Chemical Fingerprinting of Wood Sampled along a Pith-to-Bark Gradient for Individual Comparison and Provenance Identification

 ,
,  and
and
Abstract
1. Introduction
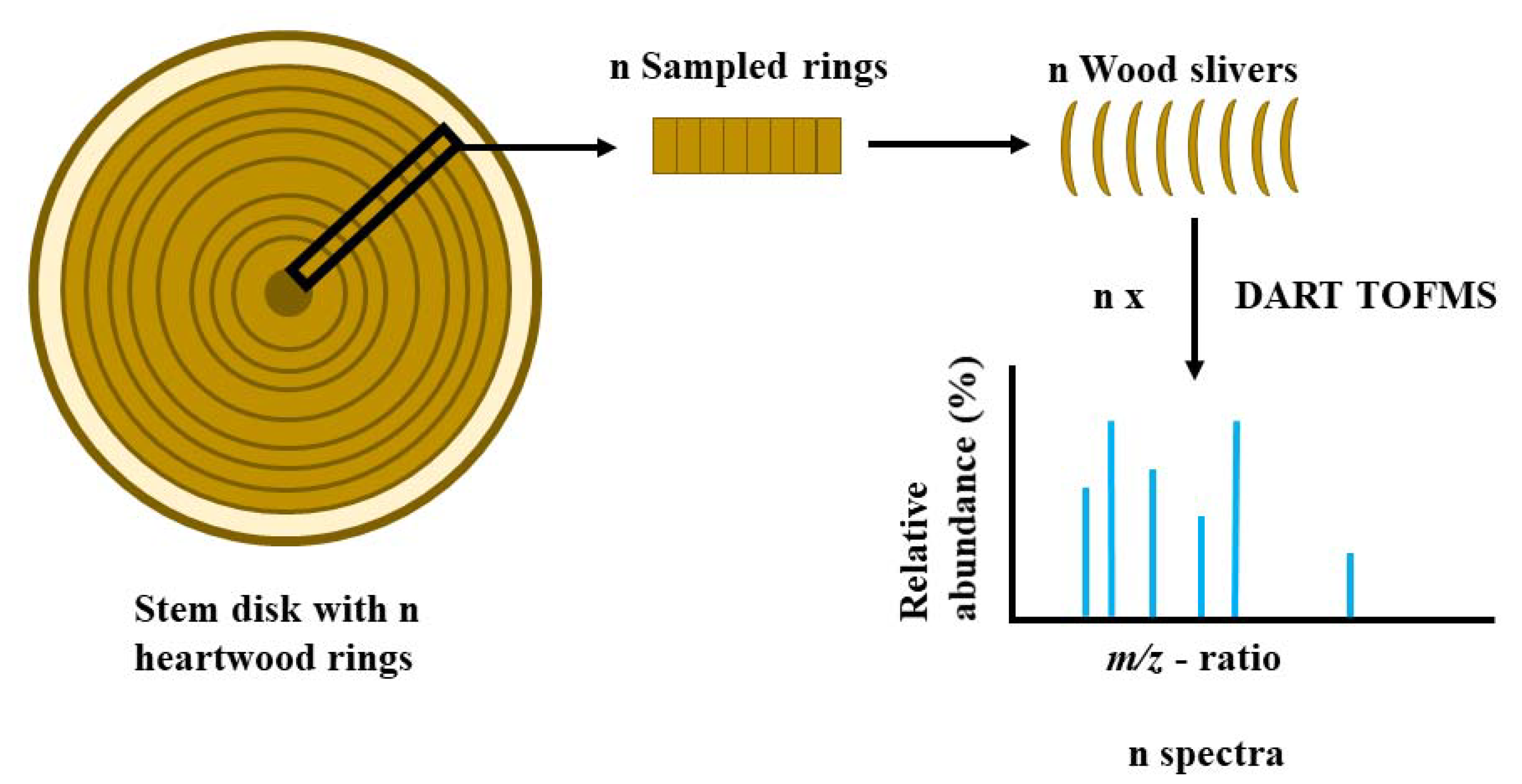
2. Materials and Methods
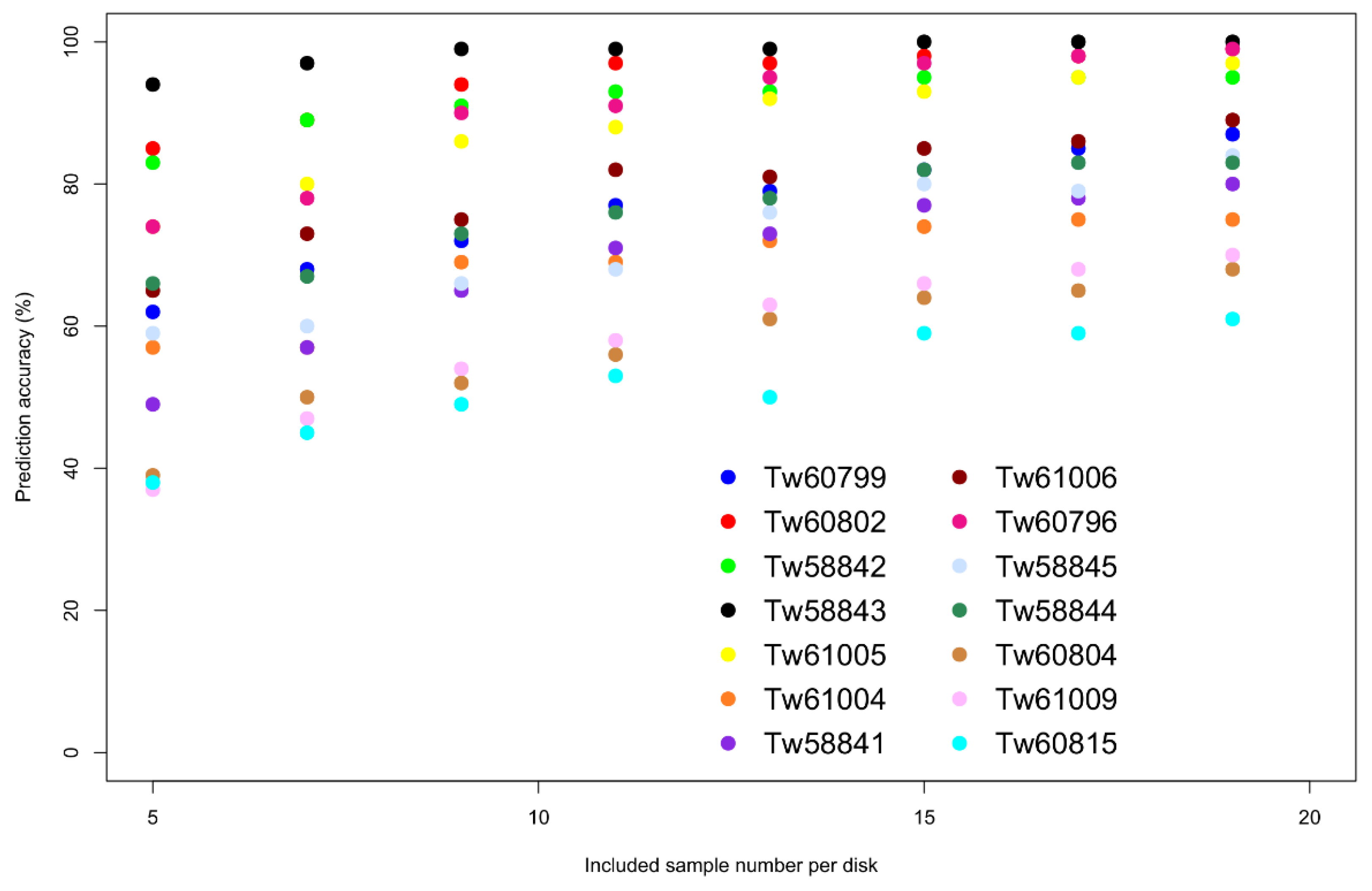
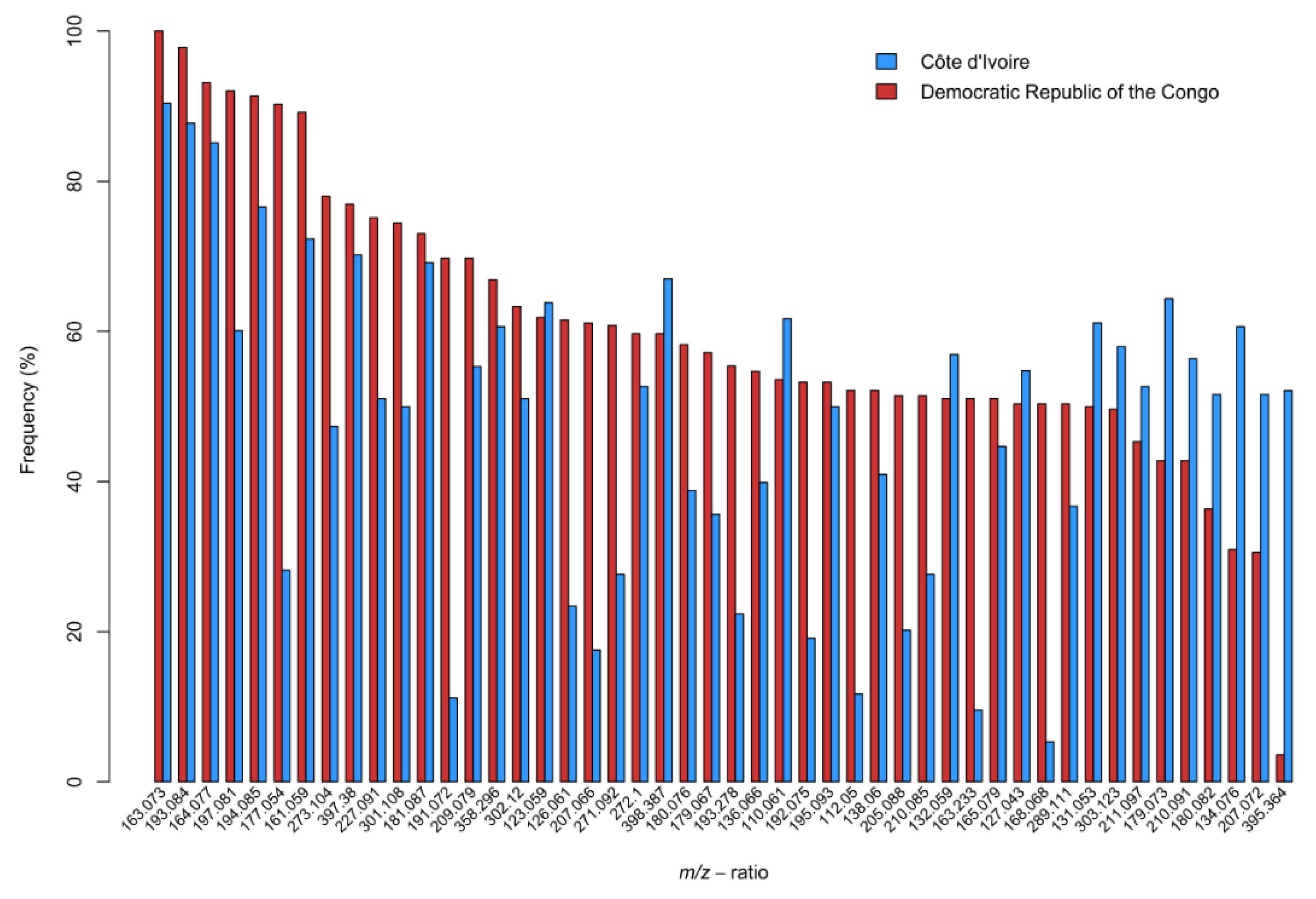
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Beeckman, H. The Luki and Yangambi Biosphere Reserves: Laboratories for climage change research and sustainable development. IOP Conf. Ser. Earth Environ. Sci. 2019, 298, 10. [Google Scholar] [CrossRef]
- May, C. Transnational Crime and the Developing World. Glob. Financ. Integr. 2017, 30, 2019. [Google Scholar]
- Hirschberger, P. Illegal wood for the European market. In An Analysis of the EU Import and Export of Illegal Wood and Related Products; Griesshammer, N., Schulmeister, A., Eds.; WWF: Frankfurt am Main, Germany, 2008; p. 45. [Google Scholar]
- Hoare, A. Tracking Illegal Logging and the Related Trade. In What Progress and Where Next; The Royal Institute of International Affairs (Chatham House): London, UK, 2015; p. 79. [Google Scholar]
- Vlam, M.; Arjen de Groot, G.; Boom, A.; Copini, P.; Laros, I.; Veldhuijzen, K.; Zakamdi, D.; Zuidema, P.A. Developing forensic tools for an African timber: Regional origin is revealed by genetic characteristics, but not by isotopic signature. Biol. Conserv. 2018, 220, 262–271. [Google Scholar] [CrossRef]
- Degen, B.; Fladung, M. Use of DNA-markers for tracing illegal logging. In Proceedings of the International Workshop Fingerprinting Methods for the Identification of Timber Origins, Bonn, Germany, 8–9 October 2007; pp. 6–14. [Google Scholar]
