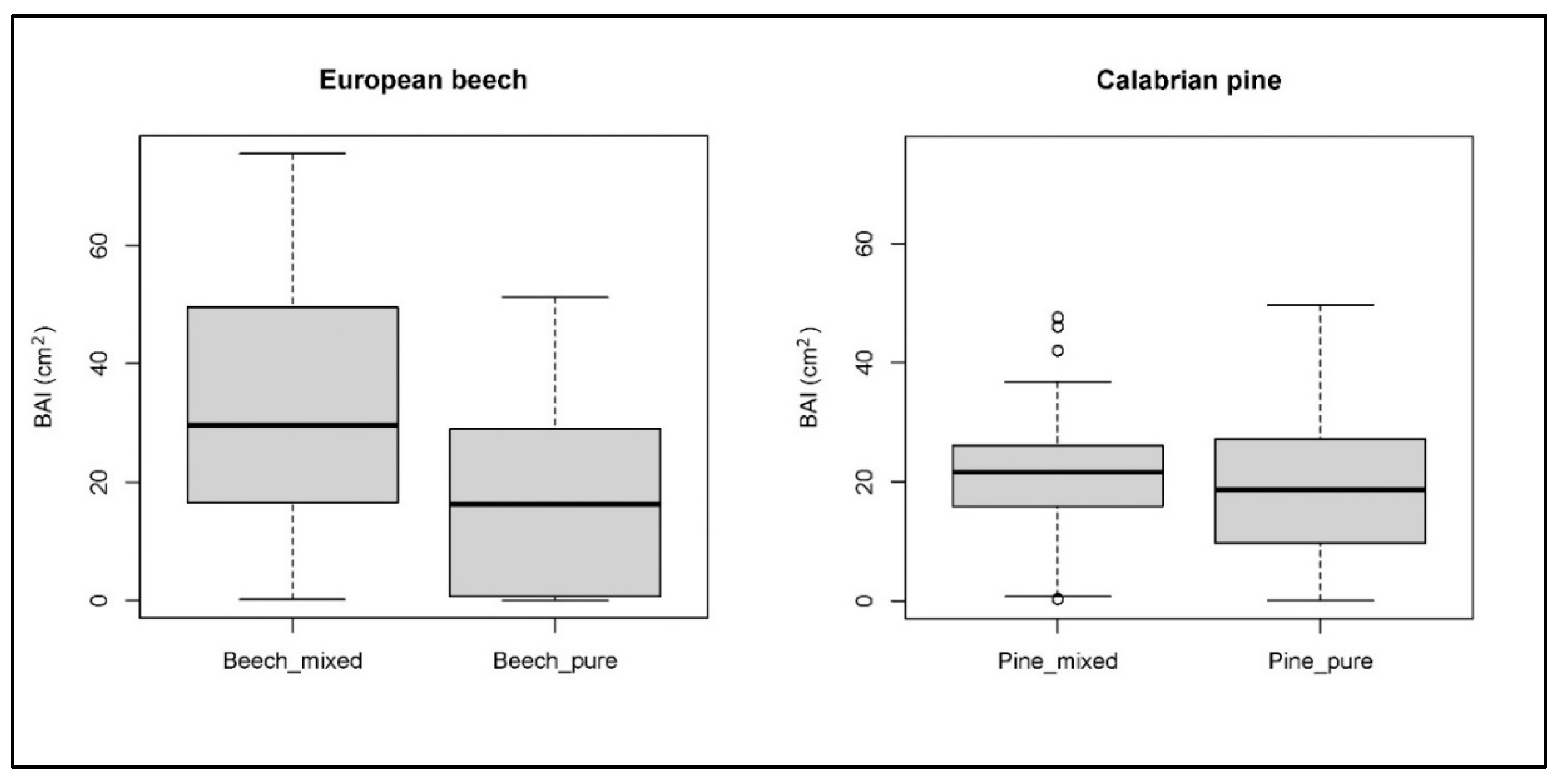
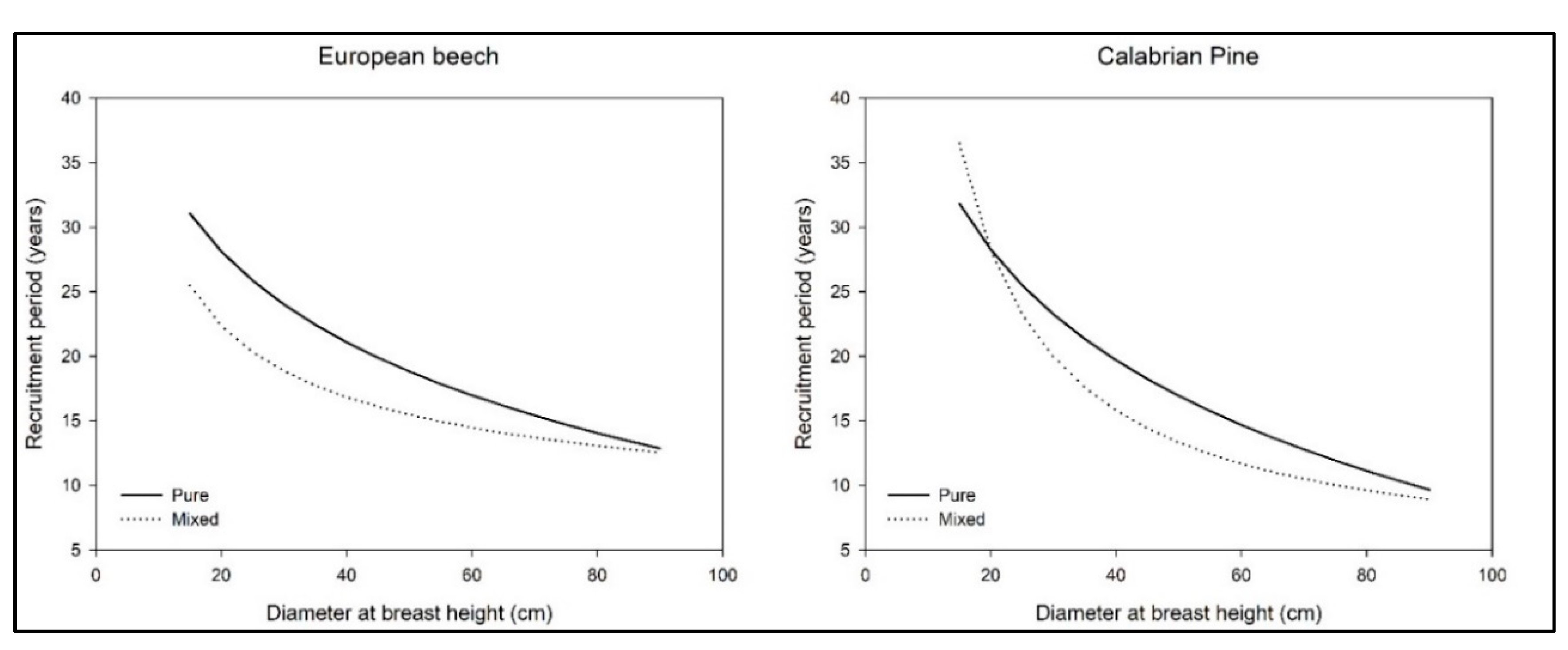
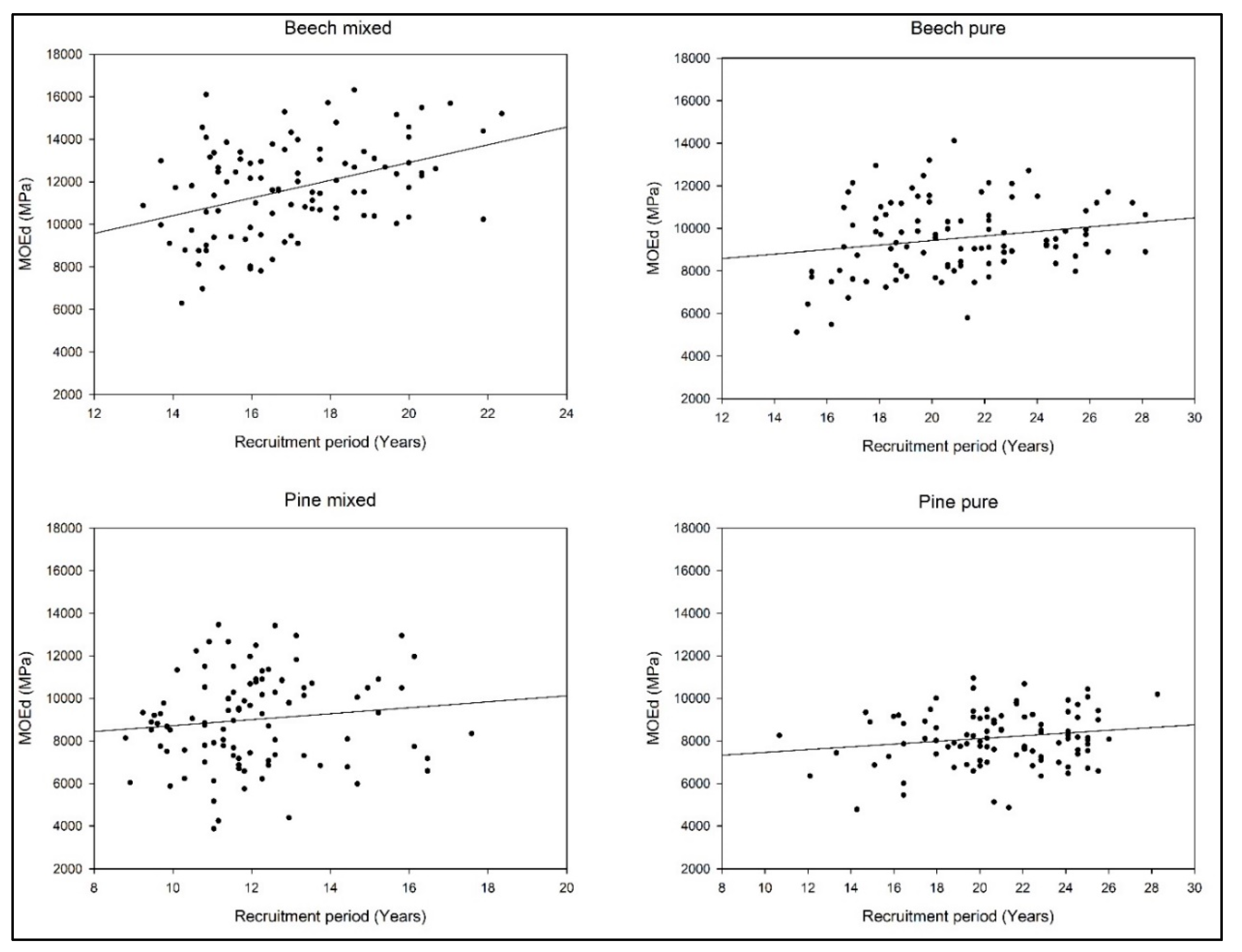
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gamfeldt, L.; Snäll, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P.; Ruiz-Jaen, M.C.; Fröberg, M.; Stendahl, J.; Philipson, C.D.; et al. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun. 2013, 4, 1340. [Google Scholar] [CrossRef] [PubMed]
- van der Plas, F.; Manning, P.; Soliveres, S.; Allan, E.; Scherer-Lorenzen, M.; Verheyen, K.; Wirth, C.; Zavala, M.A.; Ampoorter, E.; Baeten, L.; et al. Biotic homogenization can decrease landscape-scale forest multifunctionality. Proc. Natl. Acad. Sci. USA 2016, 113, 3557–3562. [Google Scholar] [CrossRef] [PubMed]
- Cudlín, P.; Klopčič, M.; Tognetti, R.; Máliš, F.; Alados, C.L.; Bebi, P.; Grunewald, K.; Zhiyanski, M.; Andonowski, V.; La Porta, N.; et al. Frans Emil Wielgolaski Drivers of treeline shift in different European mountains. Clim. Res. 2017, 73, 135–150. [Google Scholar] [CrossRef]
- Pretzsch, H.; Bielak, K.; Bruchwald, A.; Dieler, J.; Dudzinska, M.; Erhart, H.P.; Jensen, A.M.; Johannsen, V.K.; Kohnle, U.; Nagel, J.; et al. Mischung und Produktivität von Waldbeständen. Ergebnisse langfristiger ertragskundlicher Versuche. Allgemeine Forst-und Jagdzeitung 2013, 184, 177–196. [Google Scholar]
- Yachi, S.; Loreau, M. Does complementary resource use enhance ecosystem functioning? A model of light competition in plant communities. Ecol. Lett. 2007, 10, 54–62. [Google Scholar] [CrossRef]
- Piotto, D. A meta-analysis comparing tree growth in monocultures and mixed plantations. For. Ecol. Manag. 2008, 255, 781–786. [Google Scholar] [CrossRef]
- Morin, X.; Fahse, L.; Scherer-Lorenzen, M.; Bugmann, H. Tree species richness promotes productivity in temperate forests through strong complementarity between species. Ecol. Lett. 2011, 14, 1211–1219. [Google Scholar] [CrossRef]
- Paquette, A.; Messier, C. The effect of biodiversity on tree productivity: From temperate to boreal forests. Glob. Ecol. Biogeogr. 2011, 20, 170–180. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G.; Uhl, E. Resistance of European tree species to drought stress in mixed versus pure forests: Evidence of stress release by inter-specific facilitation. Plant Biol. 2013, 15, 483–495. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G. Effect of tree species mixing on the size structure, density and yield forest stands. Eur. J. For. Res. 2015, 135, 1–22. [Google Scholar] [CrossRef]
- Jucker, T.; Bouriaud, O.; Avacaritei, D.; Coomes, D.A. Stabilizing effects of diversity on aboveground wood production in forest ecosystems: Linking patterns and processes. Ecol. Lett. 2014, 17, 1560–1569. [Google Scholar] [CrossRef] [PubMed]
