The Effect of Tree Crown Allometry on Community Dynamics in Mixed-Species Stands versus Monocultures. A Review and Perspectives for Modeling and Silvicultural Regulation


Abstract
1. Introduction
The Relevance of Crown Size for Tree Growth and Stand Dynamics
- (i.)
- We first review the crown allometry of primary tree species in mono-specific forest stands.
- (ii.)
- Second, we show how tree species mixing can modify the individual tree allometry.
- (iii.)
- Third, we show how the individual tree crown allometry translates to species-specific stand density and self-thinning at the stand level in mixed-species stands compared with monocultures.
- (iv.)
- Fourth, we use equivalence coefficients for revelation and quantification species specific differences in density and growing space requirement and for converting them into a common currency.
- (v.)
- Fifth, we show how the species-specific crown allometry should be considered when regulating the density and mixing proportions in forest stands.
2. Materials and Methods
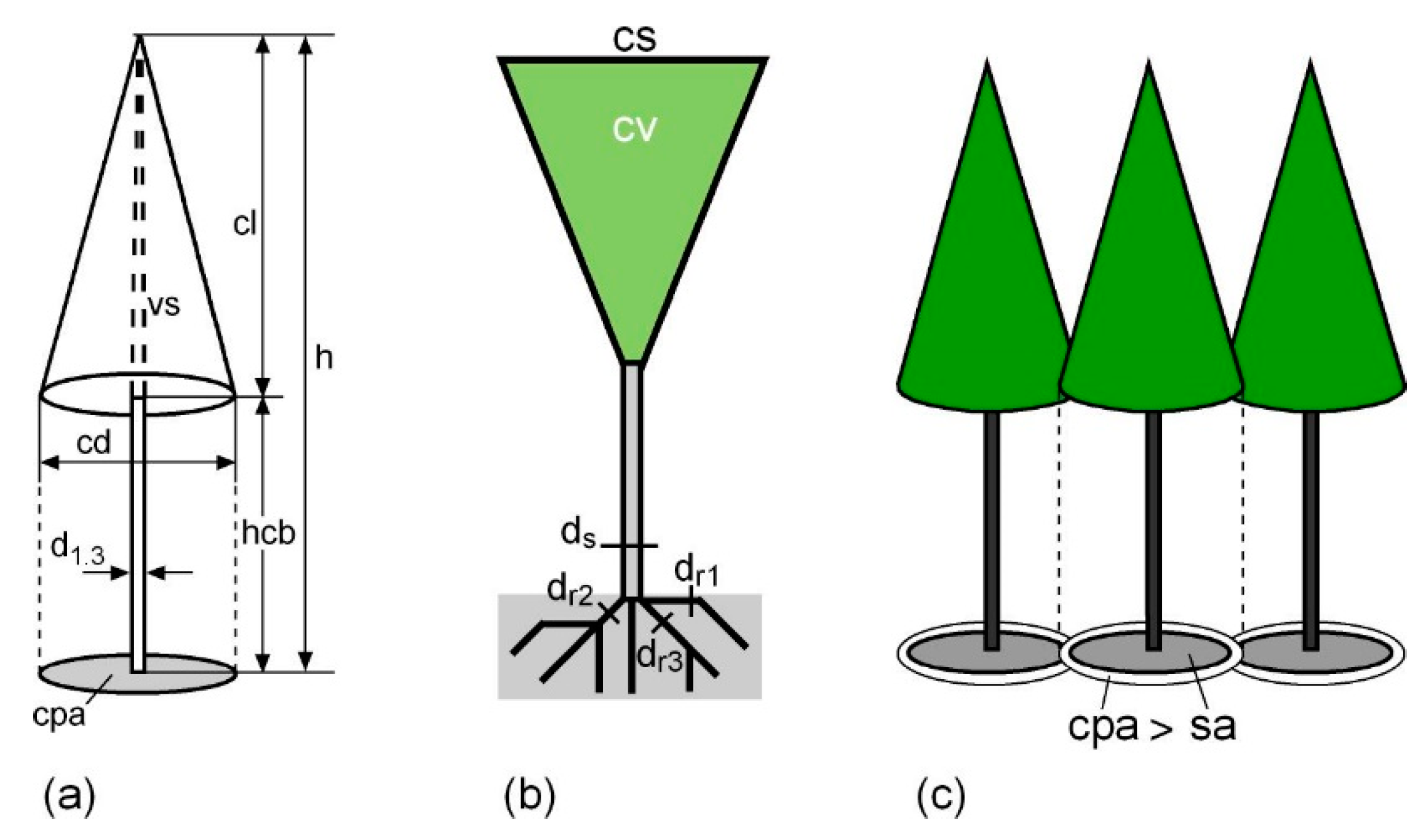
2.1. Dendrometrical Characteristics of Tree Crowns
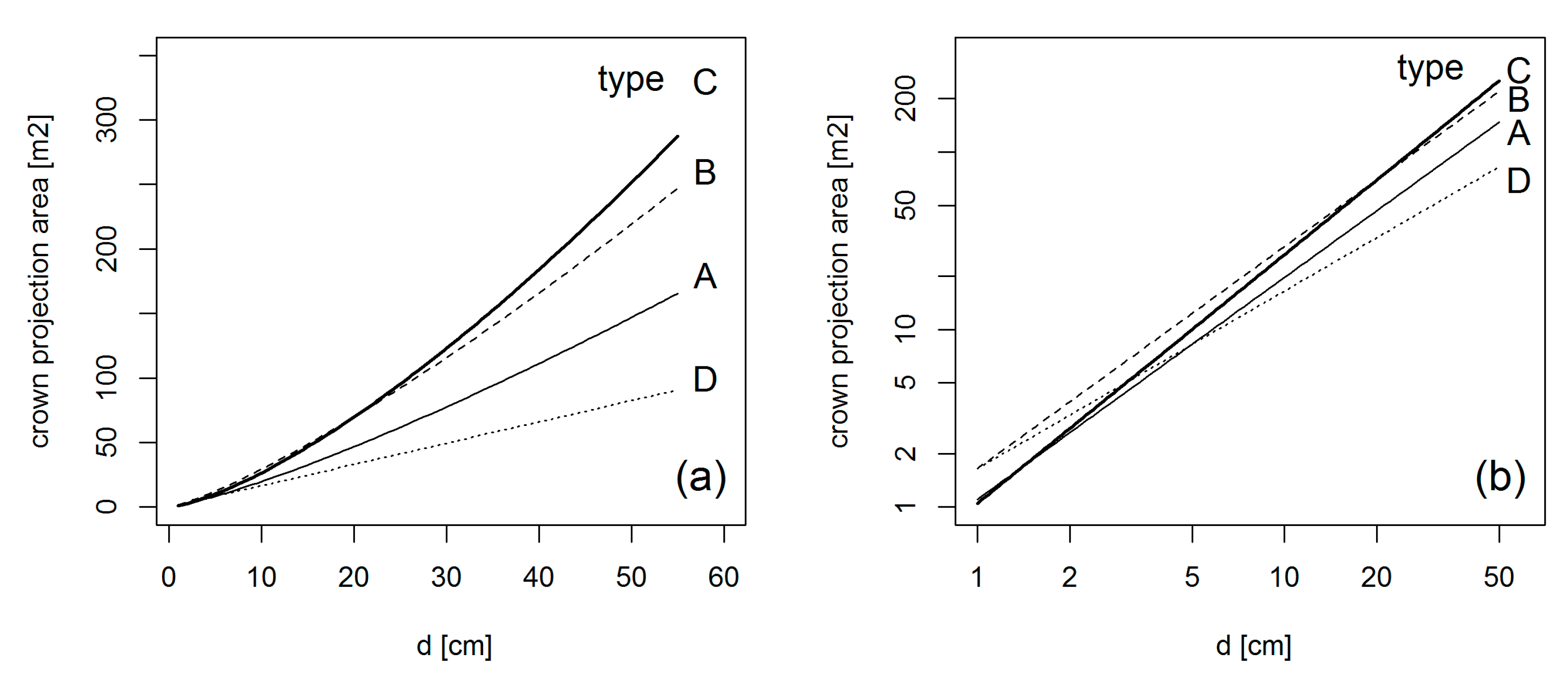
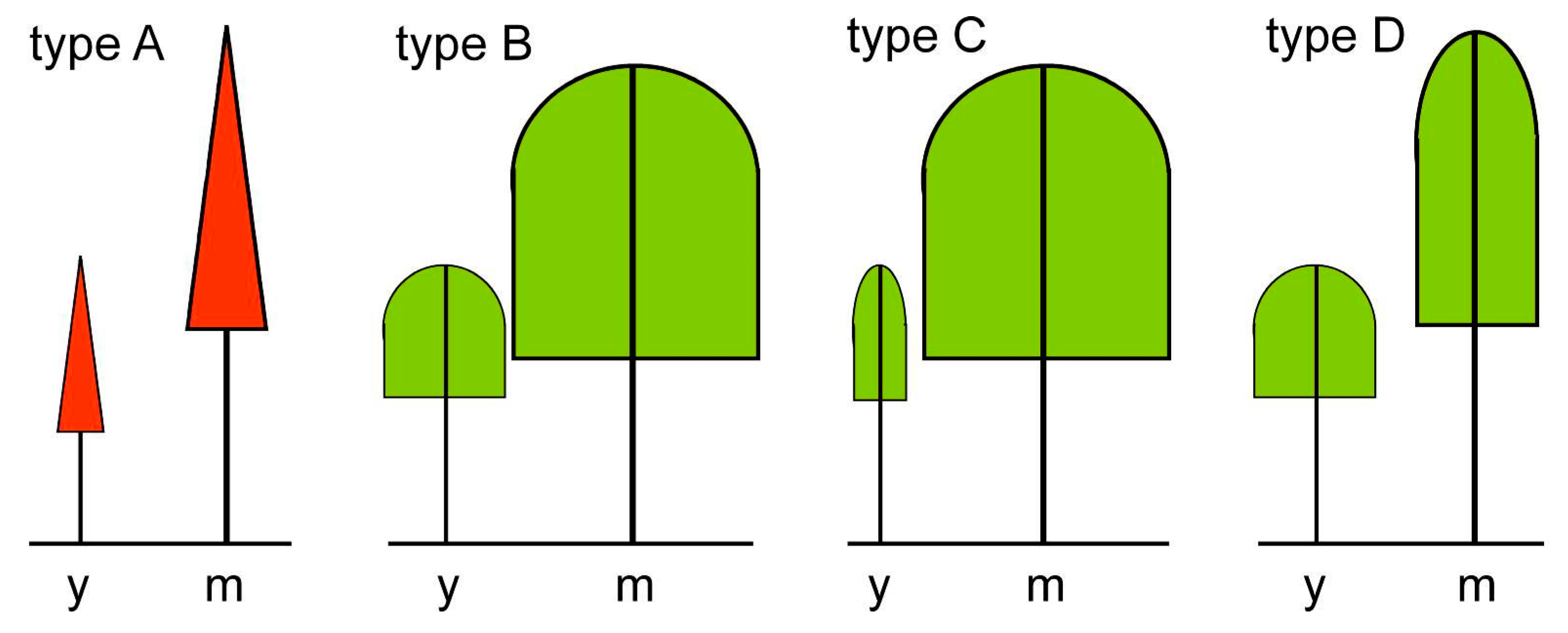
2.2. Basis Types of Crown Allometry
