Ozone Amplifies Water Loss from Mature Trees in the Short Term But Decreases It in the Long Term



Abstract
1. Introduction
2. Materials and Methods
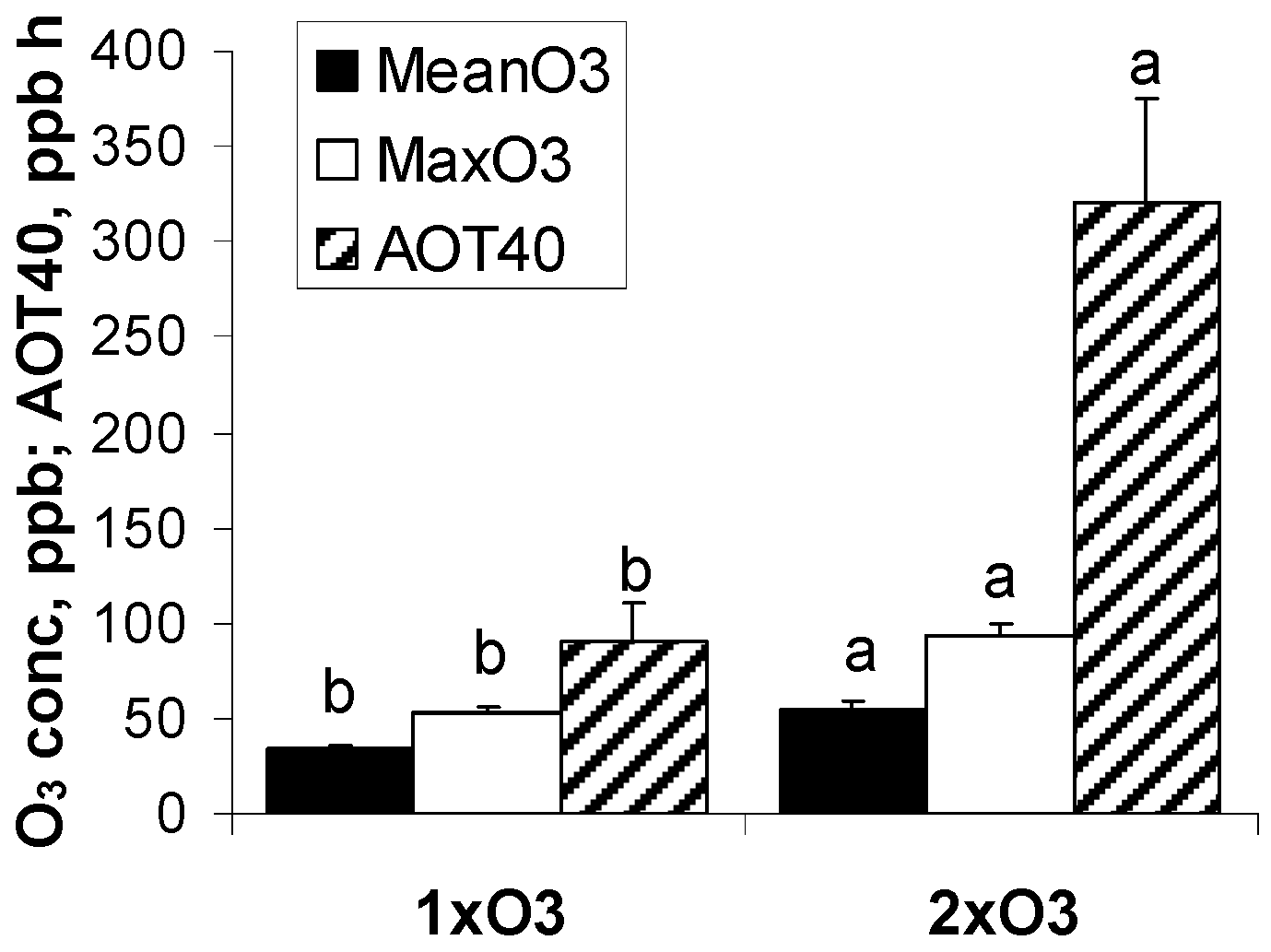
2.1. Experimental Design and Environmental Data
2.2. Assessment of Xylem Sap Flow and Calculation of Whole-Tree Transpiration
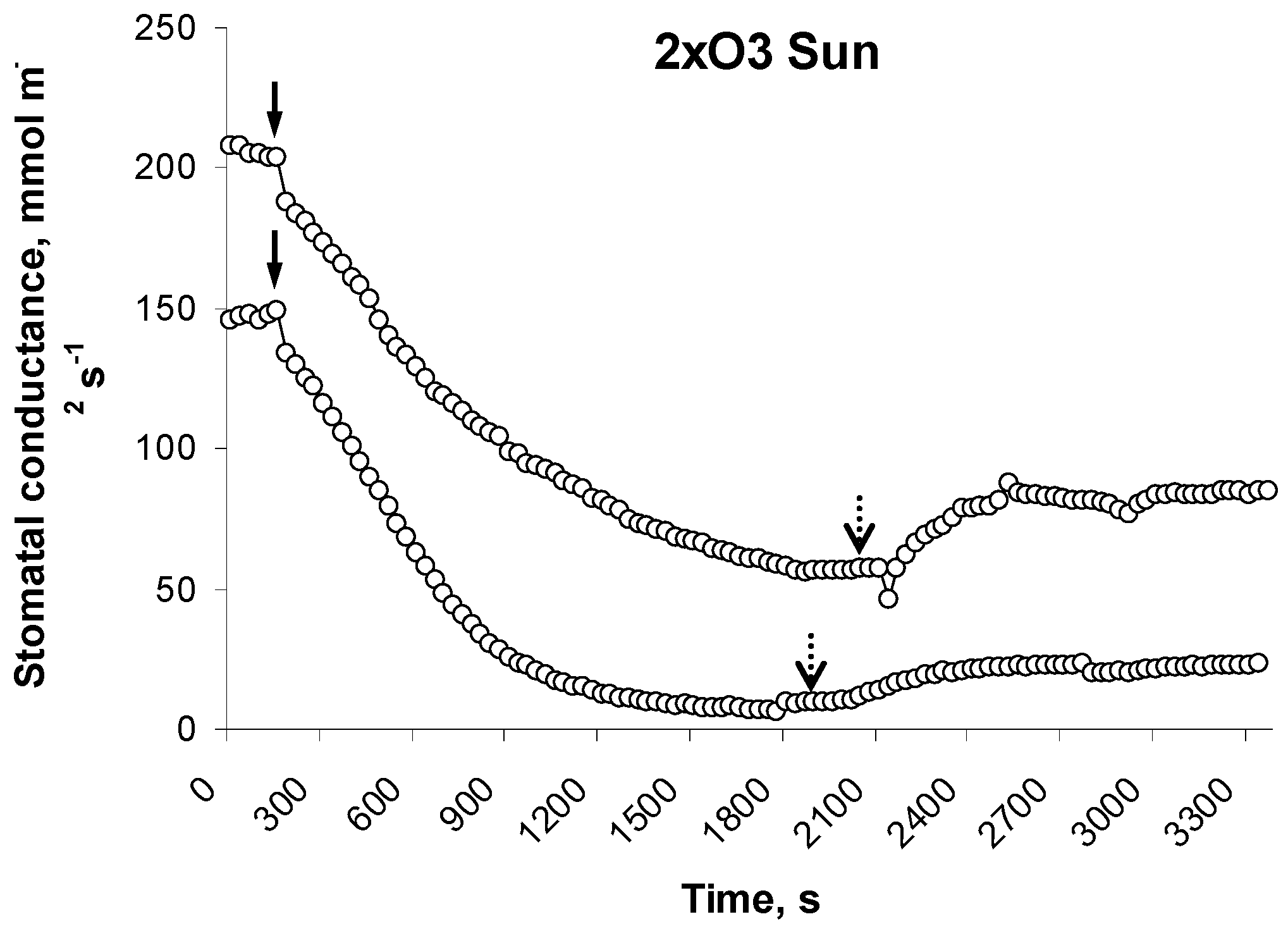
2.3. Assessment of Stomatal Responses to Fleck Light
2.4. Data Analysis
3. Results
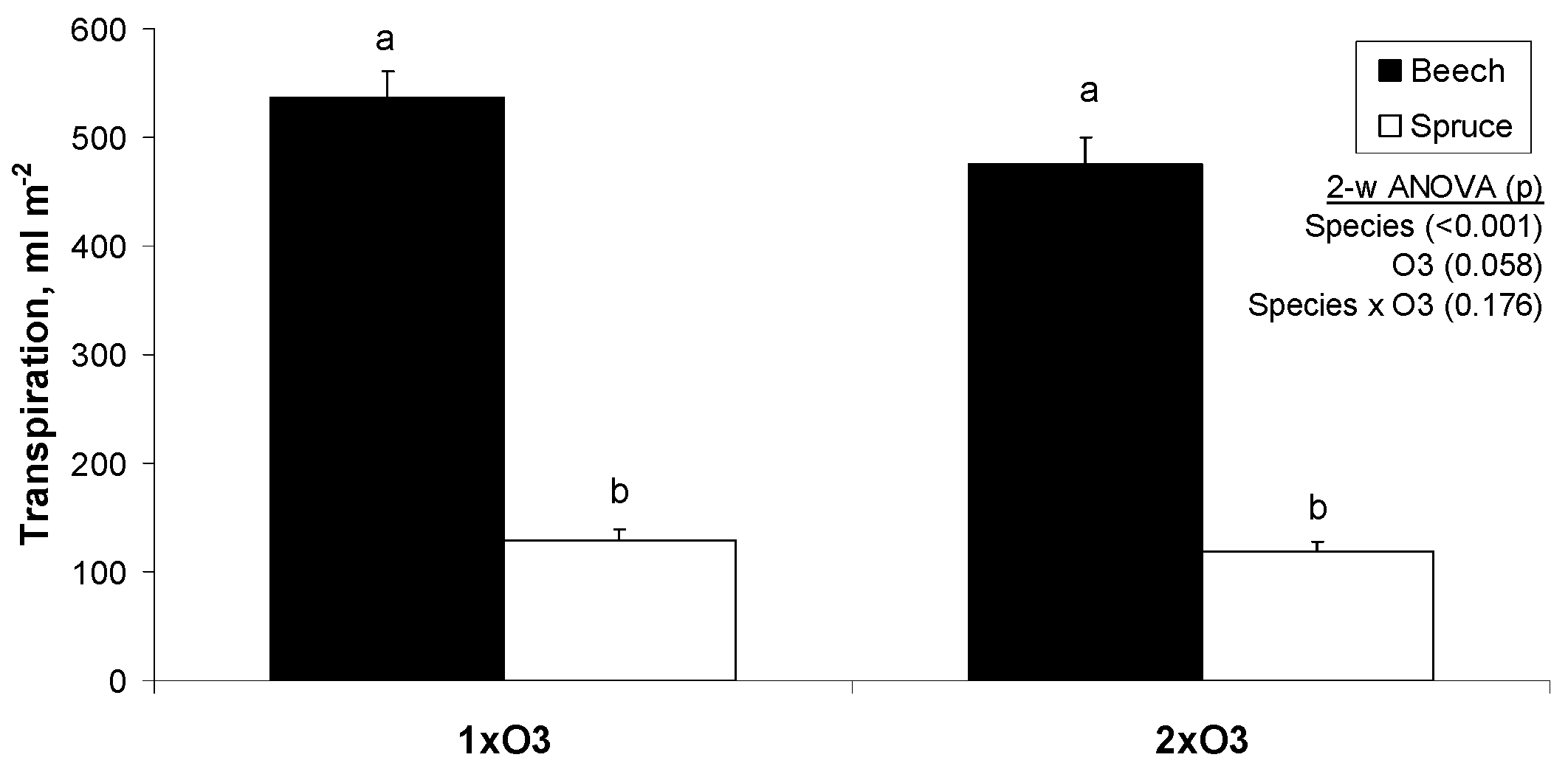
3.1. Tree-Level Transpiration
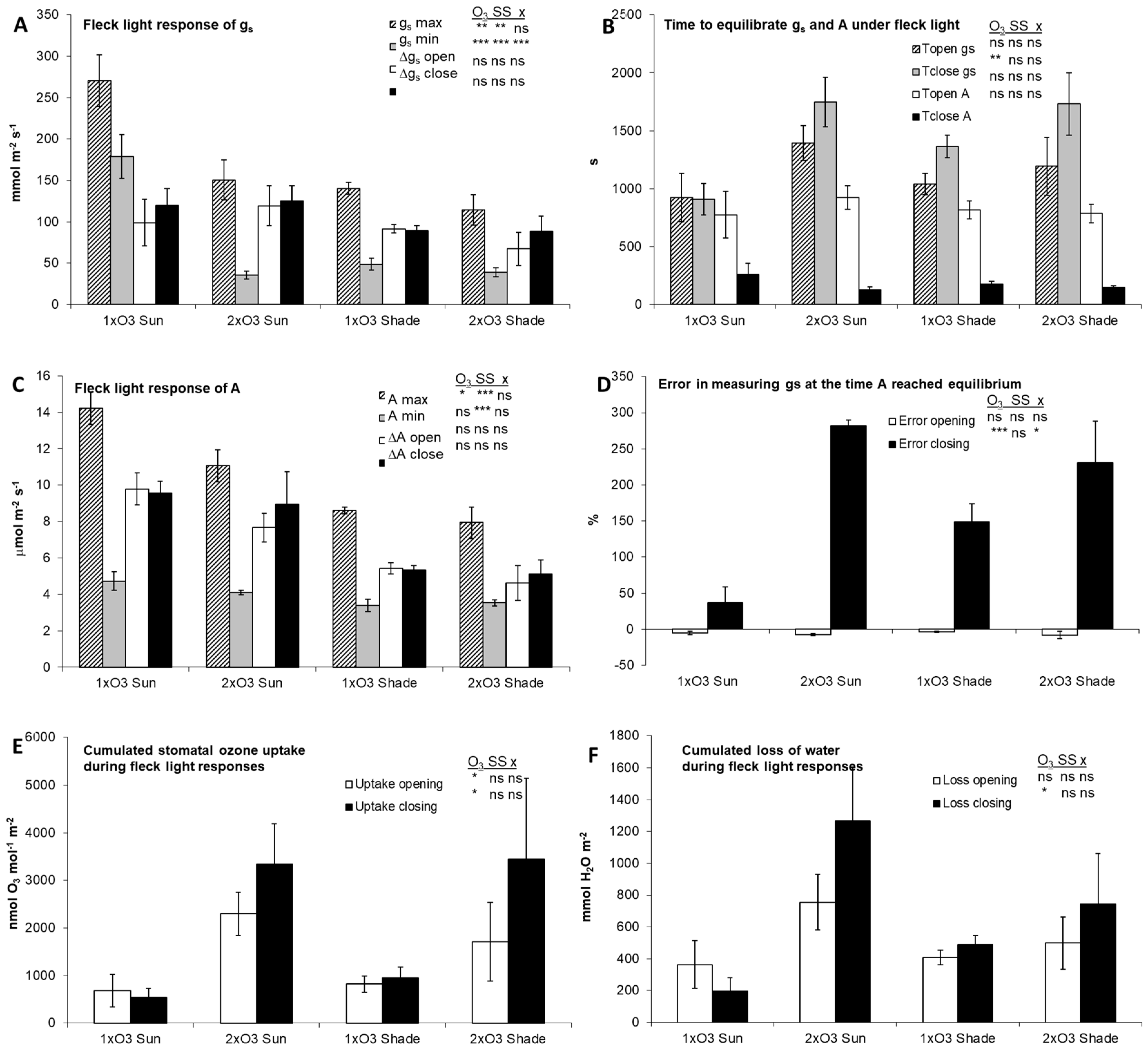
3.2. Leaf-Level Flecky Light Responses
4. Discussion
4.1. Responses of Tree-Level Transpiration
4.2. Steady-State and Dynamic Responses of Leaf-Level Gas Exchange (Sun/Shade)
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef]
- Paoletti, E. Ozone impacts on forests. Cab Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2007, 2, 13. [Google Scholar] [CrossRef]
- Mills, G.; Pleijel, H.; Malley, C.S.; Sinha, B.; Cooper, O.R.; Schultz, M.G.; Neufeld, H.S.; Simpson, D.; Sharps, K.; Feng, Z.; et al. Tropospheric Ozone Assessment Report: Present-day tropospheric ozone distribution and trends relevant to vegetation. Elem. Sci. Anthr. 2018, 6, 47. [Google Scholar] [CrossRef]
- Matyssek, R.; Kozovits, A.R.; Wieser, G.; King, J.; Rennenberg, H. Woody-plant ecosystems under climate change and air pollution—response consistencies across zonobiomes? Tree Physiol. 2017, 37, 706–732. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Feng, Z.; Catalayud, V.; Yuan, X.; Xu, Y.; Paoletti, E. A meta-analysis on growth, physiological, and biochemical responses of woody species to ground-level ozone highlights the role of plant functional types. Plant Cell Environ. 2017, 40, 2369–2380. [Google Scholar] [CrossRef] [PubMed]
- Grulke, N.E.; Heath, R.L. Ozone effects on plants in natural ecosystems. Plant Biol. 2019, 22, 1435–8603. [Google Scholar] [CrossRef] [PubMed]
- Matyssek, R.; Kozovits, A.R.; Schnitzler, J.; Pretzsch, J.; Dieler, J.; Wieser, G. Forest trees under air pollution as a factor of climate change. In Trees in a Changing Environment; Tausz, M., Grulke, N., Eds.; Plant Ecophysiology; Springer: Berlin/Heidelberg, Germany, 2014; Volume 9, pp. 117–163. [Google Scholar]
- Weigt, R.; Haeberle, K.-H.; Roetzer, T.; Matyssek, R. Whole-tree seasonal nitrogen uptake and partitioning in adult Fagus sylvatica L. and Picea abies L. [Karst.] trees exposed to elevated ground-level ozone. Environ. Pollut. 2015, 196, 511–517. [Google Scholar] [CrossRef]
- McLaughlin, S.B.; Nosal, M.; Wullschleger, S.D.; Sun, G. Interactive effects of ozone and climate on tree growth and water use in a southern Appalachian forest in the USA. New Phytol. 2007, 174, 109–124. [Google Scholar] [CrossRef]
