Disturbance History and Dynamics of an Old-Growth Nothofagus Forest in Southern Patagonia

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling
2.3. Dendrochronological Methods
2.4. Analyses
2.4.1. Forest Structure
2.4.2. Forest Dynamics
3. Results
3.1. Forest Structure
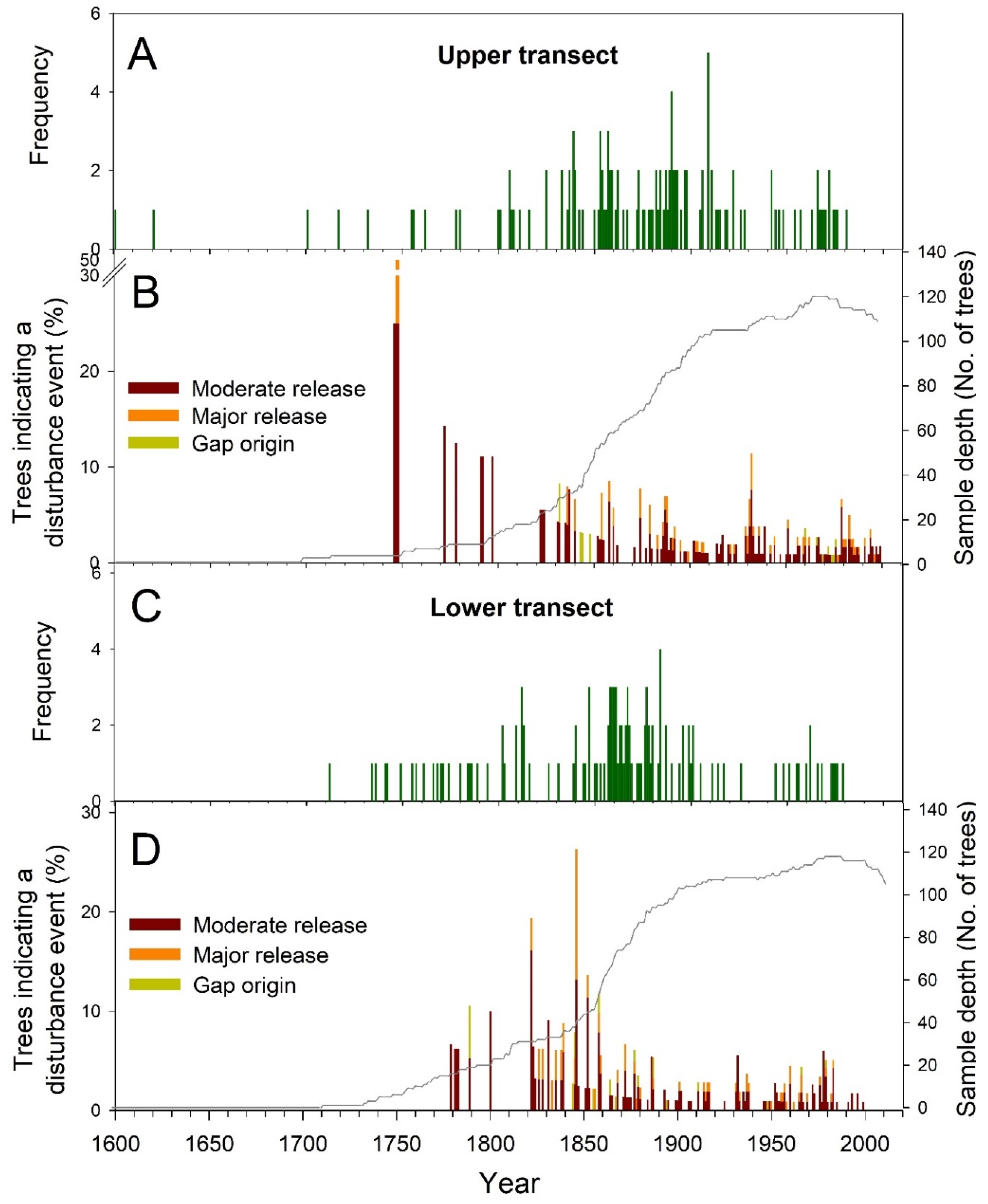
3.2. Forest Dynamics and Disturbance History
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sousa, W.P. The role of disturbance in natural communities. Annu. Rev. Ecol. Syst. 1984, 15, 353–391. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; White, P.S. The Ecology of Natural Disturbance and Patch Dynamics; Academic Press Inc.: Cambridge, MA, USA, 1985; p. 472. [Google Scholar]
- Turner, M.G. Disturbance and landscape dynamics in a changing world. Ecology 2010, 91, 2833–2849. [Google Scholar] [CrossRef] [PubMed]
- Buma, D. Disturbance interactions: Characterization, prediction, and the potential for cascading effects. Ecosphere 2015, 6, 1–15. [Google Scholar] [CrossRef]
- Kimmins, J.P. Forest Ecology. A Foundation for Sustainable Management and Environmental Ethics in Forestry, 3rd ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2004; p. 720. [Google Scholar]
- North, M.P.; Keeton, W.S. Emulating Natural Disturbance Regimes: An Emerging Approach for Sustainable Forest Management. In Patterns and Processes in Forest Landscapes; Lafortezza, R., Sanesi, G., Chen, J., Crow, T.R., Eds.; Springer: Dordrecht, The Netherlands, 2008. [Google Scholar]
- Drake, D.R.; Mueller-Dombois, D. Population development of rainforest trees on a chronosequence of Hawaiian lava flows. Ecology 1993, 74, 1012–1019. [Google Scholar] [CrossRef]
- Walker, L.R.; Wardle, D.A.; Bardgett, R.D.; Clarkson, B.D. The use of chronosequences in studies of ecological succession and soil development. J. Ecol. 2010, 8, 725–736. [Google Scholar] [CrossRef]
- Lutz, J.A.; Halpern, C.B. Tree mortality during early forest development: A long-term study of rates, causes, and consequences. Ecol. Monogr. 2006, 76, 257–275. [Google Scholar] [CrossRef]
- Lorimer, C.G.; Frelich, L.E. A methodology for estimating canopy disturbance frequency and intensity in dense temperate forests. Can. J. For. Res. 1989, 19, 651–663. [Google Scholar] [CrossRef]
- Swetnam, T.W.; Allen, C.D.; Betancourt, J.L. Applied historical ecology using the past to manage for the future. Ecol. Appl. 1999, 9, 1189–1206. [Google Scholar] [CrossRef]
- Splechtna, B.E.; Gratzer, G.; Black, B.A. Disturbance history of a European old-growth mixed-species forest—A spatial dendro-ecological analysis. J. Veg. Sci. 2005, 16, 511–522. [Google Scholar]
- Amoroso, M.M.; David Coates, K.; Astrup, R. Stand recovery and self-organization following large-scale mountain pine beetle induced canopy mortality in northern forests. For. Ecol. Manag. 2013, 310, 300–311. [Google Scholar] [CrossRef]
- Veblen, T.T.; Kitzberger, T.; Villalba, R.; Arturi, M.F.; Frangi, J.L.; Goya, J.F. Nuevos paradigmas en ecología y su influencia sobre el conocimiento de la dinámica de los bosques del sur de Argentina y Chile. In Ecología y Manejo de Bosques de Argentina; Editorial de la Universidad Nacional de La Plata: La Plata, Argentina, 2004; pp. 1–48. [Google Scholar]