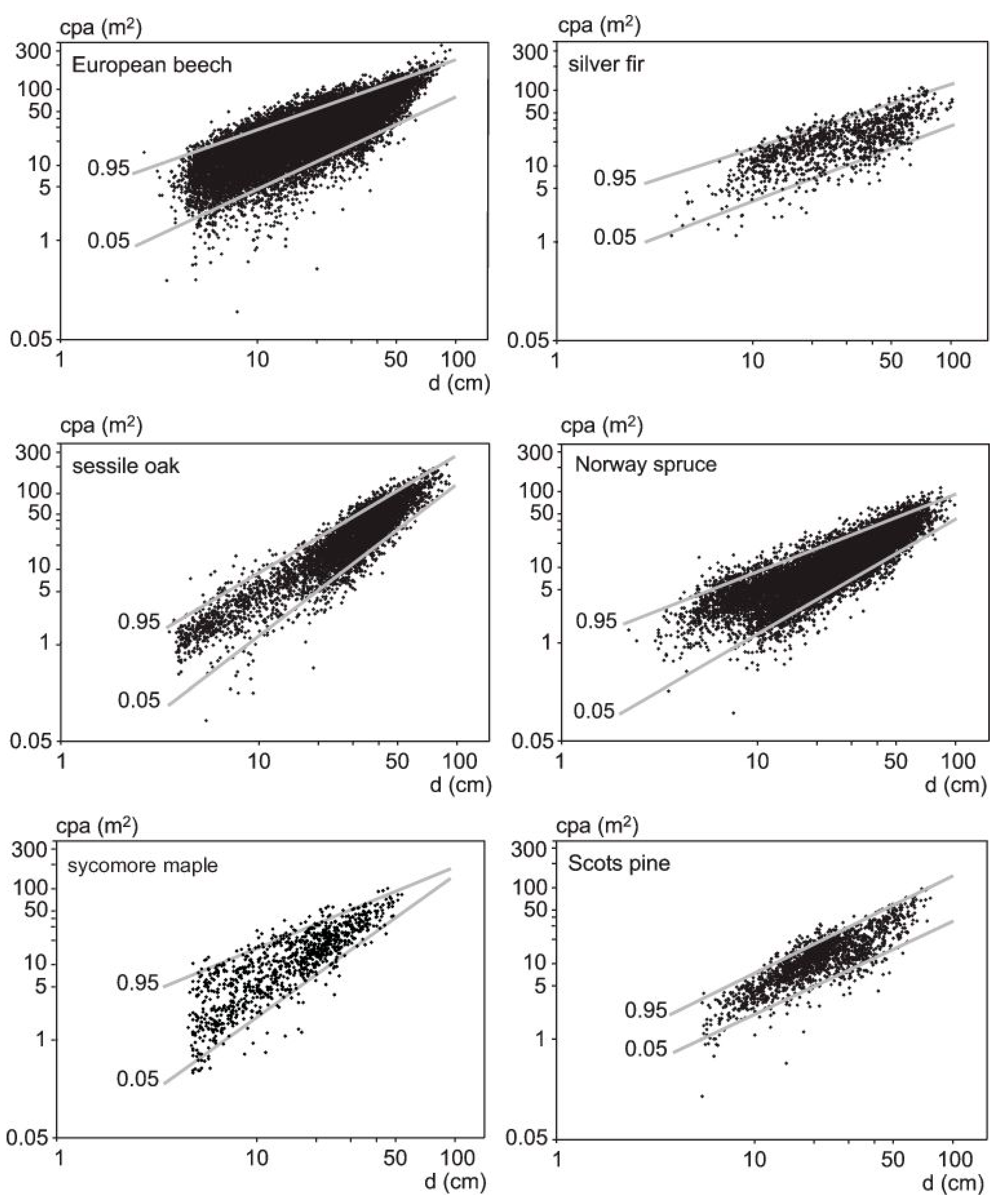
2.3. Crown Data
2.4. Statistical Models
3. Tree Crown Allometry in Mixed versus Monospecific Stands
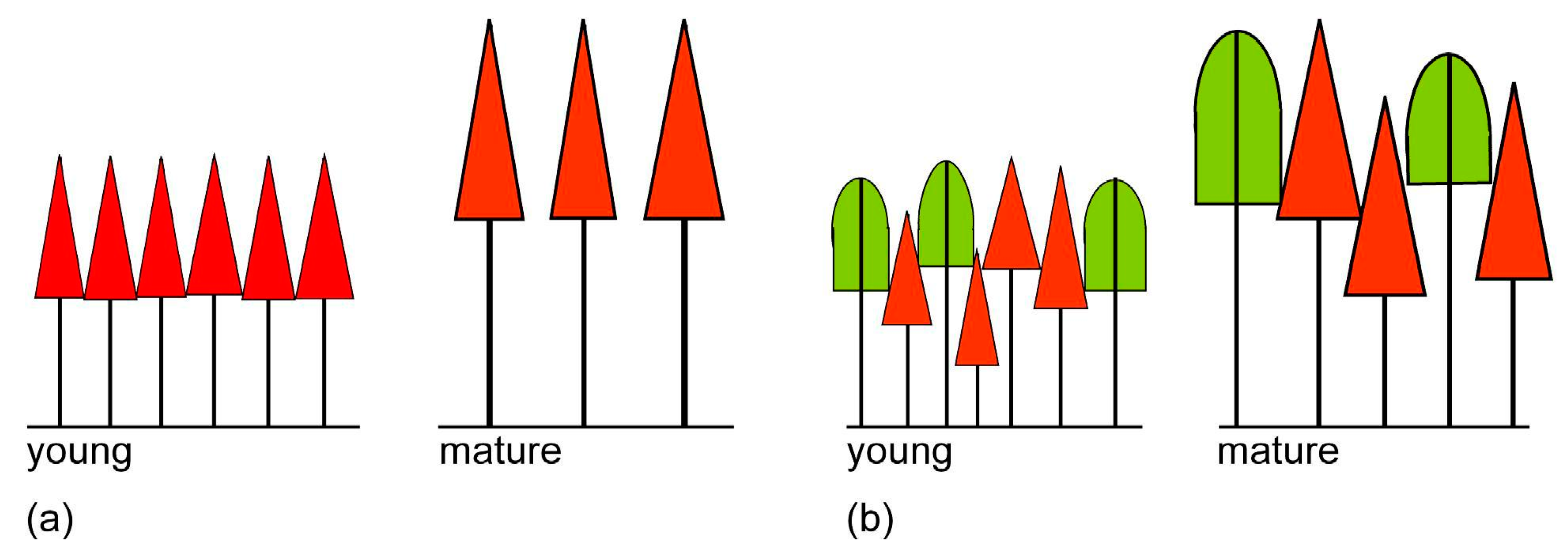
3.1. Allometric Plasticity
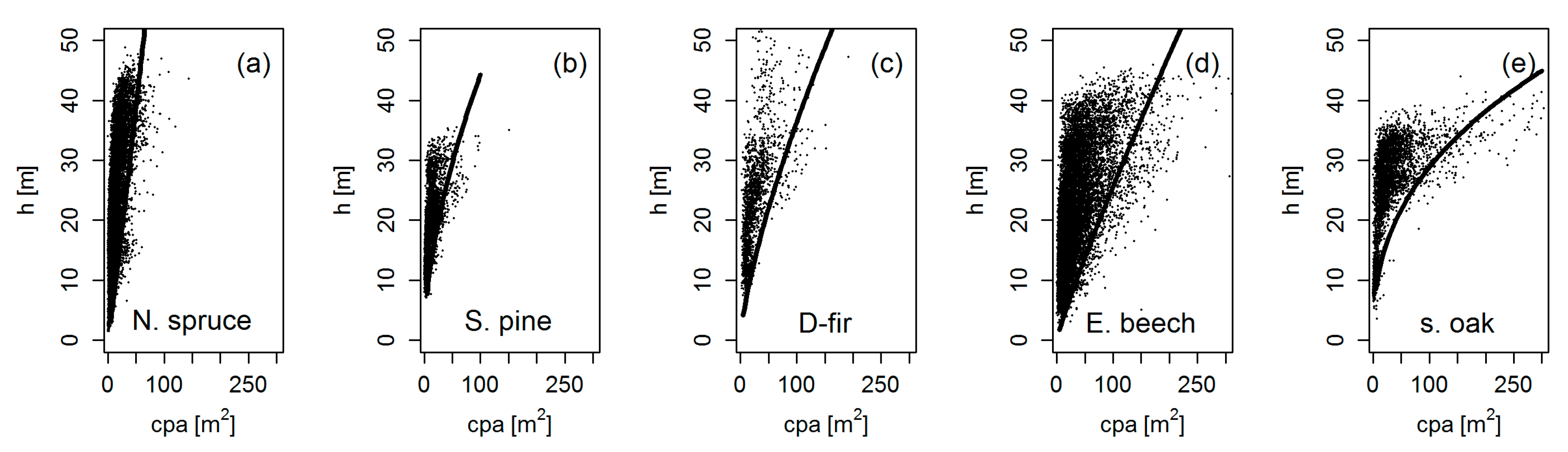
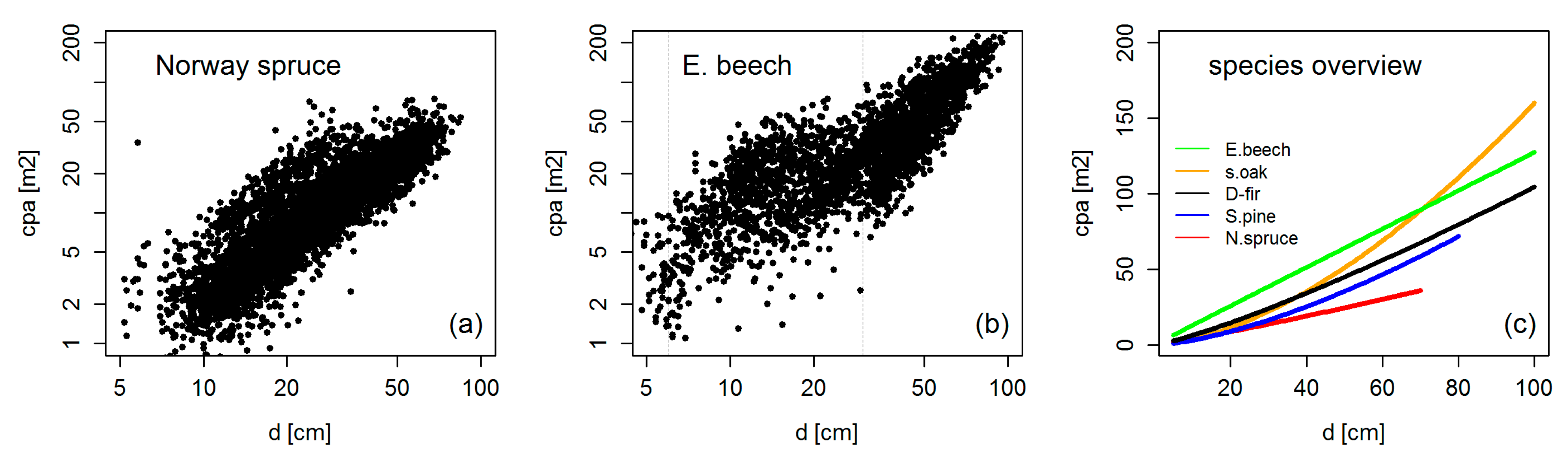
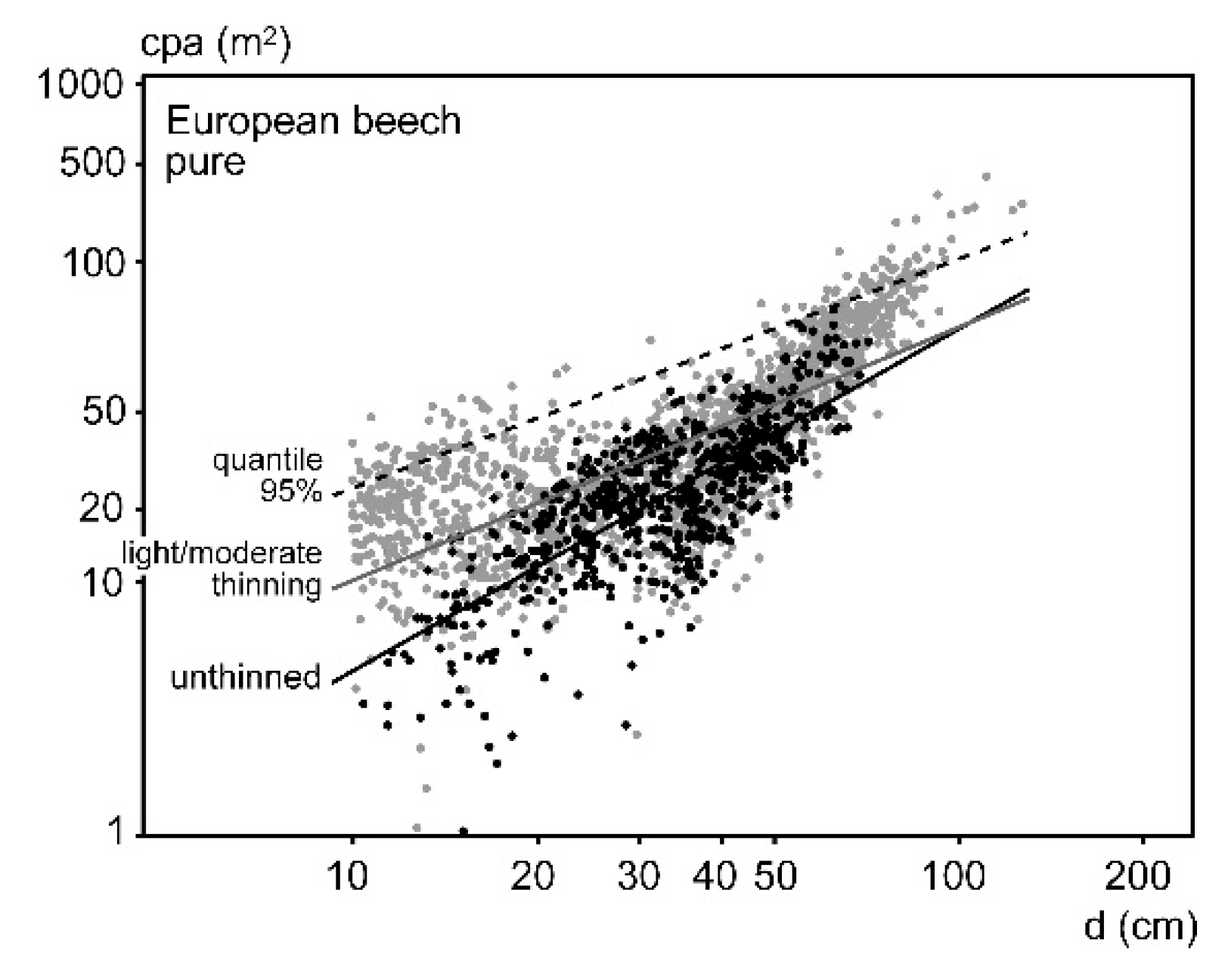
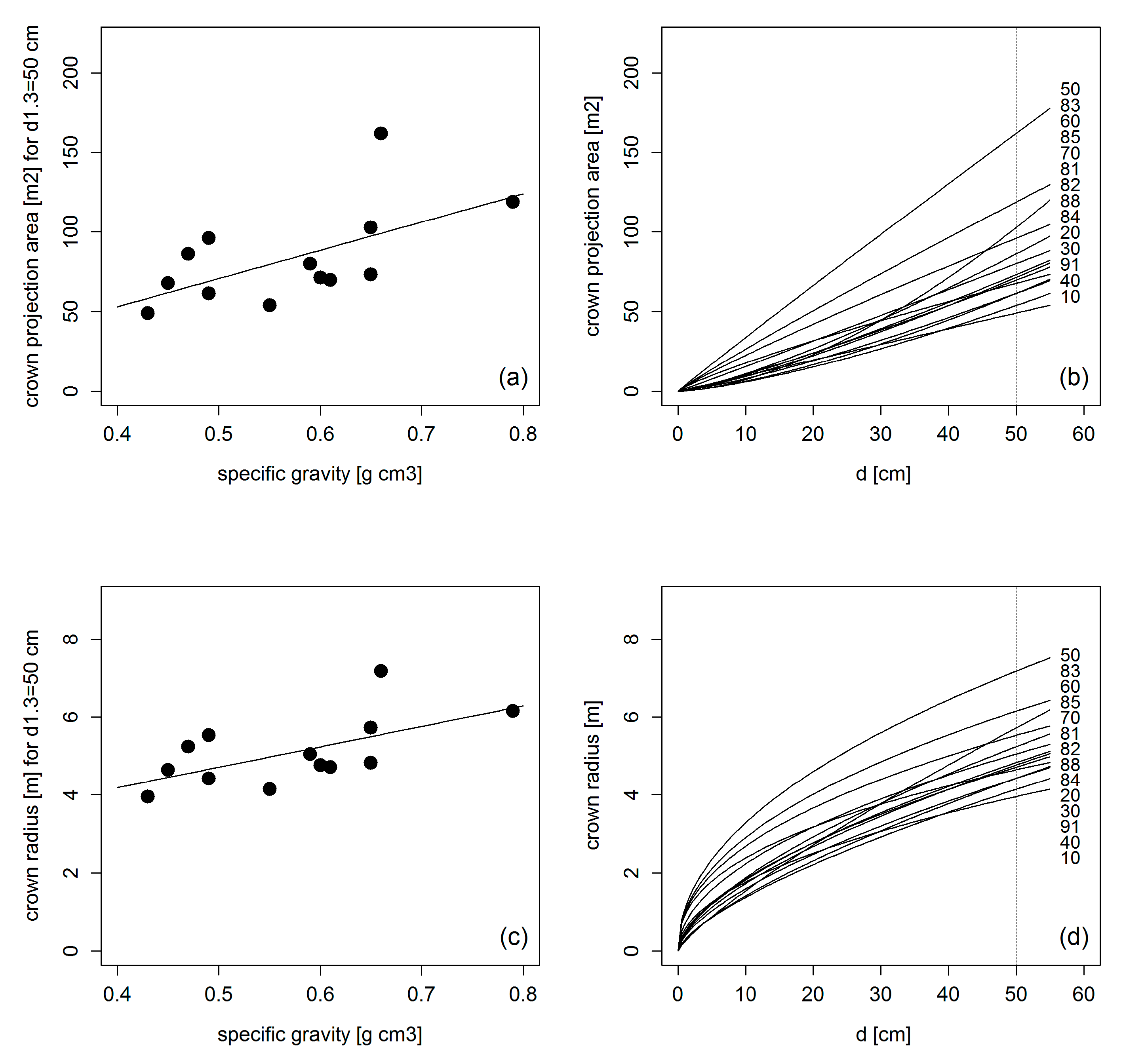
3.2. Crown Allometry in Mono-Specific Stands
3.3. Crown Widening by Stand Density Reduction
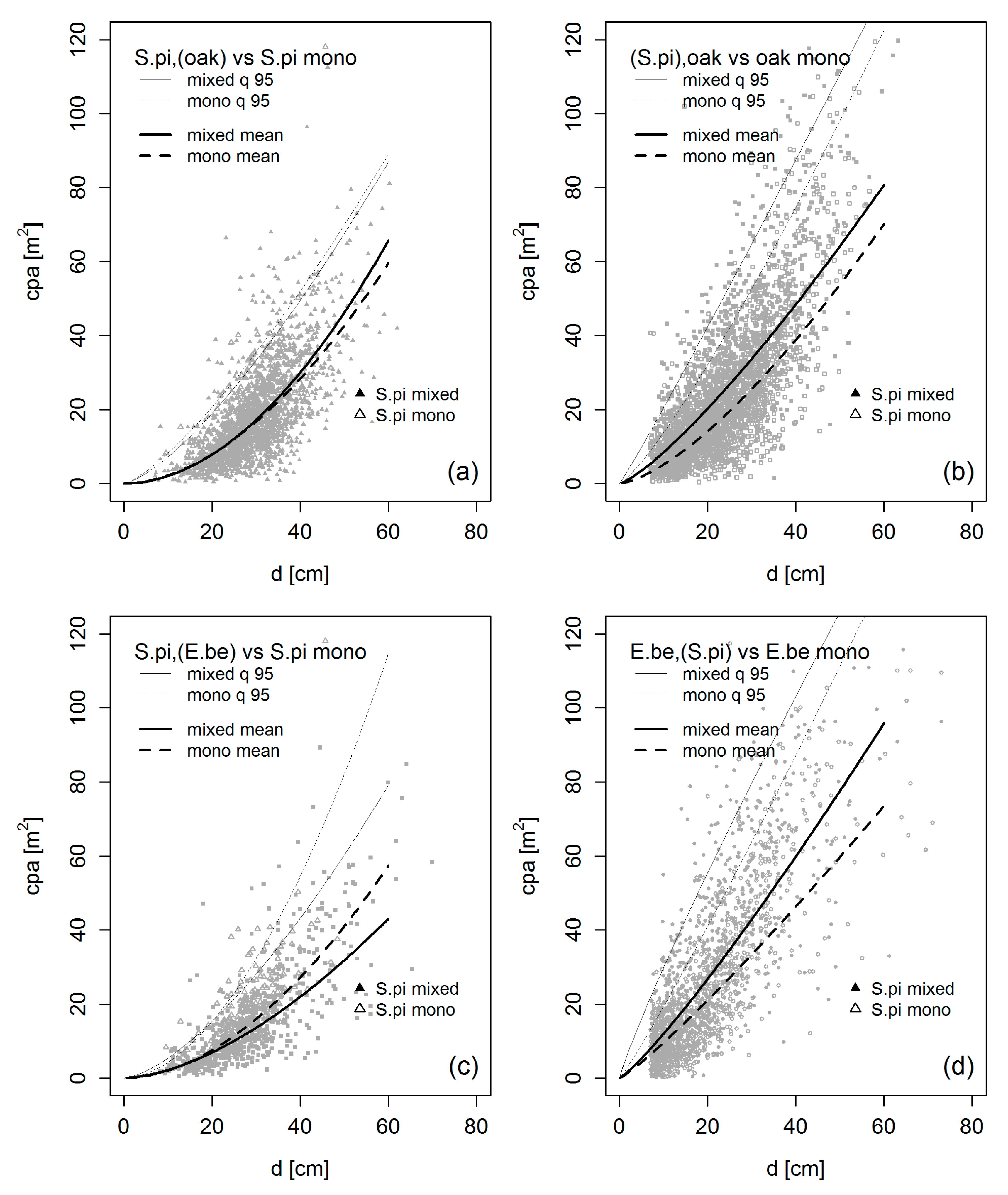
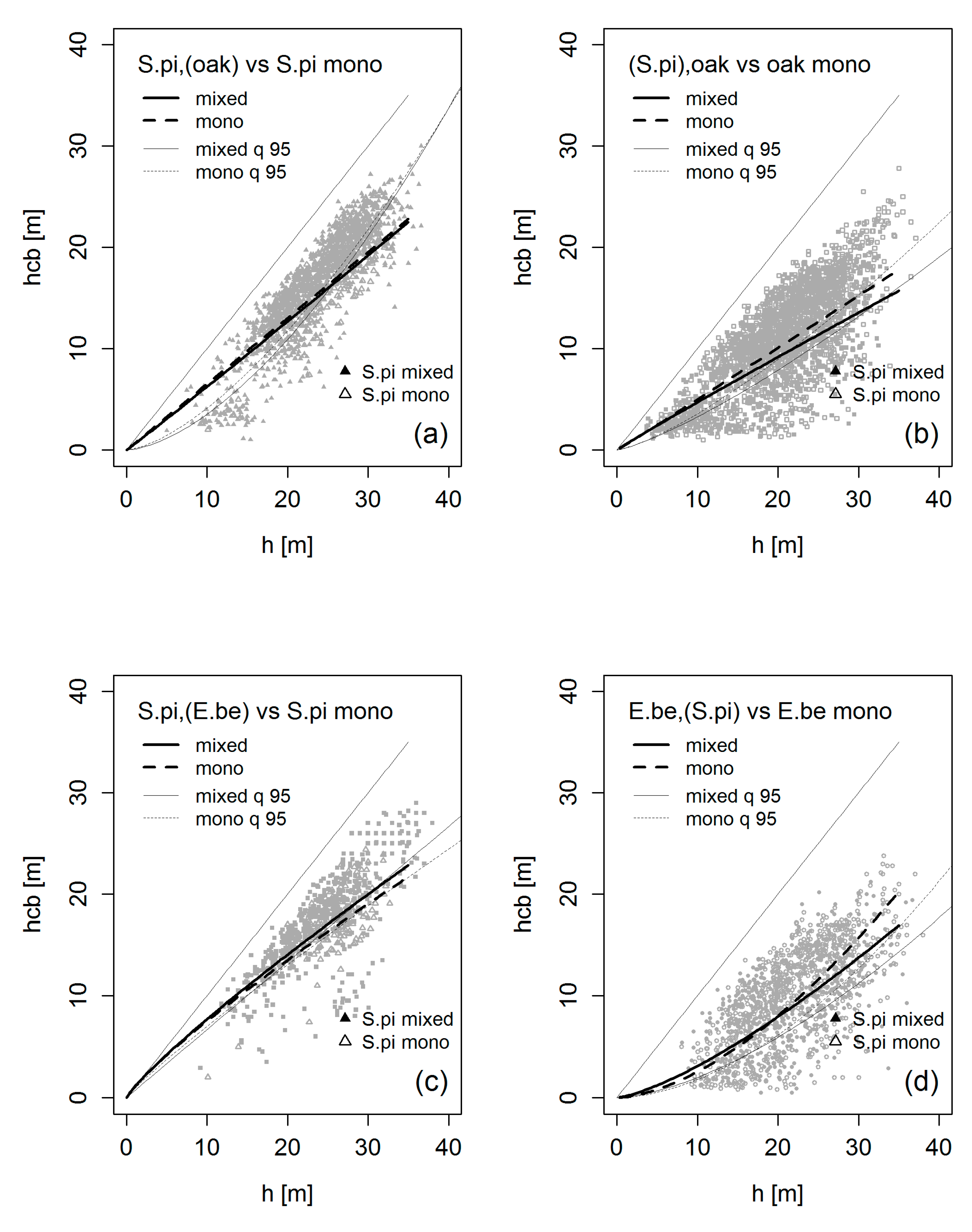
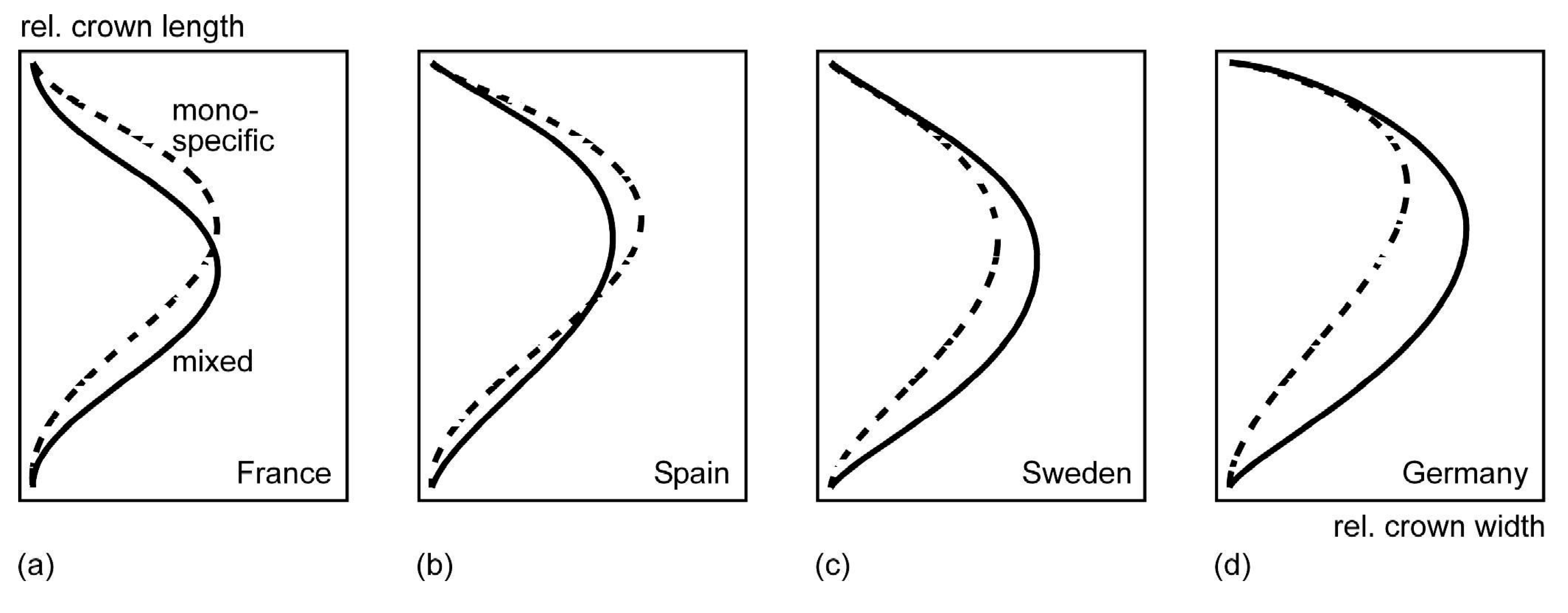
3.4. Crown Allometry in Mixed-Species Stands
4. Tracing the Effect of the Individual Tree Allometry to the Stand Level
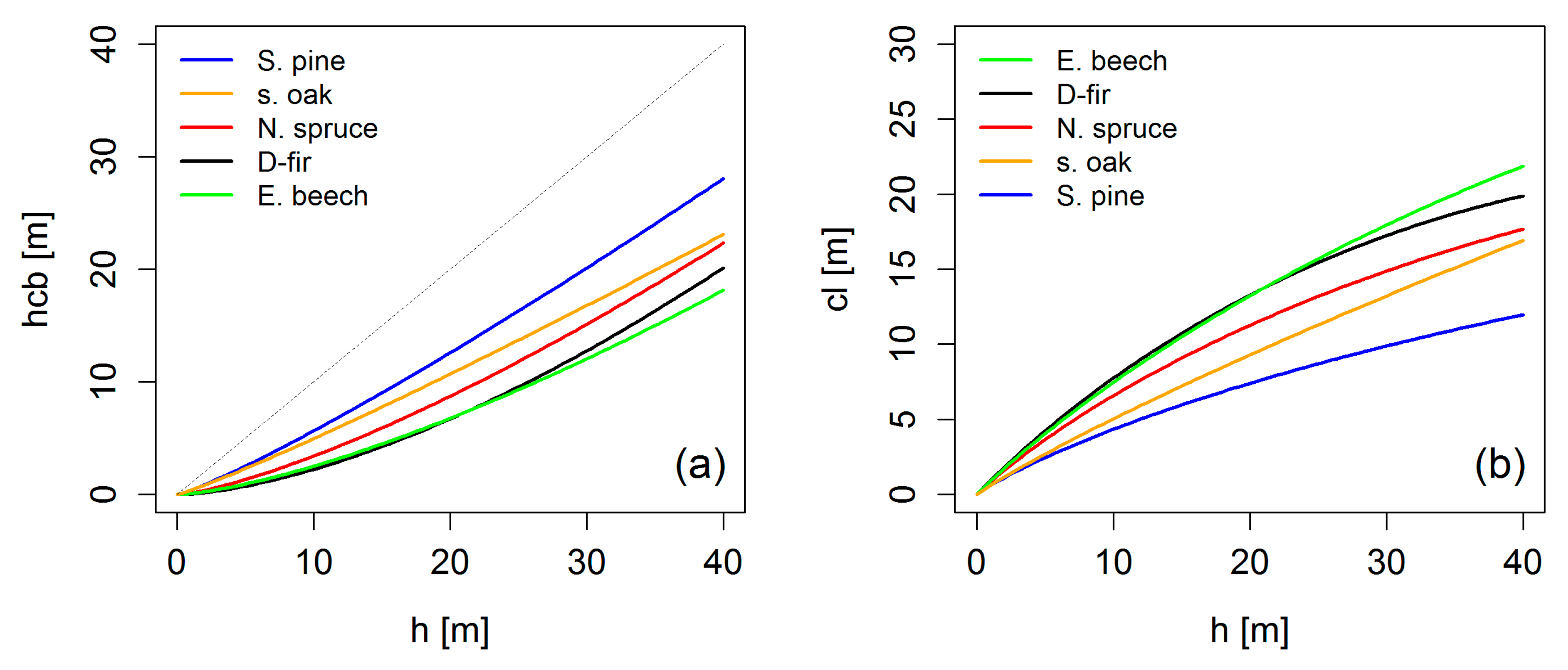
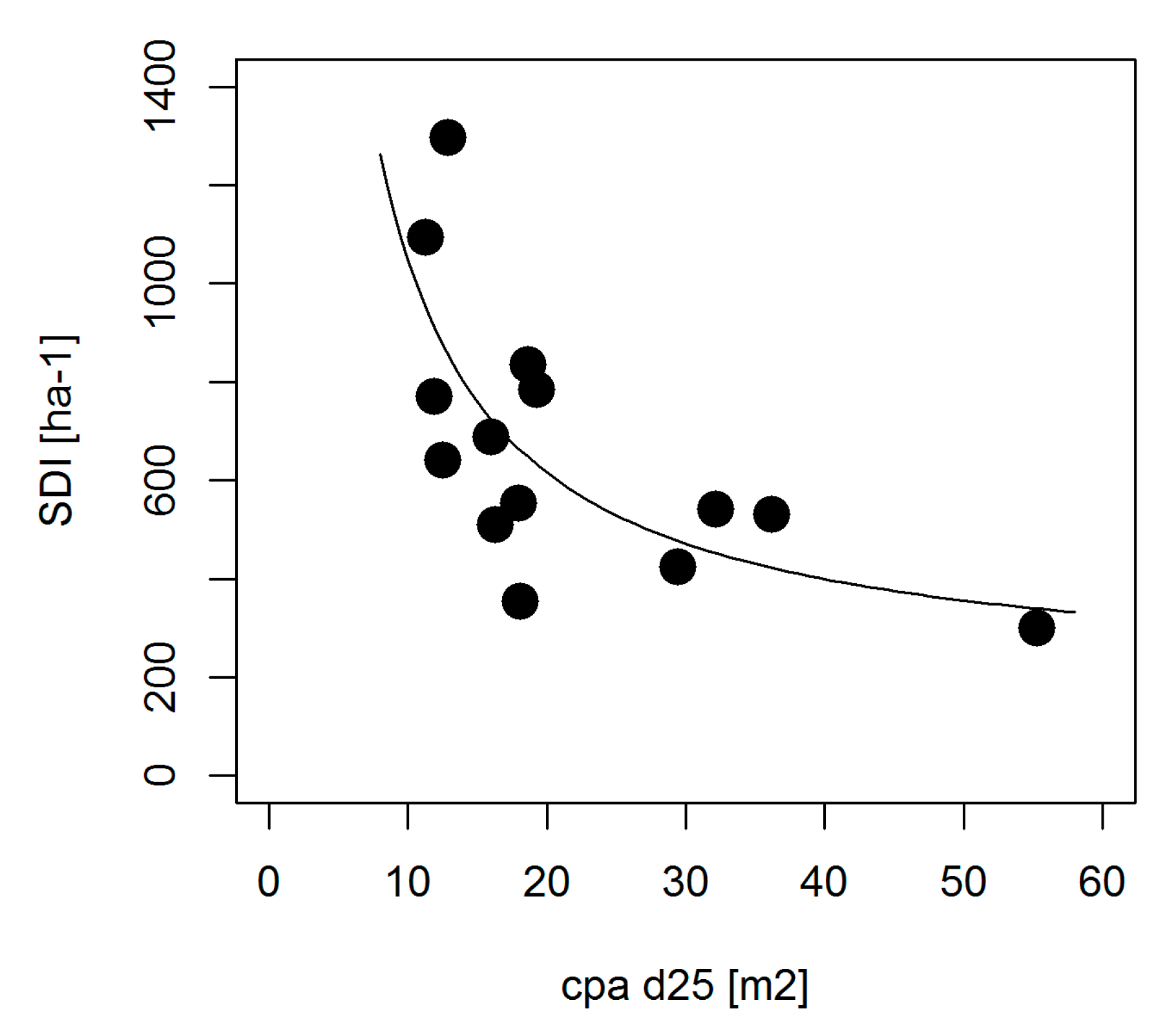
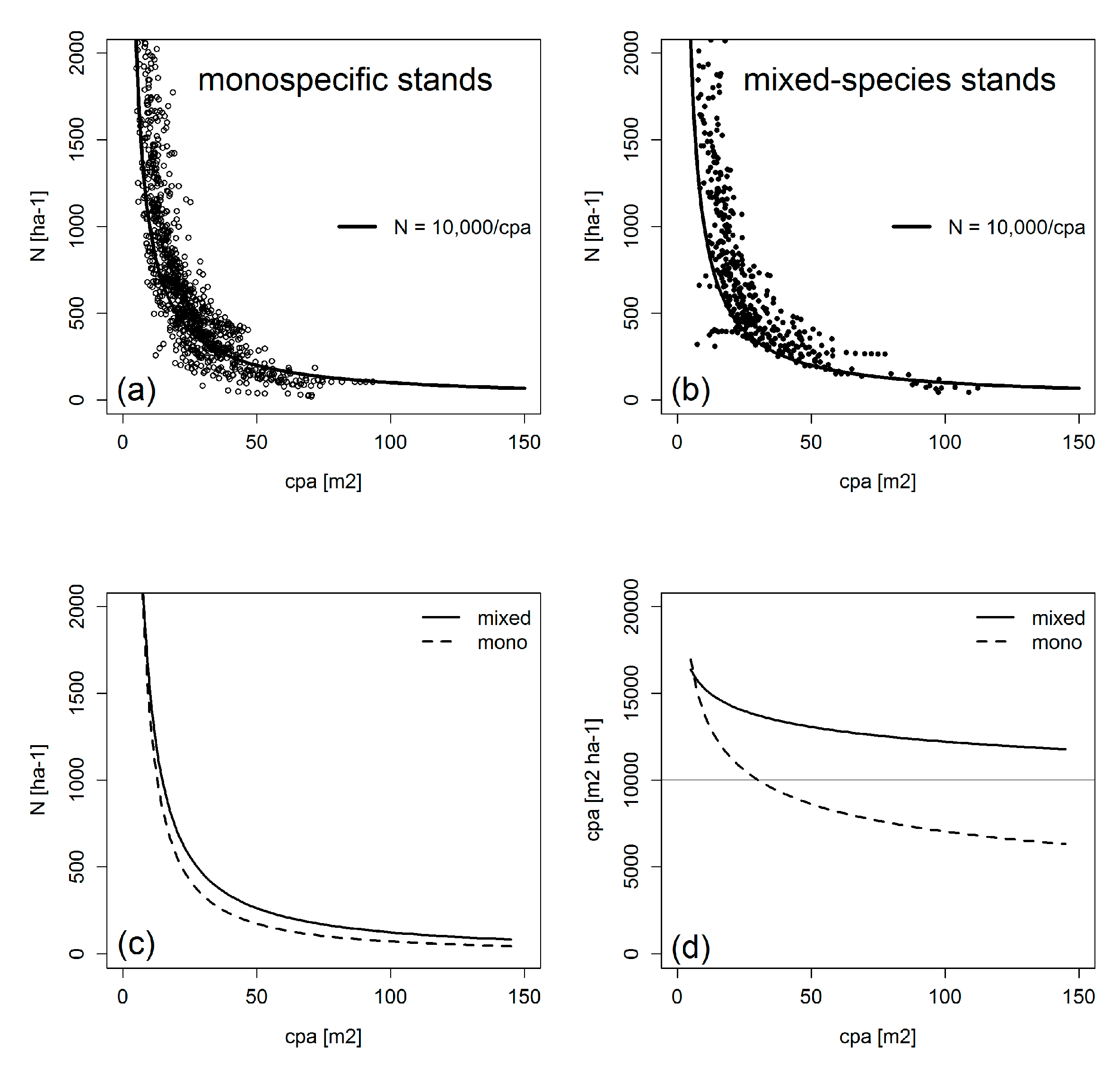
4.1. Crowns and Self-Thinning in Mono-Specific Stands
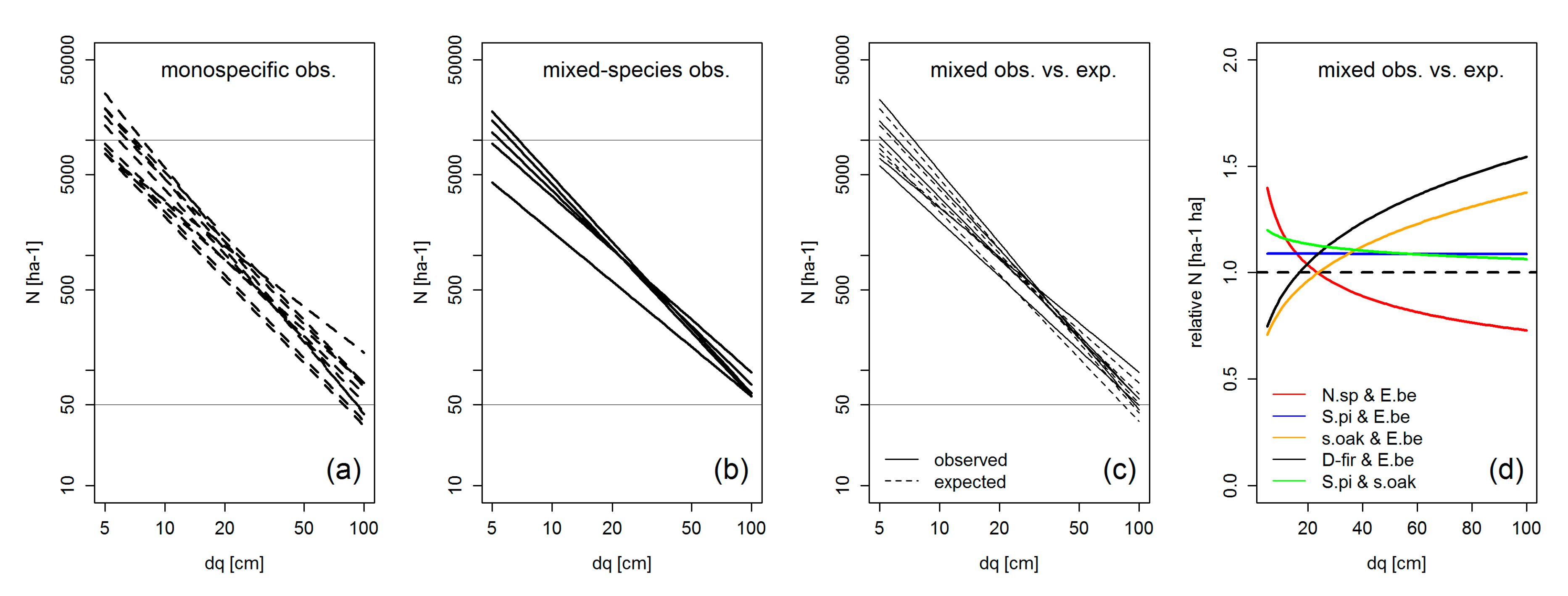
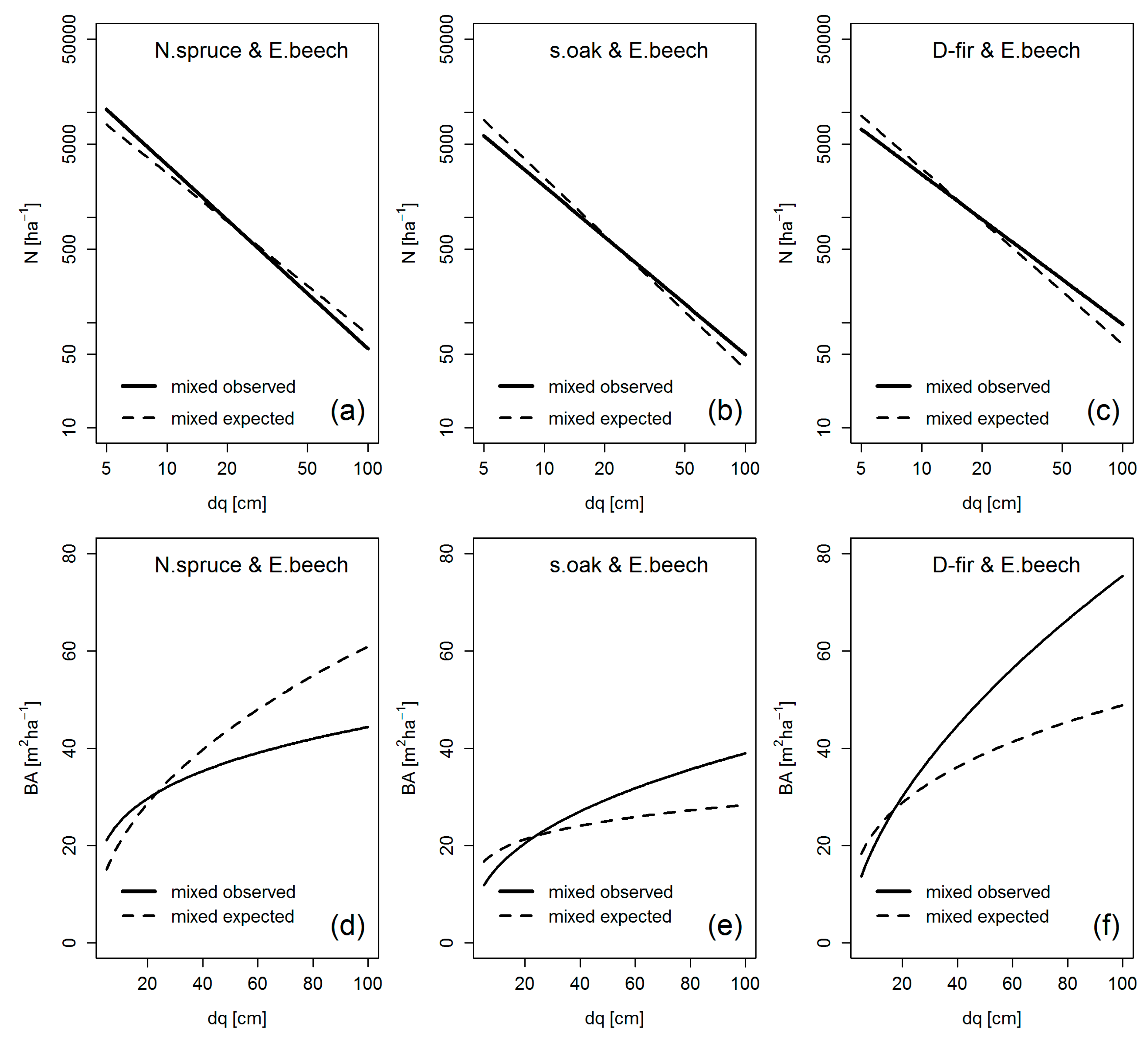
4.2. Crown Allometry and Stand Density in Mixed-Species Stands
5. Discussion
5.1. Crown Allometry and Stand Density in Mixed versus Monospecific Stands
5.2. Tracing Crown Allometry to the Stand Level
5.3. Consequences for Modelling
5.4. Consequences for Stand Density Regulation
