Functional Trait Plasticity but Not Coordination Differs in Absorptive and Transport Fine Roots in Response to Soil Depth



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Root Sample Collection
2.3. Root Anatomy, Morphology, and Biomass
2.4. Root Respiration and Chemistry
2.5. Data Analysis
3. Results
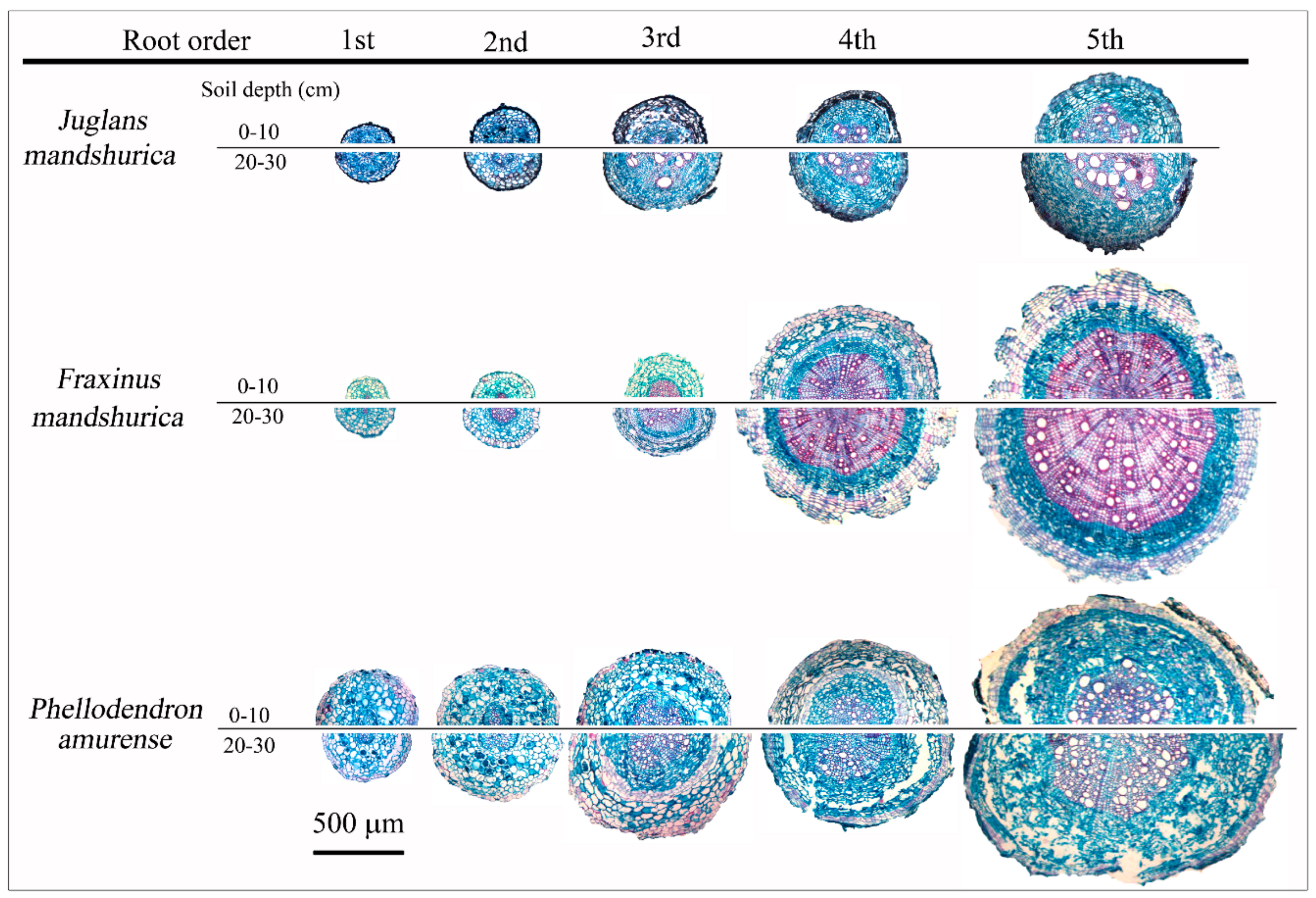
3.1. Assessment of Absorptive and Transport Fine Roots at Different Soil Layers
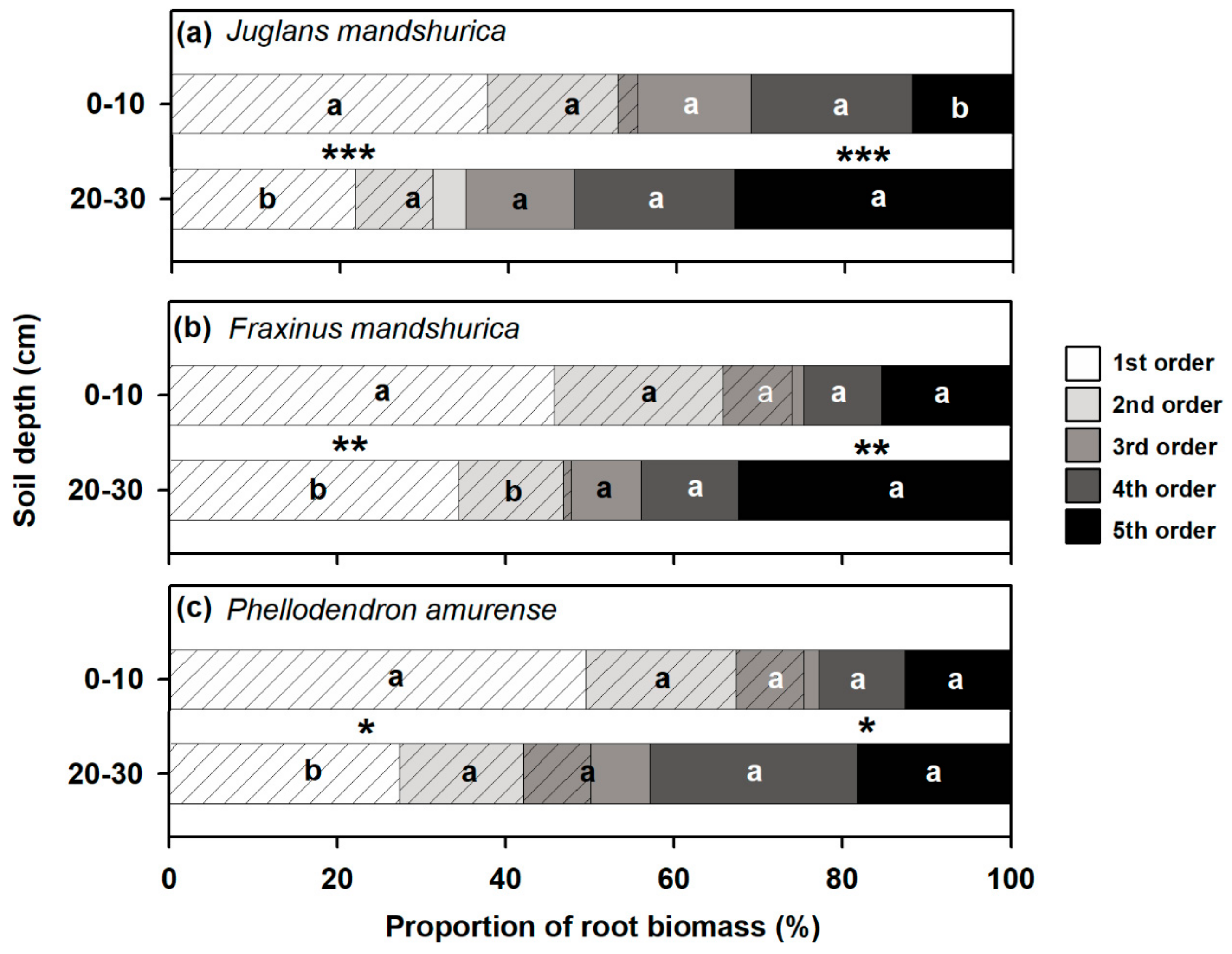
3.2. Relative Share of Biomass between Absorptive and Transport Fine Roots at Different Soil Layers
3.3. Root Functional Trait Responses and Plasticity to Soil Depth
3.4. Root Functional Trait Shift Coordinately along Soil Depth
4. Discussion
4.1. Variations in Biomass Repartition between Absorptive and Transport Fine Roots along Soil Depth
4.2. Variations of Functional Traits of Absorptive and Transport Fine Roots along Soil Depth
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Guo, D.; Xia, M.; Wei, X.; Chang, W.; Liu, Y.; Wang, Z. Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species. New Phytol. 2008, 180, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Pregitzer, K.S.; Deforest, J.L.; Burton, A.J.; Allen, M.F.; Ruess, R.W.; Hendrick, R.L. Fine root architecture of nine North American trees. Ecol. Monogr. 2002, 72, 293–309. [Google Scholar] [CrossRef]
- McCormack, M.L.; Dickie, I.A.; Eissenstat, D.M.; Fahey, T.J.; Fernandez, C.W.; Guo, D.; Helmisaari, H.; Hobbie, E.A.; Iversen, C.M.; Jackson, R.B.; et al. Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytol. 2015, 207, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Hodge, A. The plastic plant: Root responses to heterogeneous supplies of nutrients. New Phytol. 2004, 162, 9–24. [Google Scholar] [CrossRef]
- Maeght, J.; Rewald, B.; Pierret, A. How to study deep roots-and why it matters. Front Plant Sci. 2013, 4, 299. [Google Scholar] [CrossRef]
- Majdi, H.; Truus, L.; Johansson, U.; Nylund, J.; Wallander, H. Effects of slash retention and wood ash addition on fine root biomass and production and fungal mycelium in a Norway spruce stand in SW Sweden. Forest Ecol. Manag. 2008, 255, 2109–2117. [Google Scholar] [CrossRef]
- McElrone, A.J.; Pockman, W.T.; Vilalta, J.M.; Jackson, R.B. Variation in xylem structure and function in stems and roots of trees to 20 m depth. New Phytol. 2004, 163, 507–517. [Google Scholar] [CrossRef]