- Matyssek, R.; Wieser, G.; Fleischmann, F.; Grünhage, L. Ozone Research, quo vadis? Lessons from the free-air canopy fumigation experiment at Kranzberg Forest. In Climate Change, Air Pollution and Global Challenges: Understanding and Perspectives from Forest Research; Matyssek, R., Clarke, N., Cudlin, P., Mikkelsen, T.N., Tuovinen, J.-P., Wieser, G., Paoletti, E., Eds.; Developments in Environmental Science 13; Elsevier: Amsterdam, The Netherlands, 2013; pp. 103–129. [Google Scholar]
- Kolb, T.E.; Matyssek, R. Limitations and perspectives about scaling ozone impact in trees. Environ. Pollut. 2001, 115, 373–393. [Google Scholar] [CrossRef]
- Löw, M.; Herbinger, K.; Nunn, A.J.; Haberle, K.-H.; Leuchner, M.; Heerdt, C.; Werner, H.; Wipfler, P.; Pretzsch, H.; Tausz, M.; et al. Extraordinary drought of 2003 overrules ozone impact on adult beech trees (Fagus sylvatica). Trees 2006, 20, 539–548. [Google Scholar] [CrossRef]
- Paoletti, E.; Grulke, N.E. Does living in elevated CO2 ameliorate tree response to ozone? A review on stomatal responses. Environ. Pollut. 2005, 137, 483–493. [Google Scholar] [CrossRef] [PubMed]
- McAinsh, M.R.; Evans, N.H.; Montgomery, L.T.; North, K.A. Calcium signalling in stomatal responses to pollutants. New Phytol. 2002, 153, 441–447. [Google Scholar] [CrossRef]
- Paoletti, E.; Grulke, N.E. Ozone exposure and stomatal sluggishness in different plant physiognomic classes. Environ. Pollut. 2010, 158, 2664–2671. [Google Scholar] [CrossRef] [PubMed]
- Matyssek, R.; Günthardt-Goerg, M.S.; Keller, T.; Scheidegger, C. Impairment of the gas exchange and structure in birch leaves (Betula pendula) caused by low ozone concentrations. Trees 1991, 5, 5–13. [Google Scholar] [CrossRef]
- Grulke, N.E. Physiological responses of ponderosa pine to gradients of environmental stressors. In Oxidant Air Pollution Impacts in the Montane Forests of Southern California; Miller, P.R., McBride, J.R., Eds.; Springer-Verlag: New York, NY, USA, 1999. [Google Scholar]
- Paoletti, E.; Nali, C.; Lorenzini, G. Early responses to acute ozone in two Fagus sylvatica clones differing in xeromorphic adaptations: Photosynthetic and stomatal processes, membrane and epicuticular characteristics. Environ. Monit. Assess. 2007, 128, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Grulke, N.E.; Paoletti, E.; Heath, R.L. Chronic vs. short-term acute O3 exposure effects on nocturnal transpiration in two Californian oaks. Sci. World J. 2007, 7, 134–140. [Google Scholar] [CrossRef][Green Version]