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- White, E.P.; Ernest, S.M.; Kerkhoff, A.J.; Enquist, B.J. Relationships between body size and abundance in ecology. Trends Ecol. Evol. 2007, 22, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Knoke, T.; Ammer, C.; Stimm, B.; Mosandl, R. Admixing broadleaved to coniferous tree species: A review on yield, ecological stability and economics. Eur. J. For. Res. 2008, 127, 89–101. [Google Scholar] [CrossRef]
- Pretzsch, H.; Rais, A. Wood quality in complex forests versus even-aged monocultures: Review and perspectives. Wood Sci. Technol. 2016, 50, 845–880. [Google Scholar] [CrossRef]
- Enquist, B.J.; Brown, J.H.; West, G.B. Allometric scaling of plant energetics and population density. Nature 1998, 395, 163–165. [Google Scholar] [CrossRef]
- Barbeito, I.; Dassot, M.; Bayer, D.; Collet, C.; Drössler, L.; Löf, M.; Pretzsch, H. Terrestrial laser scanning reveals differences in crown structure of Fagus sylvatica in mixed vs. pure European forests. For. Ecol. Manag. 2017, 405, 381–390. [Google Scholar] [CrossRef]
- Bayer, D.; Seifert, S.; Pretzsch, H. Structural crown properties of Norway spruce (Picea abies [L.] Karst.) and European beech (Fagus sylvatica [L.]) in mixed versus pure stands revealed by terrestrial laser scanning. Trees 2013, 27, 1035–1047. [Google Scholar] [CrossRef]