- González, M.E.; Amoroso, M.M.; Lara, A.; Veblen, T.T.; Donoso, C.; Kitzberger, T.; Mundo, I.A.; Holz, A.; Casteller, A.; Paritsis, J.; et al. Ecología de disturbios y su influencia en los ecosistemas forestales templados de Chile y Argentina. In Ecología Forestal: Bases para el Manejo Sustentable de los Bosques Nativos; Donoso, C., González, M.E., Lara, A., Donoso, P., Eds.; Marisa Cuneo Editores: Valdivia, Chile, 2014. [Google Scholar]
- Kitzberger, T.; Veblen, T.T. Influences of humans and ENSO on fire history of Austrocedrus chilensis woodlands in northern Patagonia, Argentina. Ecoscience 1997, 4, 508–520. [Google Scholar] [CrossRef]
- Paritsis, J.; Veblen, T.T.; Kitzberger, T. Assessing dendroecological methods to reconstruct defoliator outbreaks on Nothofagus pumilio in northwestern Patagonia, Argentina. Can. J. For. Res. 2009, 39, 1617–1629. [Google Scholar] [CrossRef]
- Casteller, A.; Villalba, R.; Araneo, D.; Stöckli, V. Reconstructing temporal patterns of snow avalanches at Lago del Desierto, southern Patagonian Andes. Cold Reg. Sci. Technol. 2011, 67, 68–78. [Google Scholar] [CrossRef]
- Mundo, I.A.; Barrera, M.D.; Roig, F.A. Testing the utility of Nothofagus pumilio for dating a snow avalanche in Tierra del Fuego. Dendrochronologia 2007, 25, 19–28. [Google Scholar] [CrossRef]
- Veblen, T.T.; Ashton, D.H. Catastrophic influences on the vegetation of the Valdivian Andes, Chile. Vegetation 1978, 36, 147–167. [Google Scholar] [CrossRef]
- Villalba, R.; Veblen, T.T. Influences of large-scale climatic variability on episodic tree mortality in northern Patagonia. Ecology 1998, 79, 2624–2640. [Google Scholar] [CrossRef]
- Kitzberger, T.; Veblen, T.T. Fire-induced changes in northern Patagonian landscapes. Landsc. Ecol. 1999, 14, 1–15. [Google Scholar] [CrossRef]
- Suarez, M.L.; Kitzberger, T. Recruitment patterns following a severe drought: Long-term compositional shifts in Patagonian forests. Can. J. For. Res. 2008, 38, 3002–3010. [Google Scholar] [CrossRef]
- Amoroso, M.M.; Suarez, M.L.; Daniels, L.D. Nothofagus dombeyi regeneration in declining Austrocedrus chilensis forests: Effects of overstory mortality and climatic events. Dendrochronologia 2012, 30, 105–112. [Google Scholar] [CrossRef]
- Amoroso, M.M.; Larson, B.C. Stand development patterns as a consequence of the mortality in Austrocedrus chilensis forests. For. Ecol. Manag. 2010, 259, 1981–1992. [Google Scholar] [CrossRef]
- Suarez, M.L.; Kitzberger, T. Differential effects of climate variability on forest dynamics along a precipitation gradient in northern Patagonia. J. Ecol. 2010, 98, 1023–1034. [Google Scholar] [CrossRef]
- Mundo, I.A.; Villalba, R.; Veblen, T.T.; Kitzberger, T.; Holz, A.; Paritsis, J.; Ripalta, A. Fire history in southern Patagonia: Human and climate influences on fire activity in Nothofagus pumilio forests. Ecosphere 2017, 8, e01932. [Google Scholar] [CrossRef]