- Li, R.; Buongiorno, J.; Turner, J.A.; Zhu, S.; Prestemon, J. 2008 Long-term effects of eliminating illegal logging on the world forest industries, trade, and inventory. For. Policy Econ. 2008, 10, 480–490. [Google Scholar] [CrossRef]
- Nielsen, L.R.; Kjaer, E.D. Tracing Timber from Forest to Consumer with DNA Markers; Electronic publication; Danish Ministry of the Environment, Forest and Nature Agency: Copenhagen, Denmark, 2008; p. 40. Available online: www.skovognatur.dk/udgivelser (accessed on 12 December 2019).
- Lawson, S. Illegal Logging and Related Trade: Indicators of the Global Response (Chatham House—Briefing Paper); Chatham House: London, UK, 2010; p. 8. [Google Scholar]
- Dormontt, E.E.; Boner, M.; Braun, B.; Breulmann, G.; Degen, B.; Espinoza, E.; Gardner, S.; Guillery, P.; Hermanson, J.C.; Koch, G.; et al. Forensic timber identification: It’s time to integrate disciplines to combat illegal logging. Biol. Conserv. 2015, 191, 790–798. [Google Scholar] [CrossRef]
- Horacek, M.; Jakusch, M.; Krehan, H. Control of origin of larch wood: Discrimination between European (Austrian) and Siberian origin by stable isotope analysis. Rapid Commun. Mass Spectrom. 2009, 23, 3688–3692. [Google Scholar] [CrossRef]
- Kagawa, A.; Leavitt, S.W. Stable carbon isotopes of tree rings as a tool to pinpoint the geographic origin of timber. J. Wood Sci. 2010, 56, 175–183. [Google Scholar] [CrossRef]
- Förstel, H.; Boner, M.; Höltken, A.M.; Fladung, M.; Degen, B.; Zahnen, J. Fighting Illegal Logging Through the Introduction of a Combination of the Isotope Method for Identifying the Origins of Timber and DNA Analysis for Differentiation of Tree Species; Deutsche Bundesstiftung Umwelt: Osnabrück, Germany, 2011. [Google Scholar]
- Horacek, M. Christmas tree production in Europe: Control of declared geographical origin by stable isotope analysis—A pilot study. Die Bodenkult. 2012, 63, 35–41. [Google Scholar]