- Olivier, M.D.; Robert, S.; Fournier, R.A. Response of sugar maple (Acer saccharum, Marsh.) tree crown structure to competition in pure versus mixed stands. For. Ecol. Manag. 2016, 374, 20–32. [Google Scholar] [CrossRef]
- Rottmann, M. Wind—Und Sturmschäden im Wald; Verlag: Bad Orb, Germany, 1986. [Google Scholar]
- Zeide, B. Tolerance and self-tolerance of trees. For. Ecol. Mngt. 1985, 13, 149–166. [Google Scholar] [CrossRef]
- Pretzsch, H. Canopy space filling and tree crown morphology in mixed-species stands compared with monocultures. For. Ecol. Manag. 2014, 327, 251–264. [Google Scholar] [CrossRef]
- Morin, X.; Fahse, L.; Scherer-Lorenzen, M.; Bugmann, H. Tree species richness promotes productivity in temperate forests through strong complementarity between species. Ecol. Lett. 2011, 14, 1211–1219. [Google Scholar] [CrossRef]
- Pretzsch, H. Re-Evaluation of Allometry: State-of-the-Art and perspective regarding individuals and stands of woody plants. In Progress in Botany 71; Lüttge, U., Beyschlag, W., Büdel, B., Francis, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 339–369, 400. [Google Scholar]