- Prieto, I.; Roumet, C.; Cardinael, R.; Dupraz, C.; Jourdan, C.; Kim, J.H.; Maeght, J.L.; Mao, Z.; Pierret, A.; Portillo, N.; et al. Root functional parameters along a land-use gradient: Evidence of a community-level economics spectrum. J. Ecol. 2015, 103, 361–373. [Google Scholar] [CrossRef]
- Wang, Y.; Dong, X.; Wang, H.; Wang, Z.; Gu, J. Root tip morphology, anatomy, chemistry and potential hydraulic conductivity vary with soil depth in three temperate hardwood species. Tree Physiol. 2016, 36, 99–108. [Google Scholar] [CrossRef]
- Wang, G.; Liu, F.; Xue, S. Nitrogen addition enhanced water uptake by affecting fine root morphology and coarse root anatomy of Chinese pine seedlings. Plant Soil 2017, 418, 177–189. [Google Scholar] [CrossRef]
- Trocha, L.K.; Bułaj, B.; Kutczyńska, P.; Mucha, J.; Rutkowski, P.; Zadworny, M. The interactive impact of root branch order and soil genetic horizon on root respiration and nitrogen concentration. Tree Physiol. 2017, 37, 1055–1068. [Google Scholar] [CrossRef] [PubMed]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, G.; Wang, N.; Wang, Z.; Gu, J. Effects of morphology and stand structure on root biomass and length differed between absorptive and transport roots in temperate trees. Plant Soil 2019, 442, 355–367. [Google Scholar] [CrossRef]
- Wang, Z.; Guo, D.; Wang, X.; Gu, J.; Mei, L. Fine root architecture, morphology, and biomass of different branch orders of two Chinese temperate tree species. Plant Soil 2006, 288, 155–171. [Google Scholar] [CrossRef]
- Zadworny, M.; McCormack, M.L.; Mucha, J.; Reich, P.B.; Oleksyn, J. Scots pine fine roots adjust along a 2000-km latitudinal climatic gradient. New Phytol. 2016, 212, 389–399. [Google Scholar] [CrossRef]
- Kramer-Walter, K.R.; Laughlin, D.C. Root nutrient concentration and biomass allocation are more plastic than morphological traits in response to nutrient limitation. Plant Soil 2017, 416, 539–550. [Google Scholar] [CrossRef]
- Nicotra, A.B.; Atkin, O.K.; Bonser, S.P.; Davidson, A.M.; Finnegan, E.J.; Mathesius, U.; Poot, P.; Purugganan, M.D.; Richards, C.L.; Valladares, F.; et al. Plant phenotypic plasticity in a changing climate. Trends Plant Sci. 2010, 15, 684–692. [Google Scholar] [CrossRef]