- Lowe, A.J.; Wong, K.N.; Tiong, Y.S.; Iyerh, S.; Chew, F.T. A DNA Method to Verify the Integrity of Timber Supply Chain; Confirming the Legal Sourcing of Merbau Timber from Logging Concession to Sawmill. Silvae Genet. 2010, 59, 263–268. [Google Scholar] [CrossRef]
- Jolivet, C.; Degen, B. Use of DNA fingerprints to control the origin of sapelli timber (Entandrophragma cylindricum) at the forest concession level in Cameroon. Forensic Sci. Int. Genet. 2012, 6, 487–493. [Google Scholar] [CrossRef]
- Degen, B.; Ward, S.E.; Lemes, M.R.; Navarro, C.; Cavers, S.; Sebbenn, A.M. Verifying the geographic origin of mahogany (Swietenia macrophylla King) with DNA-fingerprints. Forensic Sci. Int. Genet. 2013, 7, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Hung, K.; Lin, C.; Ju, L. Tracking the geographical origin of timber by DNA fingerprinting: A study of the endangered species Cinnamomum kanehirae in Taiwan. Holzforschung 2017, 71, 853–862. [Google Scholar] [CrossRef]
- Pérez-Coello, M.S.; Sanz, J.; Cabezudo, M.D. Analysis of volatile components of oak wood by solvent extraction and direct thermal desportion-gas chromatography-mass spectrometry. J. Chromotogr. 1997, 778, 427–434. [Google Scholar] [CrossRef]
- Lancaster, C.; Espinoza, E. Analysis of select Dalbergia and trade timber using direct analysis in real time and time-of-flight mass spectrometry for CITES enforcement. Rapid Commun. Mass Spectrom. 2012, 26, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- McClure, P.J.; Chavarria, G.D.; Espinoza, E. Metabolic chemotypes of CITES protected Dalbergia timbers from Africa, Madagascar, and Asia. Rapid Commun. Mass Spectrom. 2015, 29, 783–788. [Google Scholar] [CrossRef]