- West, G.B.; Brown, J.H.; Enquist, B.J. A general model for the origin of allometric scaling laws in biology. Science 1997, 276, 122–126. [Google Scholar] [CrossRef] [PubMed]
- West, G.B.; Enquist, B.J.; Brown, J.H. A general quantitative theory of forest structure and dynamics. Proc. Natl. Acad. Sci. USA 2009, 106, 7040–7045. [Google Scholar] [CrossRef] [PubMed]
- Dahlhausen, J.; Biber, P.; Rötzer, T.; Uhl, E.; Pretzsch, H. Tree Species and Their Space Requirements in Six Urban Environments Worldwide. Forests 2016, 7111, 111. [Google Scholar] [CrossRef]
- Poorter, L.; Bongers, F.; Sterck, F.J.; Wöll, H. Architecture of 53 rain forest tree species differing in adult stature and shade tolerance. Ecology 2003, 84, 602–608. [Google Scholar] [CrossRef]
- Pretzsch, H.; del Río, M.; Ammer Ch Avdagic, A.; Barbeito, I.; Bielak, K.; Brazaitis, G.; Coll, L.; Dirnberger, G.; Drössler, L.; Fabrika, M.; et al. Growth and yield of mixed versus pure stands of Scots pine (Pinus sylvestris L.) and European beech (Fagus sylvatica L.) analysed along a productivity gradient through Europe. Eur. J. For. Res 2015, 134, 927–947. [Google Scholar] [CrossRef]