- Ryser, P.; Eek, L. Consequences of phenotypic plasticity vs. interspecific differences in leaf and root traits for acquisition of aboveground and belowground resources. Am. J. Bot. 2000, 3, 402–411. [Google Scholar] [CrossRef]
- Valladares, F.; Sanchez-Gomez, D.; Zavala, M.A. Quantitative estimation of phenotypic plasticity: Bridging the gap between the evolutionary concept and its ecological applications. J. Ecol. 2006, 94, 1103–1116. [Google Scholar] [CrossRef]
- Freschet, G.T.; Swart, E.M.; Cornelissen, J.H.C. Integrated plant phenotypic responses to contrasting above- and below-ground resources: Key roles of specific leaf area and root mass fraction. New Phytol. 2015, 206, 1247–1260. [Google Scholar] [CrossRef]
- Fort, F.; Cruz, P.; Catrice, O.; Delbrut, A.; Luzarreta, M.; Stroia, C.; Jouany, C. Root functional trait syndromes and plasticity drive the ability of grassland Fabaceae to tolerate water and phosphorus shortage. Environ. Exp. Bot. 2015, 110, 62–72. [Google Scholar] [CrossRef]
- Makita, N.; Hirano, Y.; Mizoguchi, T.; Kominami, Y.; Dannoura, M.; Ishii, H.; Finér, L.; Kanazawa, Y. Very fine roots respond to soil depth: Biomass allocation, morphology, and physiology in a broad-leaved temperate forest. Ecol. Res. 2011, 26, 95–104. [Google Scholar] [CrossRef]
- Miyatani, K.; Mizusawa, Y.; Okada, K.; Tanikawa, T.; Makita, N.; Hirano, Y. Fine root traits in Chamaecyparis obtusa forest soils with different acid buffering capacities. Trees 2016, 30, 415–429. [Google Scholar] [CrossRef]
- Jia, S.; Mclaughlin, N.B.; Gu, J.; Li, X.; Wang, Z. Relationships between root respiration rate and root morphology, chemistry and anatomy in Larix gmelinii and Fraxinus mandshurica. Tree Physiol. 2013, 33, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Xu, Y.; Dong, X.; Wang, H.; Wang, Z. Root diameter variations explained by anatomy and phylogeny of 50 tropical and temperate tree species. Tree Physiol. 2014, 34, 415–425. [Google Scholar] [CrossRef]
- Anfodillo, T.; Petit, G.; Crivellaro, A. Axial conduit widening in woody species: A still neglected anatomical pattern. IAWA J. 2013, 34, 352–364. [Google Scholar] [CrossRef]
- Olson, M.E.; Rosell, J.A. Vessel diameter–stem diameter scaling across woody angiosperms and the ecological causes of xylem vessel diameter variation. New Phytol. 2013, 197, 1204–1213. [Google Scholar] [CrossRef]