- Metz, J.; Annighofer, P.; Schall, P.; Zimmermann, J.; Kahl, T.; Schulze, E.D. Site-adapted admixed tree species reduce drought susceptibility of mature European beech. Glob. Chang. Biol. 2016, 22, 903–920. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Crowther, T.W.; Picard, N.; Wiser, S.; Zhou, M.; Alberti, G.; Schulze, E.D.; McGuire, A.D.; Bozzato, F.; Pretzsch, H.; et al. Positive biodiversity–productivity relationship predominant in global forests. Science 2016, 354, 196. [Google Scholar] [CrossRef] [PubMed]
- Grossiord, C.; Granier, A.; Gessler, A.; Jucker, T.; Bonal, D. Does drought influence the relationship between biodiversity and ecosystem functioning in boreal forests? Ecosystems 2014, 17, 394–404. [Google Scholar] [CrossRef]
- Merlin, M.; Perot, T.; Perret, S.; Korboulewsky, N.; Vallet, P. Effects of stand composition and tree size on resistance and resilience to drought in sessile oak and Scots pine. For. Ecol. Manag. 2015, 339, 22–33. [Google Scholar] [CrossRef]
- Conte, E.; Lombardi, F.; Battipaglia, G.; Palombo, C.; Altieri, S.; La Porta, N.; Marchetti, M.; Tognetti, R. Growth dynamics, climate sensitivity and water use efficiency in pure vs. mixed pine and beech stands in Trentino (Italy). For. Ecol. Manag. 2018, 409, 707–718. [Google Scholar] [CrossRef]
- Gebauer, T.; Horna, V.; Leuschner, C. Canopy transpiration of pure and mixed forest stands with variable abundance of European beech. J. Hydrol. 2012, 442, 2–14. [Google Scholar] [CrossRef]
- IPCC. Meeting report of the intergovernmental panel on climate change expert meeting on mitigation, sustainability and climate stabilization scenarios. In IPCC Working Group III Technical Support Unit; Shukla, P.R.J., Skea, R., van Diemen, K., Calvin, Ø., Christophersen, F., Creutzig, J., Eds.; Imperial College London: London, UK, 2017. [Google Scholar]
- Hanewinkel, M.; Cullmann, D.; Schelhaas, M.J.; Nabuurs, G.J.; Zimmermann, N.E. Climate change may cause severe loss in economic value of European forestland. Nat. Clim. Chang. 2013, 3, 204–207. [Google Scholar] [CrossRef]
- Altieri, V.; De Franco, S.; Lombardi, F.; Marziliano, P.A.; Menguzzato, G.; Porto, P. The role of silvicultural systems and forest types in preventing soil erosion processes in mountain forests: A methodological approach using cesium-137 measurements. J. Soils Sediments 2018, 18, 1–10. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.Y.H.; Reich, P.B. Forest productivity increases with evenness, species richness and trait variation: A global meta-analysis. J. Ecol. 2012, 100, 742–749. [Google Scholar] [CrossRef]
- Kanowski, J.; Catterall, C.P.; Wardell-Johnson, G.W. Consequences of broadscale timber plantations for biodiversity in cleared rainforest landscapes of tropical and subtropical Australia. For. Ecol. Manag. 2005, 208, 359–372. [Google Scholar] [CrossRef]
- Petit, B.; Montagnini, F. Growth in pure and mixed plantations of tree species used in reforesting rural areas of the humid region of Costa Rica, Central America. For. Ecol. Manag. 2006, 233, 338–343. [Google Scholar] [CrossRef]
- Richards, A.E.; Forrester, D.I.; Bauhus, J.; Scherer-Lorenzen, M. The influence of mixed tree plantations on the nutrition of individual species: A review. Tree Physiol. 2010, 30, 1192–1208. [Google Scholar] [CrossRef]