- Pretzsch, H.; Biber, P.; Uhl, E.; Dahlhausen, J.; Rötzer, T.; Caldentey, J.; Koike, T.; van Con, T.; Chavanne, A.; Seifert, T.; et al. Crown size and growing space requirement of common tree species in urban centres, parks and forests. Urban For. Urban Green. 2015, 14, 466–479. [Google Scholar] [CrossRef]
- Pretzsch, H.; (Centre of Life and Food Sciences Weihenstephan, Technical University of Munich, Hans-Carl-von-Carlowitz-Platz 2, 85354 Freising, Germany). Weiterentwicklung der Dichte—und Mischungsregulierung forstwirtschaftlich wichtiger Baumarten. Äquivalenz-Koeffizienten und Dichte-Steigerungs-Koeffizienten für generische waldbauliche Behandlungsalgorithmen, Allgemeine Forst- und Jagdzeitung. Unpublished work. 2019. [Google Scholar]
- Heym, M.; Ruiz-Peinado, R.; del Rio, M.; Bielak, K.; Forrester, D.I.; Dirnberger, G.; Barbeito, I.; Brazaitis, G.; Ruskytke, I.; Coll, L.; et al. EuMIXFOR empirical forest mensuration and ring width data from pure and mixed stands of Scots pine (Pinus sylvestris L.) and European beech (Fagus sylvatica L.) through Europe. Ann. For. Sci. 2017, 74, 9. [Google Scholar] [CrossRef]
- Pretzsch, H.; Block, J.; Dieler, J.; Dong, P.H.; Kohnle, U.; Nagel, J.; Spellmann, H.; Zingg, A. Comparison between the productivity of pure and mixed stands of Norway spruce and European beech along an ecological gradient. Ann. For. Sci. 2010, 67, 712. [Google Scholar] [CrossRef]