- Burton, A.J.; Pregitzer, K.S.; Ruess, R.W.; Hendrick, R.L.; Allen, M.F. Root respiration in North American forests: Effects of nitrogen concentration and temperature across biomes. Oecologia 2002, 131, 559–568. [Google Scholar] [CrossRef]
- Tyree, M.T.; Ewers, F.W. The hydraulic architecture of trees and other woody plants. New Phytol. 1991, 119, 345–360. [Google Scholar] [CrossRef]
- Sterck, F.J.; Zweifel, R.; Sass-Klaassen, U.; Qumruzzaman, C. Persisting soil drought reduces leaf specific conductivity in Scots pine (Pinus sylvestris) and pubescent oak (Quercus pubescens). Tree Physiol. 2008, 28, 529–536. [Google Scholar] [CrossRef]
- Zhang, S.; Cao, K.; Fan, Z.; Zhang, J. Potential hydraulic efficiency in angiosperm trees increases with growth-site temperature but has no trade-off with mechanical strength. Glob. Ecol. Biogeogr. 2013, 22, 971–981. [Google Scholar] [CrossRef]
- Scoffoni, C.; Kunkle, J.; Pasquet-Kok, J.; Vuong, C.; Patel, A.J.; Montgomery, R.A.; Givnish, T.J.; Sack, L. Light-induced plasticity in leaf hydraulics, venation, anatomy, and gas exchange in ecologically diverse Hawaiian lobeliads. New Phytol. 2015, 207, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Tsakaldimi, M.; Tsitsoni, T.; Ganatsas, P.; Zagas, T. A comparison of root architecture and shoot morphology between naturally regenerated and container-grown seedlings of Quercus ilex. Plant Soil 2009, 324, 103–113. [Google Scholar] [CrossRef]
- Wickham, H.; Chang, W.; Henry, L.; Pederson, T.L.; Takahashi, K.; Wikle, C.; Woo, K.; Yutani, H.; RStudio. ggplot2: Create Elegant Data Visualisations Using the Grammar of Graphics. Available online: http://lib.ugent.be/CRAN/web/packages/ggplot2/index.html (accessed on 7 November 2019).
- Borken, W.; Kossmann, G.; Matzner, E. Biomass, morphology and nutrient contents of fine roots in four Norway spruce stands. Plant Soil 2007, 292, 79–93. [Google Scholar] [CrossRef]
- Liu, R.; Huang, Z.; McCormack, M.L.; Zhou, X.; Wan, X.; Yu, Z.; Wang, M.; Zheng, L. Plasticity of fine-root functional traits in the litter layer in response to nitrogen addition in a subtropical forest plantation. Plant Soil 2017, 415, 317–330. [Google Scholar] [CrossRef]
- Wang, W.; Wu, X.; Hu, K.; Liu, J.; Tao, J. Understorey fine root mass and morphology in the litter and upper soil layers of three Chinese subtropical forests. Plant Soil 2016, 406, 219–230. [Google Scholar] [CrossRef]