- Bielak, K.; Dudzinska, M.; Pretzsch, H. Mixed stands of Scots pine (Pinus sylvestris L.) and Norway spruce [Picea abies (L.) Karst] can be more productive than monocultures. Evidence from over 100 years of observation of long-term experiments. For. Syst. 2014, 23, 573. [Google Scholar] [CrossRef]
- Forrester, D.I. The spatial and temporal dynamics of species interactions in mixed-species forests: From pattern to process. For. Ecol. Manag. 2014, 312, 282–292. [Google Scholar] [CrossRef]
- Petráš, R.; Mecko, J.; Bošelȃ, M.; Šebeň, V. Wood quality and value production in mixed fir-spruce-beech stands: Long-term research in the Western Carpathians. For. J. 2016, 62, 98–104. [Google Scholar] [CrossRef]
- Magin, R. Ertragskundliche Untersuchungen in montanen Mischwäldern. Forstwissenschaftliches Centralblatt 1954, 73, 103–113. [Google Scholar] [CrossRef]
- Kennel, R. Untersuchungen über die Leistung von Fichte und Buche im Reinund Mischbestand. Sauerländer 1965, 136, 149–161. [Google Scholar]
- Kennel, R. Soziale Stellung, Nachbarschaft und Zuwachs. Forstwissenschaftliches Centralblatt 1966, 85, 193–204. [Google Scholar] [CrossRef]
- Hausser, K.; Troeger, R. Beitrag zur Frage der Massen und Wertleistung gepflanzter Weisstannen und Fichtenbestände auf gleichen Standorten. Allgemeine Forst-und Jagdzeitung 1967, 138, 150–157. [Google Scholar]
- Mitscherlich, G. Ertragskundlich-ökologische Untersuchungen im Rein-und Mischbestand. Mitteilungen der Forstlichen Bundes-Versuchsanstalt 1967, 77, 9–35. [Google Scholar]
- Hink, V. Das Wachstum von Fichte und Tanne auf den wichtigsten Standortseinheiten des Einzelwuchsbezirks “Flächenschwarzwald” (Südwürtemberg-Hohenzollern). Allgemeine Forst-und Jagdzeitung 1972, 143, 80–85. [Google Scholar]
- Mettin, C. Betriebswirtschaftliche und ökologische Zusammenhänge zwischen Standortskraft und Leistung in Fichtenreinbeständen und Fichten Buchen-Mischbeständen. Allgemeine Forst-und Jagdzeitung 1985, 40, 803–810. [Google Scholar]
- Kramer, H. Waldwachstumslehre (Forest Growth and Yield Science); Paul Parey: Hamburg, Germany, 1988; p. 374. (In German) [Google Scholar]
- Pretzsch, H. Forest Dynamics, Growth and Yield: From Measurement to Model; Springer: Berlin, Germany, 2010. [Google Scholar]
- Saha, S.; Kuehne, C.; Kohnle, U.; Brang, P.; Ehring, A.; Geisel, J.; Leder, B.; Muth, M.; Petersen, R.; Peter, J.; et al. Growth and quality of young oaks (Quercus robur and Quercus petraea) grown in cluster plantings in central Europe: A weighted meta-analysis. For. Ecol. Manag. 2012, 283, 106–118. [Google Scholar] [CrossRef]
- Liu, C.L.C.; Kuchma, O.; Krutovsky, K.V. Mixed secies versus monocultures in plantation forestry: Development, benefits, ecosystem services and perspectives for the future. Glob. Ecol. Conserv. 2018, 15, e00419. [Google Scholar] [CrossRef]