- Pretzsch, H.; Bielak, K.; Block, J.; Bruchwald, A.; Dieler, J.; Ehrhart, H.P.; Kohnle, U.; Nagel, J.; Spellmann, H.; Zasada, M.; et al. Productivity of mixed versus pure stands of oak (Quercus pretraea (Matt.) Liebl. and Quercus robur L.) and European beech (Fagus sylvatica L.) along an ecological gradient. Eur. J. For. Res. 2013, 132, 263–280. [Google Scholar] [CrossRef]
- Thurm, E.A.; Pretzsch, H. Improved productivity and modified tree morphology of mixed versus pure stands of European beech (Fagus sylvatica) and Douglas-fir (Pseudotsuga menziesii) with increasing precipitation and age. Ann. For. Sci. 2016, 73, 1047–1061. [Google Scholar] [CrossRef]
- Thurm, E.A.; Biber, P.; Pretzsch, H. Stem growth is favored at expenses of root growth in mixed stands and humid conditions for Douglas-fir (Pseudotsuga menziesii) and European beech (Fagus sylvatica). Trees 2017, 31, 349–365. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. Linear and Nonlinear Mixed Effects Models. R Package Version 3.1–131. 2017. Available online: https://CRAN.R-project.org/package=nlme> (accessed on 23 July 2019).