- Grulke, N.E.; Paoletti, E.; Heath, R.L. Comparison of calculated and measured foliar O3 flux in crop and forest species. Environ. Pollut. 2007, 146, 640–647. [Google Scholar] [CrossRef]
- Hoshika, Y.; Carriero, G.; Feng, Z.; Zhang, Y.; Paoletti, E. Determinants of stomatal sluggishness in ozone-exposed deciduous tree species. Sci. Total Environ. 2014, 481, 453–458. [Google Scholar] [CrossRef]
- Hoshika, Y.; De Carlo, A.; Baraldi, R.; Neri, L.; Carrari, E.; Agathokleous, E.; Zhang, L.; Fares, S.; Paoletti, E. Ozone-induced impairment of night-time stomatal closure in O3-sensitive poplar clone is affected by nitrogen but not by phosphorus enrichment. Sci. Total Environ. 2019, 692, 713–722. [Google Scholar] [CrossRef]
- Matyssek, R.; Gunthardt-Goerg, M.; Maurer, S.; Keller, T. Nighttime exposure to ozone reduces whole-plant production in Betula pendula. Tree Physiol. 1995, 15, 159–165. [Google Scholar] [CrossRef]
- Matyssek, R.; Baumgarten, M.; Hummel, U.; Häberle, K.-H.; Kitao, M.; Wieser, G. Canopy-level stomatal narrowing in adult Fagus sylvatica under O3 stress–means of preventing enhanced O3 uptake under high O3 exposure? Environ. Pollut. 2015, 196, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Paoletti, E. Ozone slows stomatal response to light and leaf wounding in a Mediterranean evergreen broadleaf, Arbutus unedo. Environ. Pollut. 2005, 134, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Patterson, M.T.; Rundel, P.W. Seasonal physiological responses of ozone stressed Jeffrey pine in Sequoia National Park, California. In Effects of Air Pollution on Western Forests; Olson, R.K., Lefohn, A., Eds.; Air and Waste Management Association: Pittsburgh, PA, USA, 1989. [Google Scholar]
- Pearson, M.; Mansfield, T.A. Interacting effects of ozone and water stress on the stomatal resistance of beech (Fagus sylvatica L.). New Phytol. 1993, 123, 351–358. [Google Scholar] [CrossRef]
- Keller, T.; Häsler, R. The influence of a fall fumigation with ozone on the stomatal behavior of spruce and fir. Oecologia 1984, 64, 284–286. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Lassoie, J.P. Effects of low level ozone exposure on leaf diffusive conductance and water use efficiency caused be air pollutants. Plant Cell Environ. 1984, 7, 661–668. [Google Scholar] [CrossRef]
- Morison, J.I.L. Stomatal response to increased CO2 concentration. J. Exp. Bot. 1998, 49, 443–452. [Google Scholar] [CrossRef]
- Wilkinson, S.; Davies, W. Drought, ozone, ABA and ethylene: New insights from cell to plant community. Plant Cell Environ. 2010, 33, 510–525. [Google Scholar] [CrossRef]
- Hoshika, Y.; Katata, G.; Deushi, M.; Watanabe, M.; Koike, T.; Paoletti, E. Ozone-induced stomatal sluggishness changes carbon and water balance of temperate deciduous forests. Sci. Rep. 2016, 5, 9871. [Google Scholar] [CrossRef]