- Chomel, M.; DesRochers, A.; Baldy, V.; Larchevȇque, M.; Gauquelin, T. Non-additive effects of mixing hybrid poplar and white spruce on aboveground and soil carbon storage in boreal plantations. For. Ecol. Manag. 2014, 328, 292–299. [Google Scholar] [CrossRef]
- Battipaglia, G.; Pelleri, F.; Lombardi, F.; Altieri, S.; Vitone, A.; Conte, E.; Tognetti, R. Effects of associating Quercus robur L. and Alnus cordata Loisel. on plantation productivity and water use efficiency. For. Ecol. Manag. 2017, 391, 106–114. [Google Scholar] [CrossRef]
- Kelty, M.J. The role of species mixtures in plantation forestry. For. Ecol. Manag. 2006, 233, 195–204. [Google Scholar] [CrossRef]
- Machado, J.S.; Louzada, J.L.; Santos, A.J.A.; Nunes, L.; Anjos, O.; Rodrigues, J.; Simoes, R.M.S.; Pereira, H. Variation of wood density and mechanical properties of blackwood (Acacia melanoxylon r. Br.). Mater. Des. 2014, 56, 975–980. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G. Transgressive overyielding in mixed compared with pure stands of Norway spruce and European beech in Central Europe: Evidence on stand level and explanation on individual tree level. Eur. J. For. Res. 2009, 128, 183–204. [Google Scholar] [CrossRef]
- Russo, D.; Marziliano, P.A.; Macri, G.; Proto, A.R.; Zimbalatti, G.; Lombardi, F. Does thinning intensity affect wood quality? An analysis of Calabrian Pine in Southern Italy using a non-destructive acoustic method. Forests 2019, 30, 303. [Google Scholar] [CrossRef]
- Guntekin, G.; Emiroglu, Z.G.; Yolmaz, T. Prediction of bending properties for Turkish red pine (Pinus brutia Ten.) Lumber using stress wave method. Bioresources 2013, 8, 231–237. [Google Scholar] [CrossRef]
- Teder, M.; Pilt, K.; Miljan, M.; Lainurm, M.; Kruuda, R. Overview of some non-destructive methods for in-situ assessment of structural timber. In Proceedings of the 3rd International Conference Civil Engineering, Jelgava, Latvia, 12–13 May 2011. [Google Scholar]
- Wessels, C.B.; Malan, F.S.; Rypstra, T. A review of measurement methods used on standing trees for the prediction of some mechanical properties of timber. Eur. J. For. Res. 2011, 130, 881–893. [Google Scholar] [CrossRef]
- Bacher, M.; Krosek, S. Bending and tension strength classes in European standards. Annals of Warsaw University of Life Sciences—SGGW. For. Wood Tech. 2014, 88, 14–22. [Google Scholar]
- Pretzsch, H.; Rais, A. Wood quality in complex forests versus even-aged monocultures: Review and perspectives. Wood Sci. Technol. 2016, 50, 845–880. [Google Scholar] [CrossRef]
- Rais, A.; Van de Kuilen, J.W.G. Critical section effect during derivation of settings for grading machines based on dynamic modulus of elasticity. Wood Mater. Sci. Eng. 2015, 4, 189–196. [Google Scholar] [CrossRef]
- FAO. World Reference Base for Soil Resources. In International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports, No. 106; FAO: Roma, Italy, 2014. [Google Scholar]
- Ciancio, O.; Iovino, F.; Menguzzato, G.; Nicolaci, A.; Nocentini, S. Structure and growth of a small group selection forest of Calabrian pine in Southern Italy: A hypothesis for continuous cover forestry based on traditional silviculture. For. Ecol. Manag. 2006, 224, 229–234. [Google Scholar] [CrossRef]