- Deklerck, V.; Mortier, T.; Goeders, N.; Cody, R.B.; Waegeman, W.; Espinoza, E.; Van Acker, J.; Van den Bulcke, J.; Beeckman, H. A protocol for automated timber species identification using metabolome profiling. Wood Sci. Technol. 2019, 53, 953–965. [Google Scholar] [CrossRef]
- Espinoza, E.; Lancaster, C.A.; Kreitals, N.M.; Hata, M.; Cody, R.B.; Blanchette, R.A. Distinguishing wild from cultivated agarwood (Aquilaria spp.) using direct analysis in real time and time of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 2014, 28, 281–289. [Google Scholar] [CrossRef]
- Paredes-Villanueva, K.; Espinoza, E.; Ottenburghs, J.; Sterken, M.G.; Bongers, F.; Zuidema, P.A. Chemical differentiation of Bolivian Cedrela species as a tool to trace illegal timber trade. Forestry 2018, 91, 603–613. [Google Scholar] [CrossRef]
- Finch, K.; Espinoza, E.; Jones, F.A.; Cronn, R. Source Identification of Western Oregon Douglas-Fir Wood Cores Using Mass Spectrometry and Random Forest Classification. Appl. Plant Sci. 2017, 5, 1600158. [Google Scholar] [CrossRef]
- De Ridder, M.; Trouet, V.; Van den Bulcke, J.; Hubau, W.; Van Acker, J.; Beeckman, H. A tree-ring based comparison of Terminalia superba climate-growth relationships in West and Central Africa. Trees 2013, 27, 1225–1238. [Google Scholar] [CrossRef]
- De Ridder, M.; Toirambe, B.; Van den Bulcke, J.; Bourland, N.; Van Acker, J.; Beeckman, H. Dendrochronological potential in a semi-deciduous rainforest: The case of pericopsis elata in Central Africa. Forests 2014, 5, 3087–3106. [Google Scholar] [CrossRef]
- Evans, P.D.; Mundo, I.A.; Wiemann, M.C.; Chavarria, G.D.; McClure, P.J.; Voin, D.; Espinoza, E. Identification of selected CITES-Protected Araucariaceae using DART TOFMS. IAWA J. 2017, 38, 266–281. [Google Scholar] [CrossRef]