- Grulke, N.E.; Alonso, R.; Nguyen, T.; Cascio, C.; Dobrowolski, W. Stomata open at night in pole-sized and mature ponderosa pine: Implications for O3 exposure metrics. Tree Physiol. 2004, 24, 1001–1010. [Google Scholar] [CrossRef]
- McLaughlin, S.B.; Wullschleger, S.D.; Sun, G.; Nosal, M. Interactive effects of ozone and climate on water use, soil moisture content and streamflow in a southern Appalachian forest in the USA. New Phytol. 2007, 174, 125–136. [Google Scholar] [CrossRef]
- Giovannelli, A.; Traversi, M.L.; Anichini, M.; Hoshika, Y.; Fares, S.; Paoletti, E. Effect of long-term vs. short-term ambient ozone exposure on radial stem growth, sap flux and xylem morphology of O3-sensitive poplar trees. Forests 2019, 10, 396. [Google Scholar] [CrossRef]
- Sun, G.; Mclaughlin, S.M.; Porter, J.H.M.; Uddling, J.; Mulholland, P.J.; Adams, M.B.; Pederson, N. Interactive influences of ozone and climate on streamflow of forested watersheds. Glob. Chang. Biol. 2012, 18, 3395–3409. [Google Scholar] [CrossRef]
- Hoshika, Y.; Watanabe, M.; Carrari, E.; Paoletti, E.; Koike, T. Ozone-induced stomatal sluggishness changes stomatal parameters of Jarvis-type model in white birch and deciduous oak. Plant Biol. 2018, 20, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Matyssek, R.; Bahnweg, G.; Ceulemans, R.; Fabian, P.; Grill, D.; Hanke, D.E.; Kraigher, H.; Osswald, W.; Rennenberg, H.; Sandermann, H.; et al. Synopsis of the CASIROZ case study: Carbon sink strength of Fagus sylvatica L. in a changing environment: Experimental risk assessment of mitigation by chronic ozone impact. Plant Biol. 2007, 9, 163–180. [Google Scholar] [CrossRef] [PubMed]
- Matyssek, R.; Wieser, G.; Ceulemans, R.; Rennenberg, H.; Pretzsch, H.; Haberer, K.; Löw, M.; Nunn, J.J.; Werner, H.; Wipfler, P.; et al. Enhanced ozone strongly reduces carbon sink strength of adult beech (Fagus sylvatica)-Resume from the free-air fumigation study at Kranzberg Forest. Environ. Pollut. 2010, 158, 2527–2532. [Google Scholar] [CrossRef]
- Pearcy, F.W. Sunflecks and photosynthesis in plant canopies. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1990, 41, 421–453. [Google Scholar] [CrossRef]
- Hoshika, Y.; De Marco, A.; Materassi, A.; Paoletti, E. Light intensity affects ozone-induced stomatal sluggishness in snapbean. Water Air Soil Pollut. 2016, 227, 419–425. [Google Scholar] [CrossRef]
- Nunn, A.J.; Reiter, I.M.; Häberle, K.H.; Werner, H.; Langebartels, C.; Sandermann, H.; Heerdt, C.; Fabian, P.; Matyssek, R. “Free-Air” ozone canopy fumigation in an old-growth mixed forest: Concept and observations in beech. Phyton 2002, 42, 105–119. [Google Scholar]