- Tabacchi, G.; Di Cosmo, L.; Gasparini, P. Aboveground tree volume and phytomass prediction equations for forest species in Italy. Eur. J. For. Res. 2011, 130, 911–934. [Google Scholar] [CrossRef]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Wang, X. Acoustic measurements on trees and logs: A review and analysis. Wood Sci. Technol. 2013, 47, 965–975. [Google Scholar] [CrossRef]
- Vanninen, P.; Ylitalo, H.; Sievanen, R.; Makela, A. Effect of age and site quality on the distribution of biomass in Scots pine (Pinus sylvestris L.). Trees 1996, 10, 231–238. [Google Scholar] [CrossRef]
- Vargas-Hernandez, J.; Adams, W.T. Genetic relationships between wood density components and cambial growth rhythm in young coastal Douglas-fir. Can. J. For. Res. 1994, 24, 1871–1876. [Google Scholar] [CrossRef]
- Zhembo, L.; Yixing, L.; Haipeng, Y.; Junqi, Y. Measurement of the dynamic modulus of elasticity of wood panels. Front. For. China 2006, 1, 425–430. [Google Scholar]
- Todaro, L.; Macchioni, N. Wood properties of young Douglas-fir in Southern Italy: Results over a 12-year post-thinning period. Eur. J. For. Res. 2011, 130, 251–261. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, X.; Su, J. Experimental Investigation of Stress Wave Propagation in Standing Trees. Holzforschung 2011, 65, 743–748. [Google Scholar] [CrossRef]
- Krajnc, L.; Farrelly, N.; Harte, A.M. Evaluating Timber Quality in Larger-Diameter Standing Trees: Rethinking the Use of Acoustic Velocity. Holzforschung 2019, 73, 797–806. [Google Scholar] [CrossRef]
- Marziliano, P.A.; Menguzzato, G.; Scuderi, A.; Corona, P. Simplified methods to inventory the current annual increment of forest standing volume. iForest 2012, 5, 276–282. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 4th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 1999. [Google Scholar]
- Detters, A.; Cowell, C.; McKeown, L.; Howard, P. Evaluation of Current Rigging and Dismantling Practices Used in Arboriculture; The Health and Safety Executive: London, UK; The Forestry Commission: Edinburgh, UK, 2008; p. 355. [Google Scholar]
- Binkley, D.; Dunkin, K.A.; DeBell, D.; Ryan, M.G. Production and nutrient cycling in mixed plantations of eucalyptus and albizia in Hawaii. For. Sci. 1992, 38, 393–408. [Google Scholar]
- Kelty, M.J. Comparative productivity of monocultures and mixed-species stands. In The Ecology and Silviculture of Mixed-Species Forests; Springer: Dordrecht, The Netherlands, 1992; pp. 125–141. [Google Scholar]
- Forrester, D.I.; Lancaster, K.; Collopy, J.J.; Warren, C.R.; Tausz, M. Photosynthetic capacity of Eucalyptus globulus is higher when grown in mixture with Acacia mearnsii. Trees 2012, 26, 1203–1213. [Google Scholar] [CrossRef]
- Saranpää, P. Wood density and growth. In Wood Quality and Its Biological Basis Blackwell; Biological Sciences Series; Barnett, J.R., Jeronimidis, G., Eds.; CRC Press: Oxford, FL, USA, 2003; pp. 87–113. [Google Scholar]