- Deklerck, V.; Finch, K.; Gasson, P.; Van den Bulcke, J.; Van Acker, J.; Beeckman, H.; Espinoza, E. Comparison of species classification models of mass spectrometry data: Kernel Discriminant Analysis vs Random Forest; A case study of Afrormosia (Pericopsis elata (Harms) Meeuwen). Rapid Commun. Mass Spectrom. 2017, 31, 1582–1588. [Google Scholar] [CrossRef] [PubMed]
- Liaw, A.; Wiener, M. Breiman and Cutler’s Random Forests for Classification and Regression, Package “Random Forest”. Available online: http://math.furman.edu/~dcs/courses/math47/R/library/randomForest/html/00Index.html (accessed on 12 December 2019).
- Afendi, F.M.; Okada, T.; Yamazaki, M.; Morita, A.H.; Nakamura, Y.; Nakamura, K.; Ikeda, S.; Takahashi, H.; Altaf-Ul-Amin, M.; Darusman, L.; et al. KNApSAck family databases: Integrated metabolite-plant species database for multifaceted plant research. Plant Cell Physiol. 2012, 53, e1. [Google Scholar] [CrossRef]
- Höltken, A.M.; Schröder, H.; Wischnewski, N.; Degen, B.; Magel, E.; Fladung, M. Development of DNA-based methods to identify CITES-protected timber species: A case study in the Meliaceae family. Holzforschung 2012, 66, 97–104. [Google Scholar]
- Kelly, S.; Heaton, K.; Hoogewerff, J. Tracing the geographical origin of food: The application of multi-element and multi-isotope analysis. Trends Food Sci. Technol. 2005, 16, 555–567. [Google Scholar] [CrossRef]
- Colombaroli, D.; Cherubini, P.; De Ridder, M.; Saurer, M.; Toirambe, B.; Zweifel, N.; Beeckman, H. Stable carbon and oxygen isotopes in tree rings show physiological responses of Pericopsis elata to precipitation in the Congo Basin. J. Trop. Ecol. 2016, 32, 213–225. [Google Scholar] [CrossRef]
- Groulez, J.; Wood, P.J. A Monograph on Terminalia Superba; Centre Technique Forestier Tropical: Nogent-sur-Marne, France; Commonwealth Forestry Institute: Oxford, UK, 1985. [Google Scholar]