- Werner, H.; Fabian, P. Free-air fumigation of mature trees. Environ. Sci. Pollut. Res. 2002, 9, 117–121. [Google Scholar] [CrossRef]
- Matyssek, R. Ozone sensitivity of adult forest trees. Plant Biol. 2007, 9, 163–356. [Google Scholar] [CrossRef]
- CLRTAP. Mapping Critical Levels for Vegetation, Chapter III of manual on methodologies and criteria for modelling and mapping critical loads and levels and air pollution effects, risks and trends. UNECE Convention on Long-range Transboundary Air Pollution. 2017. Available online: http://icpvegetation.ceh.ac.uk/ (accessed on 18 October 2017).
- Granier, A. Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements. Tree Physiol. 1987, 3, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Laisk, A.; Kull, O.; Moldau, H. Ozone concentration in leaf intercellular air spaces is close to zero. Plant Physiol. 1989, 90, 1163–1167. [Google Scholar] [CrossRef] [PubMed]
- Wieser, G.; Matyssek, R.; Then, C.; Cieslik, S.; Paoletti, E.; Ceulemans, R. Upscaling ozone flux in forests from leaf to landscape. Ital. J. Agron. 2008, 1, 35–41. [Google Scholar] [CrossRef]
- Draper, N.; Smith, H. Applied Regression Analysis; John Wiley and Sons Inc.: New York, NY, USA, 1981; p. 209. [Google Scholar]
- Jung, M.; Reichstein, M.; Ciais, P.; Seneviratne, S.I.; Sheffield, J.; Goulden, M.L.; Bonan, G.; Cescatti, A.; Chen, J.; de Jeu, R.; et al. Recent Deceleration Glob. Land Evapotranspiration Due Moisture Supply Limit. Nat. 1981, 467, 951–954. [Google Scholar]
- Pääkkonen, E.; Vahala, J.; Pohjola, M.; Holopainen, T.; Karenlampi, L. Physiological, stomatal and ultrastructural ozone responses in birch (Betula pendula Roth.) are modified by water stress. Plant Cell Environ. 1998, 21, 671–684. [Google Scholar] [CrossRef]
- Li, L.; Manning, W.J.; Tong, L.; Wang, X.K. Chronic drought stress reduced but not protected Shantung maple (Acer truncatum Bunge.) from adverse effects of ozone O3 on growth and physiology in the suburb of Beijing. China. Environ. Pollut. 2015, 201, 34–41. [Google Scholar] [CrossRef]
- Gao, F.; Catalayud, V.; Paoletti, E.; Hoshika, Y.; Feng, Z. Water stress mitigates the negative effects of ozone on photosynthesis and biomass in poplar plants. Environ. Pollut. 2007, 230, 268–279. [Google Scholar] [CrossRef]
- Grulke, N.E.; Preisler, H.K.; Rose, C.; Kirsch, J.; Balduman, L. O3 uptake and drought stress effects on carbon acquisition of ponderosa pine in natural stands. New Phytol. 2002, 154, 621–631. [Google Scholar] [CrossRef]
- Paoletti, E.; De Marco, A.; Anav, A.; Gasparini, P.; Pompei, E. Five-year volume growth of European beech does not respond to ozone pollution in Italy. Environ. Sci. Pollut. Res. 2018, 25, 8233–8239. [Google Scholar] [CrossRef]