- Arredondo, J.T.; Johnson, D.A. Root architecture and biomass allocation of three range grasses in response to nonuniform supply of nutrients and shoot defoliation. New Phytol. 1999, 143, 373–385. [Google Scholar] [CrossRef]
- Fransen, B.; De Kroon, H. Long-term disadvantages of selective root placement: Root proliferation and shoot biomass of two perennial grass species in a 2-year experiment. J. Ecol. 2001, 89, 711–722. [Google Scholar] [CrossRef]
- Liu, B.; Li, H.; Zhu, B.; Koide, R.T.; Eissenstat, D.M.; Guo, D. Complementarity in nutrient foraging strategies of absorptive fine roots and arbuscular mycorrhizal fungi across 14 coexisting subtropical tree species. New Phytol. 2015, 208, 125–136. [Google Scholar] [CrossRef]
- Gebauer, R.; Volařík, D. Root hydraulic conductivity and vessel structure modification with increasing soil depth of two oak species: Quercus pubescens and Quercus robur. Trees 2013, 27, 523–531. [Google Scholar] [CrossRef]
- Pate, J.S.; Jeschke, W.D.; Aylward, M.J. Hydraulic architecture and xylem structure of the dimorphic root systems of South-West Australian species of Proteaceae. J. Exp. Bot. 1995, 289, 907–915. [Google Scholar] [CrossRef]
- Aguirrezabal, L.A.; Pellerin, S.; Rtradieu, F. Carbon nutrition, root branching and elongation: Can the present state of knowledge allow a predictive approach at a whole-plant level? Environ. Exp. Bot. 1993, 33, 121–130. [Google Scholar] [CrossRef]
- North, G.B. A long drink of water how xylem changes with depth. New Phytol. 2004, 163, 447–449. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; Laskowski, M.J.; Burton, A.J.; Lessard, V.C.; Zak, D.R. Variation in sugar maple root respiration with root diameter and soil depth. Tree Physiol. 1998, 18, 665–670. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Layer | Root Order | Tree Species | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Juglans mandshurica | Fraxinus mandshurica | Phelldendron amurense | ||||||||
| n | Absorptive Roots | Transportive Roots | n | Absorptive Roots | Transportive Roots | n | Absorptive Roots | Transportive Roots | ||
| 0–10 cm | 1 | 30 | 100 | 0 | 30 | 100 | 0 | 30 | 100 | 0 |
| 2 | 21 | 100 | 0 | 30 | 100 | 0 | 30 | 100 | 0 | |
| 3 | 20 | 15 | 85 | 20 | 85 | 15 | 11 | 82 | 18 | |
| 4 | 14 | 0 | 100 | 20 | 0 | 100 | 5 | 0 | 100 | |
| 5 | 7 | 0 | 100 | 7 | 0 | 100 | 5 | 0 | 100 | |
| 20–30 cm | 1 | 21 | 100 | 0 | 30 | 100 | 0 | 30 | 100 | 0 |