- Schmitz, N.; Blanc-Jolivet, C.; Boner, M.; Cervera, M.T.; Chavesta, M.; Cronn, R.; Degen, B.; Deklerck, V.; Diaz-Sala, C.; Dormontt, E.; et al. General Sampling Guide for Timber Tracking. In Global Timber Tracking Network, GTTN Secretariat, European Forest Institute and Thuenen Institute; The GTTN Secretariat: Braunschweig, Germany, 2019; p. 46. [Google Scholar]
- Laporte, N.T.; Stabach, J.A.; Grosch, R.; Tiffany, S.L.; Goetz, S.J. Expansion of Industrial Logging in Central Africa. Science 2007, 316, 145. [Google Scholar] [CrossRef]
- Fuller, D.O. Tropical forest monitoring and remote sensing: A new era of transparency in forest governance? Singap. J. Trop. Geogr. 2006, 27, 15–29. [Google Scholar] [CrossRef]
- Tarelkin, Y.; Hufkens, K.; Hahn, S.; Van den Bulcke, J.; Bastin, J.F.; Ilondea, B.A.; Debeir, O.; Van Acker, J.; Beeckman, H.; De Cannière, C. Wood anatomy variability under contrasted environmental conditions of common deciduous and evergreen species from central African forests. Trees 2019, 33, 893–909. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Tw Number of the Disk | Country | Transect | Number of Tree Rings/Samples |
|---|---|---|---|---|
| Pericopsis elata (4 individuals) | Tw60928 | DRC | 1 | 95 |
| 2 | 93 | |||
| 3 | 95 | |||
| Tw60946 | 1 | 86 | ||
| 2 | 82 | |||
| Tw60948 | 1 | 93 | ||
| 2 | 93 | |||
| Tw60955 | 1 | 82 | ||
| 2 | 101 | |||
| Terminalia superba (14 individuals) | Tw60796 | CDI | 1 | 50 |
| Tw60799 | 1 | 50 | ||
| Tw60802 | 1 | 20 | ||
| Tw60804 | 1 | 29 | ||
| Tw60815 | 1 | 39 | ||
| Tw58841 | DRC | 1 | 25 | |
| Tw58842 | 1 | 20 | ||
| Tw58843 | 1 | 27 | ||
| Tw58844 | 1 | 33 | ||
| Tw58845 | 1 | 20 | ||
| Tw61004 | 1 | 36 | ||
| Tw61005 | 1 | 35 | ||