- Hoshika, Y.; Moura, B.B.; Paoletti, E. Ozone risk assessment in three oak species as affected by soil water availability. Environ. Sci. Pollut. Res. 2018, 25, 8125–8136. [Google Scholar] [CrossRef]
- Calatayud, V.; Cerveró, J.; Sanz, M.J. Foliar, physiological and growth responses of four maple species exposed to ozone. Water Air Soil Pollut. 2007, 185, 239–254. [Google Scholar] [CrossRef]
- Nunn, A.J.; Reiter, I.M.; Haberle, K.-H.; Langebartels, C.; Bahnweg, G.; Pretzsch, H.; Sandermann, H.; Matyssek, R. Response patterns in adult forest trees to chronic ozone stress: Identification of variations and consistencies. Environ. Pollut. 2005, 136, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Kitao, M.; Löw, M.; Heerdt, C.; Grams, T.E.E.; Häberle, K.H.; Matyssek, R. Effects of chronic elevated ozone exposure on gas exchange responses of adult beech trees (Fagus sylvatica) as related to the within-canopy light gradient. Environ. Pollut. 2009, 157, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Lombardozzi, D.; Sparks, J.P.; Bonan, G.; Levis, S. Ozone exposure causes a decoupling of conductance and photosynthesis: Implications for the Ball-Berry stomatal conductance model. Oecologia 2012, 169, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Long, S.P.; Bernacchi, C.J. Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Proced. Sources Error. J. Exp. Bot. 2003, 54, 2393–2401. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Cescatti, A.; Rodeghiero, M.; Tosens, T. Leaf internal diffusion conductance limits photosynthesis more strongly in older leaves of Mediterranean evergreen broad-leaved species. Plantcell Environ. 2005, 28, 1552–1566. [Google Scholar] [CrossRef]
- Matyssek, R.; Sandermann, H.; Wieser, G.; Booker, F.; Cieslik, S.; Musselman, R.; Ernst, D. The challenge of making ozone risk assessment for forest trees more mechanistic. Environ. Pollut. 2008, 156, 567–582. [Google Scholar] [CrossRef]
- Zhang, L.; Hoshika, Y.; Carrari, E.; Badea, O.; Paoletti, E. Ozone risk assessment is affected by nutrient availability: Evidence from a simulation experiment under free air controlled exposure (FACE). Environ. Pollut. 2018, 328, 812–822. [Google Scholar]
- Cermák, J.; Kucera, J.; Bauerle, W.L.; Phillips, N.; Hinckley, T.M. Tree water storage and its diurnal dynamics related to sap flow and changes in stem volume in old-growth Douglas-fir trees. Tree Physiol. 2007, 27, 181–198. [Google Scholar] [CrossRef]