| 2 | 30 | 70 | 30 | 30 | 100 | 0 | 27 | 100 | 0 | |
| 3 | 18 | 0 | 100 | 20 | 10 | 90 | 17 | 53 | 47 | |
| 4 | 11 | 0 | 100 | 8 | 0 | 100 | 6 | 0 | 100 | |
| 5 | 8 | 0 | 100 | 7 | 0 | 100 | 5 | 0 | 100 | |
| Source of Variation | df | p Value | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RD | SRL | RTD | RBD | RRR | RNC | RCC | CNR | CT | SD | SRR | MECD | MACD | HWCD | NC | Ks | PRB | ||
| Species | 2 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.841 |
| Soil depth | 1 | <0.001 | 0.107 | 0.039 | 0.002 | <0.001 | <0.001 | <0.001 | <0.001 | 0.422 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.005 | 0.605 |
| Root order | 4 | <0.001 | <0.001 | 0.000 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Species × Soil depth | 2 | <0.001 | 0.173 | 0.024 | 0.512 | <0.001 | 0.002 | 0.015 | 0.043 | <0.001 | 0.003 | <0.001 | <0.001 | <0.001 | <0.001 | 0.099 | <0.001 | 0.666 |
| Soil depth × Root order | 4 | <0.001 | <0.001 | 0.004 | 0.032 | <0.001 | <0.001 | 0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Species × Root order | 8 | <0.001 | 0.082 | 0.216 | 0.111 | <0.001 | <0.001 | 0.026 | <0.001 | 0.994 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.068 | 0.122 |
| Species × Soil depth × Root order | 8 | 0.025 | 0.597 | 0.663 | 0.421 | <0.001 | <0.001 | 0.096 | <0.001 | <0.001 | 0.223 | 0.059 | <0.001 | <0.001 | <0.001 | 0.256 | 0.013 | 0.684 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Li, Z.; Wang, Z.; Gu, J. Functional Trait Plasticity but Not Coordination Differs in Absorptive and Transport Fine Roots in Response to Soil Depth. Forests 2020, 11, 42. https://doi.org/10.3390/f11010042
Wang Y, Li Z, Wang Z, Gu J. Functional Trait Plasticity but Not Coordination Differs in Absorptive and Transport Fine Roots in Response to Soil Depth. Forests. 2020; 11(1):42. https://doi.org/10.3390/f11010042
Chicago/Turabian StyleWang, Yan, Zhongyue Li, Zhengquan Wang, and Jiacun Gu. 2020. "Functional Trait Plasticity but Not Coordination Differs in Absorptive and Transport Fine Roots in Response to Soil Depth" Forests 11, no. 1: 42. https://doi.org/10.3390/f11010042
APA StyleWang, Y., Li, Z., Wang, Z., & Gu, J. (2020). Functional Trait Plasticity but Not Coordination Differs in Absorptive and Transport Fine Roots in Response to Soil Depth. Forests, 11(1), 42. https://doi.org/10.3390/f11010042