- Srur, A.M.; Villalba, R.; Rodríguez-Catón, M.; Amoroso, M.M.; Marcotti, E. Climate and Nothofagus pumilio Establishment at Upper Treelines in the Patagonian Andes. Front. Earth Sci. 2018, 6, 57. [Google Scholar] [CrossRef]
- Rebertus, A.; Veblen, T.T. Structure and tree-fall gap dynamics of old-growth Nothofagus forests in Tierra del Fuego, Argentina. J. Veg. Sci. 1993, 4, 461–654. [Google Scholar] [CrossRef]
- Rebertus, A.; Kitzberger, T.; Veblen, T.; Roovers, L. Blowdown history and landscape patterns in Nothofagus forests in southern Andes, Tierra del Fuego. Ecology 1997, 78, 678–692. [Google Scholar] [CrossRef]
- Puigdefábregas, J.; Gallart, F.; Bianciotto, O.; Allogia, M.; Del Barrio, G. Banded vegetation patterning in a subantarctic forest of Tierra del Fuego, as an outcome on the interaction between wind and tree growth. Acta Oecol. 1999, 20, 135–146. [Google Scholar] [CrossRef]
- Casteller, A.; Villalba, R.; Mayer, A.; Stöckli, V. Reconstrucción espacial y temporal de la ocurrencia de avalanchas de nieve en los Andes patagónicos utilizando técnicas dendrocronológicas. Rev. Chil. Hist. Nat. 2009, 82, 245–264. [Google Scholar] [CrossRef][Green Version]
- Ruiz, M. Reconstrucción de Volteos por Vientos en los Valles del Río Toro, y del Río de las Vueltas (El Chaltén, Provincia de Santa Cruz) a Través del Análisis de los Anillos de Crecimiento de Nothofagus pumilio (Poepp. et Endl.) Krasser (Lenga). Bachelor’s Thesis, Facultad de Ciencias Agrarias, Universidad Nacional de Cuyo, Mendoza, Argentina, 2013. [Google Scholar]
- Nagel, T.A.; Levanic, T.; Diaci, J. A dendroecological reconstruction of disturbance in an old-growth Fagus-Abies forest in Slovenia. Ann. For. Sci. 2007, 64, 891–897. [Google Scholar] [CrossRef]
- Firm, D.; Nagel, T.A.; Diaci, J. Disturbance history and dynamics of and old-growth mixed species mountain forest in the Slovenian Alps. For. Ecol. Manag. 2009, 257, 1893–1901. [Google Scholar] [CrossRef]
- Svoboda, M.; Janda, P.; Nagel, T.A.; Fraver, S.; Rejzek, J.; Bače, R. Disturbance history of an old-growth sub-alpine Picea abies stand in the Bohemian Forest, Czech Republic. J. Veg. Sci. 2012, 23, 86–97. [Google Scholar] [CrossRef]
- Villegas, D.C.; Pereyra, F.X.; Ferrer, J.A. Suelos, factores de formación y procesos pedogenéticos en el centro oeste de la provincia de Santa Cruz. Rev. Asoc. Geol. Argent. 2007, 62, 366–374. [Google Scholar]
- Cottam, G.; Curtis, J.T. The use of distance measures in phytosociological sampling. Ecology 1956, 37, 451–460. [Google Scholar] [CrossRef]
- Stockes, M.A.; Smiley, T.L. An Introduction to Tree-Ring Dating; University of Chicago Press: Chicago, IL, USA, 1968; p. 73. [Google Scholar]
- Holmes, R.L. Computer assisted quality control in tree-ring dating and measurement. Tree Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Cook, E.R.; Krusic, P.J. ARSTAN_41: A Tree-Ring Standardi-Zation Program Based on Detrending and Autoregressive Time Seriesmodeling, with Interactive Graphics; Tree-Ring Laboratory, La-mont Doherty Earth Observatory of Columbia University: New York, NY, USA, 2006. [Google Scholar]