- Schulze, E.-D.; Cermak, J.; Matyssek, R.; Penka, M.; Zimmermann, R.; Vasicek, F.; Gries, W.; Kucera, J. Canopy transpiration and water fluxes in the xylem of the trunk of Larix and Picea trees-a comparison of xylem flow, porometer and cuvette measurements. Oecologia 1985, 66, 475–483. [Google Scholar] [CrossRef]
- Hoshika, Y.; Omasa, K.; Paoletti, E. Whole-tree water use efficiency is decreased by ambient ozone and not affected by O3-induced stomatal sluggishness. PLoS ONE 2012, 7, e39270. [Google Scholar] [CrossRef] [PubMed]
- Sicard, P.; Anav, A.; De Marco, A.; Paoletti, E. Projected global tropospheric ozone impacts on vegetation under different emission and climate scenarios. Atm. Chem. Phys. 2017, 17, 12177–12196. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | 1xO3 | 2xO3 | |
|---|---|---|---|
| Air temperature (°C) | 18.33 ± 0.79 | 17.86 ± 0.80 | ns |
| Relative humidity (%) | 71.61 ± 2.34 | 71.03 ± 2.44 | ns |
| Soil water content (%) | 27.0 ± 1.0 | 27.0 ± 0.9 | ns |
| Vapour pressure deficit (MPa) | 7.87 ± 0.96 | 6.66 ± 0.91 | ns |
| Photosynthetic photon flux density (µmol m−2 s−1) | 876 ± 52 | 876 ± 52 | ns |
| Regression | Variables | Multiple R2 | Variance Explained at Each Step, % | F | p-Level |
|---|---|---|---|---|---|
| 1xO3 beech | |||||
| ppfd | 0.32 | 32 | 14.26 | 0.0007 | |
| 1xozmax | 0.40 | 8 | 3.97 | 0.0556 | |
| vpd_avg | 0.43 | 3 | 1.69 | 0.2034 | |
| t_avg | 0.48 | 5 | 2.89 | 0.1000 | |
| t_max | 0.53 | 5 | 2.86 | 0.1024 | |
| 1xO3 spruce | |||||
| vpd_max | 0.84 | 84 | 167.09 | <0.0001 | |
| ppfd | 0.88 | 4 | 10.67 | 0.0027 | |
| t_max | 0.90 | 2 | 5.08 | 0.0319 | |
| vpd_avg | 0.91 | 1 | 3.45 | 0.0738 | |
| t_avg | 0.92 | 1 | 4.26 | 0.0487 | |
| 2xO3 beech | |||||
| ppfd | 0.54 | 54 | 36.03 | <0.0001 | |
| 2xozmax | 0.62 | 5 | 3.84 | 0.0596 | |
| 2xozavg | 0.57 | 3 | 1.90 | 0.1778 | |
| 2xO3 spruce | |||||
| ppfd | 0.79 | 79 | 120.17 | <0.0001 | |
| t_max | 0.85 | 6 | 11.74 | 0.0018 | |
| vpd_avg | 0.86 | 1 | 2.07 | 0.1611 | |
| t_avg | 0.88 | 2 | 3.04 | 0.0924 | |
| vpd_max | 0.91 | 3 | 8.30 | 0.0077 |
| 1xBTr | 1xSTr | 1xOzavg | 1xOzmax | 2xBTr | 2xSTr | 2xozavg | 2xOzmax | Vpd_Avg | Vpd_Max | t_Avg | t_Max | Ppfd | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1xBTr | 1.0000 | ||||||||||||
| p= --- | |||||||||||||
| 1xSTr | r = 0.5951 | 1.0000 | |||||||||||
| p < 0.001 | p= --- | ||||||||||||
| 1xozavg | ns | r = 0.7631 | r = 1.0000 | ||||||||||
| p < 0.001 | p= --- | ||||||||||||
| 1xozmax | ns | r = 0.8052 | r = 0.9153 | r = 1.0000 | |||||||||
| p < 0.001 | p < 0.001 | p= --- | |||||||||||
| 2xBTr | r = 0.8815 | r = 0.7909 | r = 0.5012 | r = 0.4664 | 1.0000 | ||||||||
| p < 0.001 | p < 0.001 | p = 0.003 | p = 0.006 | p= --- | |||||||||
| 2xSTr | r = 0.6790 | r = 0.9743 | r = 0.6916 | r = 0.7399 | r = 0.8132 | 1.0000 | |||||||