| Tw61006 | 1 | 50 | ||
| Tw61009 | 1 | 32 |
| Disk | Country | n | %Succes | %Std |
|---|---|---|---|---|
| Tw60796 | CDI | 50 | 94.5 | 4.6 |
| Tw60799 | 50 | 72.0 | 3.6 | |
| Tw60802 | 20 | 43.0 | 2.5 | |
| Tw60804 | 29 | 96.6 | 3.8 | |
| Tw60815 | 39 | 70.3 | 4.2 | |
| Tw58841 | DRC | 25 | 100.0 | 0.0 |
| Tw58842 | 20 | 86.0 | 5.8 | |
| Tw58843 | 27 | 92.6 | 4.1 | |
| Tw58844 | 33 | 98.8 | 2.4 | |
| Tw58845 | 20 | 79.0 | 8.6 | |
| Tw61004 | 36 | 97.8 | 1.1 | |
| Tw61005 | 35 | 100.0 | 0.0 | |
| Tw61006 | 50 | 89.2 | 1.6 | |
| Tw61009 | 32 | 93.1 | 1.3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deklerck, V.; Lancaster, C.A.; Van Acker, J.; Espinoza, E.O.; Van den Bulcke, J.; Beeckman, H. Chemical Fingerprinting of Wood Sampled along a Pith-to-Bark Gradient for Individual Comparison and Provenance Identification. Forests 2020, 11, 107. https://doi.org/10.3390/f11010107
Deklerck V, Lancaster CA, Van Acker J, Espinoza EO, Van den Bulcke J, Beeckman H. Chemical Fingerprinting of Wood Sampled along a Pith-to-Bark Gradient for Individual Comparison and Provenance Identification. Forests. 2020; 11(1):107. https://doi.org/10.3390/f11010107
Chicago/Turabian StyleDeklerck, Victor, Cady A. Lancaster, Joris Van Acker, Edgard O. Espinoza, Jan Van den Bulcke, and Hans Beeckman. 2020. "Chemical Fingerprinting of Wood Sampled along a Pith-to-Bark Gradient for Individual Comparison and Provenance Identification" Forests 11, no. 1: 107. https://doi.org/10.3390/f11010107
APA StyleDeklerck, V., Lancaster, C. A., Van Acker, J., Espinoza, E. O., Van den Bulcke, J., & Beeckman, H. (2020). Chemical Fingerprinting of Wood Sampled along a Pith-to-Bark Gradient for Individual Comparison and Provenance Identification. Forests, 11(1), 107. https://doi.org/10.3390/f11010107