- Duncan, R.P. An evaluation of errors in tree age estimates based on increment cores in kahikatea (Dacrycarpus dacrydioides). N. Z. Nat. Sci. 1989, 16, 31–37. [Google Scholar]
- Di Rienzo, J.A.; Balzarini, M.; Gonzalez, L.; Casanoves, F.; Tablada, M. InfoStat Software Estadístico, Estadística y Biometría y de Diseño de Experimentos; Universidad Nacional de Córdoba: Córdoba, Argentina, 2013. [Google Scholar]
- Fraver, S.; White, A.S. Identifying growth releases in dendrochronological studies of forest disturbance. Can. J. For. Res. 2005, 35, 1648–1656. [Google Scholar] [CrossRef]
- Nowacki, G.J.; Abrams, M.D. Radial-growth averaging criteria for reconstructing disturbance histories from presettlement-origin oaks. Ecol. Monog. 1997, 67, 225–249. [Google Scholar] [CrossRef]
- Holmes, R.L. Program JOLTS: Finding Growth Surges or Suppressions in Trees; Laboratory of Tree-Ring Research, University of Arizona: Tucson, AZ, USA, 1999. [Google Scholar]
- Leak, W.B. Comparison of standard and actual tree-growth for deciduous and coniferous species in New Hampshire. Can. J. For. Res. 1987, 17, 1297–1300. [Google Scholar] [CrossRef]
- Rubino, D.L.; McCarthy, B.C. Comparative analysis of dendroecological methods used to assess disturbance events. Dendrochronologia 2004, 21, 97–115. [Google Scholar] [CrossRef]
- Veblen, T.T. Forest development in tree-fall gaps in the temperate rain forest of Chile. Nat. Geogr. Res. 1985, 1, 161–184. [Google Scholar]
- Veblen, T.T. Nothofagus regeneration in treefall gaps in northern Patagonia. Can. J. For. Res. 1989, 19, 365–371. [Google Scholar] [CrossRef]
- Veblen, T.T.; Donoso, C.; Kitzberger, T.; Rebertus, A.J. Ecology of Southern Chilean and Argentinean Nothofagus forests. In The Ecology and Biogeography of Nothofagus Forests; Veblen, T.T., Hill, R.S., Read, J., Eds.; Yale University Press: New Haven, CT, USA, 1996; pp. 293–353. [Google Scholar]
- Zielonka, T.; Malcher, P. The dynamics of a mountain mixed forest under wind disturbance in the Tatra Mountains, central Europe: A dendroecological reconstruction. Can. J. For. Res. 2009, 39, 2215–2223. [Google Scholar] [CrossRef]
- Mitchell, S.J. Wind as a natural disturbance agent in forests: A synthesis. Forestry 2013, 86, 147–157. [Google Scholar] [CrossRef]
- Kajimoto, T.; Daimaru, H.; Okamoto, T. Effects of snow avalanche disturbance on regeneration of subalpine Abies mariesii forest, Northern Japan. Arct. Antarct. Alp. Res. 2004, 36, 436–445. [Google Scholar] [CrossRef]
- Bebi, P.; Kulakowski, D.; Rixen, C. Snow avalanche disturbances in forest ecosystems-state of research and implications for management. For. Ecol. Manag. 2009, 257, 1883–1892. [Google Scholar] [CrossRef]
- Bergeron, Y.; Denneler, B.; Charron, D.; Girardin, M.P. Using dendrochronology to reconstruct distrubance and forest dynamics around Lake Duparquet, northwestern Quebec. Dendrochronologia 2002, 20, 175–189. [Google Scholar] [CrossRef]