| p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p= --- | ||||||||
| 2xozavg | ns | r = 0.4383 | r = 0.5850 | r = 0.5000 | r = 0.4719 | r = 0.3502 | 1.0000 | ||||||
| p = 0.011 | p < 0.001 | p = 0.003 | p = 0.006 | p = 0.046 | p= --- | ||||||||
| 2xozmax | ns | r = 0.5874 | r = 0.6790 | r = 0.7152 | r = 0.4270 | r = 0.4919 | r = 0.8724 | 1.0000 | |||||
| p < 0.001 | p < 0.001 | p < 0.001 | p = 0.013 | p = 0.004 | p < 0.001 | p= --- | |||||||
| vpd_avg | r = 0.4640 | r = 0.9038 | r = 0.8138 | r = 0.8202 | r = 0.6423 | r = 0.8628 | ns | r = 0.4613 | 1.0000 | ||||
| p = 0.007 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p = 0.007 | p= --- | ||||||
| vpd_max | r = 0.4433 | r = 0.9184 | r = 0.8199 | r = 0.8689 | r = 0.6450 | r = 0.8648 | r = 0.4627 | r = 0.6013 | r = 0.9503 | 1.0000 | |||
| p = 0.010 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p = 0.007 | p < 0.001 | p < 0.001 | p= --- | ||||
| t_avg | ns | r = 0.8545 | r = 0.8297 | r = 0.8430 | r = 0.5005 | r = 0.7798 | r = 0.4029 | r = 0.5424 | r = 0.8803 | r = 0.8986 | 1.0000 | ||
| p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p = 0.020 | p = 0.001 | p < 0.001 | p < 0.001 | p= --- | ||||
| t_max | r = 0.3311 | r = 0.9079 | r = 0.8095 | r = 0.8632 | r = 0.5953 | r = 0.8506 | r = 0.4546 | r = 0.6068 | r = 0.8744 | r = 0.9420 | r = 0.9671 | 1.0000 | |
| p = 0.060 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p = 0.008 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p= --- | ||
| ppfd | r = 0.5613 | r = 0.8817 | r = 0.7394 | r = 0.7652 | r = 0.7332 | r = 0.8916 | r = 0.4401 | r = 0.5622 | r = 0.8190 | r = 0.8405 | r = 0.7096 | r = 0.7883 | 1.0000 |
| p = 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p = 0.010 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p= --- |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paoletti, E.; Grulke, N.E.; Matyssek, R. Ozone Amplifies Water Loss from Mature Trees in the Short Term But Decreases It in the Long Term. Forests 2020, 11, 46. https://doi.org/10.3390/f11010046
Paoletti E, Grulke NE, Matyssek R. Ozone Amplifies Water Loss from Mature Trees in the Short Term But Decreases It in the Long Term. Forests. 2020; 11(1):46. https://doi.org/10.3390/f11010046
Chicago/Turabian StylePaoletti, Elena, Nancy E. Grulke, and Rainer Matyssek. 2020. "Ozone Amplifies Water Loss from Mature Trees in the Short Term But Decreases It in the Long Term" Forests 11, no. 1: 46. https://doi.org/10.3390/f11010046
APA StylePaoletti, E., Grulke, N. E., & Matyssek, R. (2020). Ozone Amplifies Water Loss from Mature Trees in the Short Term But Decreases It in the Long Term. Forests, 11(1), 46. https://doi.org/10.3390/f11010046