- Torquato, L.P.; Auty, D.; Herȃndez, R.E.; Duchesne, I.; Pothier, D.; Achim, A. Black spruce trees from fire-origin stands have higher wood mechanical properties than those from older, irregular stands. Can. J. For. Res. 2014, 44, 118–127. [Google Scholar] [CrossRef]
- Metz, J.Ô.; Seidel, D.; Schall, P.; Scheffer, D.; Schulze, E.D.; Ammer, C. Crown modeling by terrestrial laser scanning as an approach to assess the effect of aboveground intra- and interspecific competition on tree growth. For. Ecol. Manag. 2013, 310, 275–288. [Google Scholar] [CrossRef]
- Pretzsch, H.; Biber, P. A re-evaluation of Reineke’s rule and stand density index. For. Sci. 2005, 51, 304–320. [Google Scholar]
- Pretzsch, H. Species-specific allometric scaling under self-thinning: Evidence from long-term plots in forest stands. Oecologia 2006, 146, 572–583. [Google Scholar] [CrossRef] [PubMed]
- Pretzsch, H.; Block, J.; Dieler, J.; Dong, P.H.; Kohnle, U.; Nagel, J.; Spellmann, H.; Zingg, A. Comparison between the productivity of pure and mixed stands of Norway spruce and European beech along an ecological gradient. Ann. For. Sci. 2010, 67, 712. [Google Scholar] [CrossRef]
- Pretzsch, H.; Bielak, K.; Block, J.; Bruchwald, A.; Dieler, J.; Ehrhart, H.P.; Kohnle, U.; Nagel, J.; Spellmann, H.; Zasada, M.; et al. Productivity of mixed versus pure stands of oak (Quercus petraea (Matt.) Liebl. and Quercus robur L.) and European beech (Fagus sylvatica L.) along an ecological gradient. Eur. J. For. Res. 2013, 132, 263–280. [Google Scholar] [CrossRef]
- Condés, S.; del Río, M.; Sterba, H. Mixing effect on volume growth of Fagus sylvatica and Pinus sylvestris is modulated by stand density. For. Ecol. Manag. 2013, 292, 86–95. [Google Scholar] [CrossRef]
- Zeller, L.; Ammer, C.; Annighöfer, P.; Biber, P.; Marshall, J.; Schütze, G.; del Río Gaztelurrutia, M.; Pretzsch, H. Tree ring wood density of Scots pine and European beech lower in mixed-species stands compared with monocultures. For. Ecol. Manag. 2017, 400, 363–374. [Google Scholar] [CrossRef]
- Binkley, D.; Stape, J.L.; Ryan, M.G. Thinking about efficiency of resource use in forests. For. Ecol. Manag. 2004, 193, 5–16. [Google Scholar] [CrossRef]
- Lebourgeois, F.; Gomez, N.; Pinto, P.; Mérian, P. Mixed stands reduce Abies alba tree-ring sensitivity to summer drought in the Vosges mountains, western Europe. For. Ecol. Manag. 2013, 303, 61–71. [Google Scholar] [CrossRef]
- Del Río, M.D.; Schütze, G.; Pretzsch, H. Temporal variation of competition and facilitation in mixed species forests in Central Europe. Plant Biol. 2014, 16, 166–176. [Google Scholar] [CrossRef]
- Del Río, M.; Pretzsch, H.; Ruíz-Peinado, R.; Ampoorter, E.; Annighöfer, P.; Barbeito, I.; Bielak, K.; Brazaitis, G.; Coll, L.; Drössler, L. Species interactions increase the temporal stability of community productivity in Pinus sylvestris-Fagus sylvatica mixtures across Europe. J. Ecol. 2017, 105, 1032–1043. [Google Scholar] [CrossRef]