- Axelson, J.N.; Alfaro, R.I.; Hawkes, B.C. Influence of fire and mountain pine beetle on the dynamics of lodgepole pine stands in British Columbia, Canada. For. Ecol. Manag. 2009, 257, 1874–1882. [Google Scholar] [CrossRef]
- Amoroso, M.M.; Daniels, L.D.; Bataineh, M.; Andison, D.W. Evidence of Mixed-Severity Fires in the Foothills of the Rocky Mountains of West-Central Alberta, Canada. For. Ecol. Manag. 2011, 262, 2240–2249. [Google Scholar] [CrossRef]
- Svoboda, M.; Fraver, S.; Janda, P.; Bače, R.; Zenáhlíková, J. Natural development and regeneration of a Central European montane spruce forest. For. Ecol. Manag. 2010, 260, 707–714. [Google Scholar] [CrossRef]
- Martínez Pastur, G.; Fernández, C.; Peri, P. Variación de parámetros estructurales y de composición del sotobosque para bosques de Nothofagus pumilio en relación con gradientes ambientales indirectos. Cienc. For. 1994, 9, 11–22. [Google Scholar]
- Barrera, M.D.; Frangi, J.I.; Richter, L.I.; Perdomo, M.H.; Pinedo, L.B. Structural and functional changes in Nothofagus pumilio forests along an altitudinal gradient in Tierra del Fuego, Argentina. J. Veg. Sci. 2000, 11, 179–188. [Google Scholar] [CrossRef]
- Massaccesi, G.; Roig, F.A.; Martínez Pastur, G.J.; Barrera, M.D. Growth patterns of Nothofagus pumilio trees along altitudinal gradients in Tierra del Fuego, Argentina. Trees 2008, 22, 245–255. [Google Scholar] [CrossRef]
- Coomes, D.A.; Allen, R.B. Effects of size, competition and altitude on tree growth. J. Ecol. 2007, 95, 1084–1097. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Height (m) | Diameter (cm) | Age (Year) | Overstory Trees (n°/ha) | Understory Trees (n°/ha) | Basal Area (m2/ha) | |
|---|---|---|---|---|---|---|
| Upper transect | 5.3 (± 2.6) | 16 (± 11.9) | 136 (± 61.4) | 6455 (± 8063) | (± 31,668) | 75.1 (± 58.7) |
| Lower transect | 8.2 (± 3.2) | 24.8 (± 14.3) | 151 (± 59.3) | 4580 (± 11,222) | 3437 (± 12,854) | 85.9 (± 67.8) |
| Mean | 6.7 | 20.4 | 143.5 | 5517 | 7601 | 80.5 |
| Moderate Release (PGC 50%–99%) | Major Release (PGC > 100%) | |||||
|---|---|---|---|---|---|---|
| % Trees | Mean (Year) | Maximum (Year) | % Trees | Mean (Year) | Maximum (Year) | |
| Upper transect | 74 | 6,4 | 22 | 26 | 6,1 | 13 |
| Lower transect | 76 | 6,7 | 18 | 24 | 5,7 | 13 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amoroso, M.M.; Blazina, A.P. Disturbance History and Dynamics of an Old-Growth Nothofagus Forest in Southern Patagonia. Forests 2020, 11, 101. https://doi.org/10.3390/f11010101
Amoroso MM, Blazina AP. Disturbance History and Dynamics of an Old-Growth Nothofagus Forest in Southern Patagonia. Forests. 2020; 11(1):101. https://doi.org/10.3390/f11010101
Chicago/Turabian StyleAmoroso, Mariano Martín, and Ana Paula Blazina. 2020. "Disturbance History and Dynamics of an Old-Growth Nothofagus Forest in Southern Patagonia" Forests 11, no. 1: 101. https://doi.org/10.3390/f11010101
APA StyleAmoroso, M. M., & Blazina, A. P. (2020). Disturbance History and Dynamics of an Old-Growth Nothofagus Forest in Southern Patagonia. Forests, 11(1), 101. https://doi.org/10.3390/f11010101