- Ellenberg, H.; Leuschner, C. Vegetation Mitteleuropas Mit den Alpen in Ökologischer, Dynamischer und Historischer Sicht; Eugen Ulmer Stuttgart: Nürtingen, Germany, 2010. [Google Scholar]
- Pretzsch, H. The Effect of Tree Crown Allometry on Community Dynamics in Mixed-Species Stands versus Monocultures. A Review and Perspectives for Modeling and Silvicultural Regulation. Forests 2019, 10, 810. [Google Scholar] [CrossRef]
- Forrester, D.I.; Ammer, C.; Annighöfer, P.J.; Barbeito, I.; Bielak, K.; Bravo-Oviedo, A.; Coll, L.; del Río, M.; Drössler, L.; Heym, M.; et al. Effects of crown architecture and stand structure on light absorption in mixed and monospecific Fagus sylvatica and Pinus sylvestris forests along a productivity and climate gradient through Europe. J. Ecol. 2018, 106, 746–760. [Google Scholar] [CrossRef]
- Genet, A.; Auty, D.; Achim, A.; Bernier, M.; Pothier, D.; Cogliastro, A. Consequences of faster growth for wood density in northern red oak (Quercus rubra Liebl.). Forestry 2012, 86, 99–110. [Google Scholar] [CrossRef]
- Grammel, R. Zusammenhänge zwischen Wachstumsbedingungen und holztechnologischen Eigenschaften der Fichte. For. Cent. 1990, 109, 119–129. [Google Scholar] [CrossRef]
- Brazier, J.D.; Mobbs, I.D. The influence of planting distance on structural wood yields of unthinned Sitka spruce. Forestry 1993, 66, 333–352. [Google Scholar] [CrossRef]
- Larocque, G.R.; Marshall, P. Wood relative density development in red pine (Pinus resinosa Ait.) stands as affected by different initial spacings. For. Sci. 1995, 41, 709–728. [Google Scholar]
- Zhang, S.Y.; Chauret, G.; Swift, D.E.; Duchesne, I. Effects of precommercial thinning on tree growth and lumber quality in a jack pine stand in New Brunswick, Canada. Can. J. For. 2006, 36, 945–952. [Google Scholar] [CrossRef]
- Moore, J.R.; Cown, D.J.; McKinley, R.B.; Sabatia, C.O. Effects of stand density and seedlot on three wood properties of young radiata pine grown at a dry-land site in New Zealand. N. Z. J. For. Sci. 2015, 45, 209. [Google Scholar] [CrossRef]
- Tognetti, R.; Michelozzi, M.; Borghetti, M. The response of European beech (Fagus sylvatica L.) seedlings from two Italian populations to drought and recovery. Trees 1995, 9, 348–354. [Google Scholar] [CrossRef]
- Bolte, A.; Czajkowski, T.; Cocozza, C.; Tognetti, R.; de Miguel, M.; Pšidová, E.; Ditmarová, Ĺ.; Dinca, L.; Delzon, S.; Cochard, H.; et al. Desiccation and mortality dynamics in seedlings of different European beech (Fagus sylvatica L.) populations under extreme drought conditions. Front. Plant Sci. 2016, 7, 751. [Google Scholar] [CrossRef] [PubMed]
- Cocozza, C.; de Miguel, M.; Pšidová, E.; Ditmarová, L.; Marino, S.; Maiuro, L.; Alvino, A.; Czajkowski, T.; Bolte, A.; Tognetti, R. Variation in ecophysiological traits and drought tolerance of beech (Fagus sylvatica L.) seedlings from different populations. Front. Plant Sci. 2016, 7, 886. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Triplet 1 | Triplet 2 | Triplet 3 | |
|---|---|---|---|
| Latitude (N) | 38°06′50.53″ | 38°05′54.59″ | 38°06′15.69″ |
| Longitude (E) | 15°51′53.25″ | 15°50′37.72″ | 15°51′23.37″ |
| Exposure (°) | 59 | 59 | 59 |
| Altitude (m a.s.l.) | 1521 | 1511 | 1497 |
| Slope (°) | 28 | 30 | 34 |
| Stand Density (N. Trees ha−1) | Tree DBH (cm) | Tree Height (m) | Basal Area (m2 h−1) | Stand Volume (m3 ha−1) | |
|---|---|---|---|---|---|
| Beech pure | 743 (155.7) | 33.2 (2.8) | 24.2 (2.2) | 64.3 (4.6) | 772.7 (41.5) |
| Pine pure | 754 (96.3) | 35.6 (4.4) | 23.1 (0.8) | 75.1 (10.4) | 857.6 (160.9) |
| MBP-B | 470 (75.9) | 28.3 (2.3) | 18.8 (1.3) | 29.6 (6.3) | 277.0 (31.1) |
| MBP-P | 403 (74.1) | 40.3 (9.6) | 23.6 (5.1) | 51.4 (14.1) | 599.1 (90.7) |
| Stands | Tree Age (Years) | Wood Density (kg m−3) | Wave Velocity (m s−1) | MOEd (MPa) | ||||
|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Beech pure | 69 | 41 | 658 | 28.4 | 3841.7 | 382.4 | 9523.2 | 1738.1 |
| Pine pure | 97 | 38 | 562 | 27.6 | 3823.4 | 427.5 | 8056.6 | 1358.2 |
| MBP-B | 127 | 33 | 653 | 30.1 | 4253.0 | 349.3 | 11583.0 | 2212.6 |
| MBP-P | 103 | 21 | 555 | 25.2 | 4066.4 | 370.6 | 8999.8 | 2132.6 |
| Model | Value | t-Value | p-Value | R2 |
|---|---|---|---|---|
| European beech (BAI) | 0.502 | |||
| Intercept | 17.623 | 7.720 | <0.0001 | |
| Mix (BA) | 0.358 | 4.047 | 0.0155 | |
| Calabrian pine (BAI) | 0.015 | |||
| Intercept | 19.299 | 8.126 | <0.0001 | |
| Mix (BA) | 0.028 | 0.567 | 0.6009 | |
| European beech (MOEd) | 0.278 | |||
| Intercept | 5114.401 | 4.191 | 0.0000 | |
| RP | 210.252 | 4.119 | 0.0001 | |
| Mix (BA) | 76.434 | 3.353 | 0.0285 | |
| Calabrian pine (MOEd) | 0.045 | |||
| Intercept | 6487.034 | 6.291 | <0.0001 | |
| RP | 83.52 | 1.833 | 0.0684 | |
| Mix (BA) | 23.23 | 2.079 | 0.1061 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Russo, D.; Marziliano, P.A.; Macrì, G.; Zimbalatti, G.; Tognetti, R.; Lombardi, F. Tree Growth and Wood Quality in Pure Vs. Mixed-Species Stands of European Beech and Calabrian Pine in Mediterranean Mountain Forests. Forests 2020, 11, 6. https://doi.org/10.3390/f11010006
Russo D, Marziliano PA, Macrì G, Zimbalatti G, Tognetti R, Lombardi F. Tree Growth and Wood Quality in Pure Vs. Mixed-Species Stands of European Beech and Calabrian Pine in Mediterranean Mountain Forests. Forests. 2020; 11(1):6. https://doi.org/10.3390/f11010006
Chicago/Turabian StyleRusso, Diego, Pasquale A. Marziliano, Giorgio Macrì, Giuseppe Zimbalatti, Roberto Tognetti, and Fabio Lombardi. 2020. "Tree Growth and Wood Quality in Pure Vs. Mixed-Species Stands of European Beech and Calabrian Pine in Mediterranean Mountain Forests" Forests 11, no. 1: 6. https://doi.org/10.3390/f11010006
APA StyleRusso, D., Marziliano, P. A., Macrì, G., Zimbalatti, G., Tognetti, R., & Lombardi, F. (2020). Tree Growth and Wood Quality in Pure Vs. Mixed-Species Stands of European Beech and Calabrian Pine in Mediterranean Mountain Forests. Forests, 11(1), 6. https://doi.org/10.3390/f11010006