- Reineke, L.H. Perfecting a stand-density index for even-aged forests. J. Agric. Res. 1933, 46, 627–638. [Google Scholar]
- Antonio, N.; Tomé, M.; Tomé, J.; Soares, P.; Fontes, L. Effect of tree, stand, and site variables on the allometry of Eucalyptus globulus tree biomass. Can. J. For. Res. 2007, 37, 895–906. [Google Scholar] [CrossRef]
- Hasenauer, H.; Monserud, R.A. A crown ratio model for Austrian forests. For. Ecol. Manag. 1996, 84, 49–60. [Google Scholar] [CrossRef]
- Hasenauer, H. Dimensional relationships of open-grown trees in Austria. For. Ecol. Manag. 1997, 96, 197–206. [Google Scholar] [CrossRef]
- Hynynen, J. Predicting tree crown ratio for unthinned and thinned Scots pine stands. Can. J. For. Res. 1995, 25, 57–62. [Google Scholar] [CrossRef]
- Longetaud, F.; Seifert, T.; Leban, J.M.; Pretzsch, H. Analysis of long-term dynamics of crowns of sessile oaks at stand level by means of spatial statistics. For. Ecol. Manag. 2008, 255, 2007–2019. [Google Scholar] [CrossRef]
- Russell, M.B.; Weiskittel, A.R. Maximum and largest crown width equations for 15 tree species in Maine. North. J. Appl. For. 2011, 28, 84–91. [Google Scholar] [CrossRef]
- Biging, G.S.; Dobbertin, M. Evaluation of competition indices in individual tree growth models. For. Sci. 1995, 41, 360–377. [Google Scholar]
- Grote, R. Estimation of crown radii and crown projection area from stem size and tree position. Ann. For. Sci. 2003, 60, 393–402. [Google Scholar] [CrossRef]
- Monserud, R.A.; Marshall, J.D. Allometric crown relations in three northern Idaho conifer species. Can. J. For. Res. 1999, 29, 521–535. [Google Scholar] [CrossRef]
- Gargaglione, V.; Peri, P.L.; Rubio, G. Allometric relations for biomass partitioning of Nothofagus antarctica trees of different crown classes over a site quality gradient. For. Ecol. Manag. 2010, 259, 1118–1126. [Google Scholar] [CrossRef]
- Mäkelä, A.; Valentine, H.T. Crown ratio influences allometric scaling in trees. Ecology 2006, 87, 2967–2972. [Google Scholar] [CrossRef]
- Pretzsch, H.; Dieler, J. Evidence of variant intra-and interspecific scaling of tree crown structure and relevance for allometric theory. Oecologia 2012, 169, 637–649. [Google Scholar] [CrossRef] [PubMed]
- Muller-Landau, H.C.; Condit, R.S.; Chave, J.; Thomas, S.C.; Bohlman, S.A.; Bunyavejchewin, S.; Harms, K.E. Testing metabolic ecology theory for allometric scaling of tree size, growth and mortality in tropical forests. Ecol. Lett. 2006, 9, 575–588. [Google Scholar] [CrossRef] [PubMed]
- Osawa, A.; Allen, R.B. Allometric theory explains self-thinning relationships of mountain beech and red pine. Ecology 1993, 74, 1020–1032. [Google Scholar] [CrossRef]
- Hein, S.; Spiecker, H. Crown and tree allometry of open-grown ash (Fraxinus excelsior L.) and sycamore (Acer pseudoplatanus L.). Agrofor. Syst. 2008, 73, 205–218. [Google Scholar] [CrossRef]
- Archibald, S.; Bond, W.J. Growing tall vs. growing wide: Tree architecture and allometry of Acacia karroo in forest, savanna, and arid environments. Oikos 2003, 102, 3–14. [Google Scholar] [CrossRef]
- Van Gelder, H.A.; Poorter, L.; Sterck, F.J. Wood mechanics, allometry, and life-history variation in a tropical rain forest tree community. N. Phytol. 2006, 171, 367–378. [Google Scholar] [CrossRef]
- White, P.S. Corner’s rules in eastern deciduous trees: Allometry and its implications for the adaptive architecture of trees. Bull. Torrey Bot. Club 1983, 1, 203–212. [Google Scholar] [CrossRef]
- Bayer, D.; Reischl, A.; Rötzer, T.; Pretzsch, H. Structural response of black locust (Robinia pseudoacacia L.) and small-leaved lime (Tilia cordata Mill.) to varying urban environments analyzed by terrestrial laser scanning: Implications for ecological functions and services. Urban For. Urban Green. 2018, 35, 129–138. [Google Scholar] [CrossRef]
- Jucker, T.; Bouriaud, O.; Coomes, D.A. Crown plasticity enables trees to optimize canopy packing in mixed-species forests. Funct. Ecol. 2015, 29, 1078–1086. [Google Scholar] [CrossRef]
- Verma, N.K.; Lamb, D.W.; Reid, N.; Wilson, B. An allometric model for estimating DBH of isolated and clustered Eucalyptus trees from measurements of crown projection area. For. Ecol. Manag. 2014, 326, 125–132. [Google Scholar] [CrossRef]
- Forrester, D.I.; Benneter, A.; Bouriaud, O.; Bauhus, J. Diversity and competition influence tree allometric relationships–developing functions for mixed-species forests. J. Ecol. 2017, 105, 761–774. [Google Scholar] [CrossRef]
- Goodman, R.C.; Phillips, O.L.; Baker, T.R. The importance of crown dimensions to improve tropical tree biomass estimates. Ecol. Appl. 2014, 24, 680–698. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, E.; Vega, J.A.; Fernández-Alonso, J.M.; Vega-Nieva, D.; Álvarez-González, J.G.; Ruiz-González, A.D. Allometric equations for estimating canopy fuel load and distribution of pole-size maritime pine trees in five Iberian provenances. Can. J. For. Res. 2012, 43, 149–158. [Google Scholar] [CrossRef]
- Mitsopoulos, I.D.; Dimitrakopoulos, A.P. Allometric equations for crown fuel biomass of Aleppo pine (Pinus halepensis Mill.) in Greece. Int. J. Wildland Fire 2007, 16, 642–647. [Google Scholar] [CrossRef]
- Aiba, S.I.; Kohyama, T. Crown architecture and life-history traits of 14 tree species in a warm-temperate rain forest: Significance of spatial heterogeneity. J. Ecol. 1997, 85, 611–624. [Google Scholar] [CrossRef]
- Condés, S.; Sterba, H. Derivation of compatible crown width equations for some important tree species of Spain. For. Ecol. Manag. 2005, 217, 203–218. [Google Scholar] [CrossRef]
- Putz, F.E.; Parker, G.G.; Archibald, R.M. Mechanical abrasion and intercrown spacing. Am. Midl. Nat. 1984, 112, 24–28. [Google Scholar] [CrossRef]
- Fish, H.; Lieffers, V.J.; Silins, U.; Hall, R.J. Crown shyness in lodgepole pine stands of varying stand height, density, and site index in the upper foothills of Alberta. Can. J. For. Res. 2006, 36, 2104–2111. [Google Scholar] [CrossRef]
- Ellenberg, H.; Leuschner, C. Vegetation Mitteleuropas mit den Alpen: In ökologischer, dynamischer und historischer Sicht; Utb: Stuttgart, Germany, 2010. [Google Scholar]
- Lyr, H.; Polster, H.; Fiedler, H.J. Gehölzphysiologie; VEB Gustav Fischer Verlag: Jena, Germany, 1967; 337p. [Google Scholar]
- Mitscherlich, G. Wald, Wachstum und Umwelt. 2. Band, Waldklima und Wasserhaushalt; JD Sauerländer’s Verlag: Frankfurt am Main, Germany, 1971; 365p. [Google Scholar]
- Dieler, J.; Pretzsch, H. Morphological plasticity of European beech (Fagus sylvatica L.) in pure and mixed-species stands. For. Ecol. Manag. 2013, 95, 97–108. [Google Scholar] [CrossRef]
- Juchheim, J.; Annighöfer, P.; Ammer, C.; Calders, K.; Raumonen, P.; Seidel, D. How management intensity and neighborhood composition affect the structure of beech (Fagus sylvatica L.) trees. Trees 2017, 31, 1723–1735. [Google Scholar] [CrossRef]
- Hajek, P.; Seidel, D.; Leuschner, C. Mechanical abrasion, and not competition for light, is the dominant canopy interaction in a temperate mixed forest. For. Ecol. Manag. 2015, 348, 108–116. [Google Scholar] [CrossRef]
- Bravo-Oviedo, A.; Condés, S.; del Rio, M.; Pretzsch, H.; Ducey, M.J. Maximum stand density strongly depends on species-specific wood stability, shade and drought tolerance. Forestry 2018, 91, 459–469. [Google Scholar] [CrossRef]
- Knigge, W.; Schulz, H. Grundriss der Forstbenutzung; Verlag Paul Parey: Hamburg/Berlin, Germany, 1966; 584p. [Google Scholar]
- Trendelenburg, R.; Mayer-Wegelin, H. Das Holz als Rohstoff; Hanser Verlag: München, Germany, 1955; 541p. [Google Scholar]
- Larcher, W. Ökophysiologie der Pflanzen, 5th ed.; Verlag Eugen Ulmer: Stuttgart, Germany, 1994; 394p. [Google Scholar]
- Canham, C.D. Growth and canopy architecture of shade-tolerant trees: Response to canopy gaps. Ecology 1988, 69, 786–795. [Google Scholar] [CrossRef]
- Delagrange, S.; Messier, C.; Lechowicz, M.J.; Dizengremel, P. Physiological, morphological and allocational plasticity in understory deciduous trees: Importance of plant size and light availability. Tree Physiol. 2004, 24, 775–784. [Google Scholar] [CrossRef]
- McCarthy, M.C.; Enquist, B.J. Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocation. Funct. Ecol. 2007, 21, 713–720. [Google Scholar] [CrossRef]
- Gedroc, J.J.; McConnaughay, K.D.M.; Coleman, J.S. Plasticity in root/shoot partitioning: Optimal, ontogenetic, or both? Funct. Ecol. 1996, 10, 44–50. [Google Scholar] [CrossRef]
- Forrester, D.I.; Tachauer, I.H.H.; Annighoefer, P.; Barbeito, I.; Pretzsch, H.; Ruiz-Peinado, R.; Saha, S. Generalized biomass and leaf area allometric equations for European tree species incorporating stand structure, tree age and climate. For. Ecol. Manag. 2017, 396, 160–175, Frankfurt a. Main. 128 p. [Google Scholar] [CrossRef]
- Forrester, D.I.; Ammer, C.; Annighöfer, P.J.; Barbeito, I.; Bielak, K.; Bravo-Oviedo, A.; Hurt, V. Effects of crown architecture and stand structure on light absorption in mixed and monospecific Fagus sylvatica and Pinus sylvestris forests along a productivity and climate gradient through. Eur. J. Ecol. 2018, 106, 746–760. [Google Scholar] [CrossRef]
- Magin, R. Struktur und Leistung mehrschichtiger Mischwälder in den bayerischen Alpen. Mitt Staatsforstverwalt. Bayerns 1959, 30, 161p. [Google Scholar]
- Preuhsler, T. Die Entwicklung von Oberstand und Naturverjüngung in Bergmischwald-Verjüngungsbeständen des Forstamtes Kreuth. Centralbl Für Das Ges Forstwes 1989, 106, 23–54. [Google Scholar]
- Osunkoya, O.O.; Omar-Ali, K.; Amit, N.; Dayan, J.; Daud, D.S.; Sheng, T.K. Comparative height–crown allometry and mechanical design in 22 tree species of Kuala Belalong rainforest, Brunei, Borneo. Am. J. Bot. 2007, 94, 1951–1962. [Google Scholar] [CrossRef]
- Sterba, H.; Dirnberger, G.; Ritter, T. Vertical Distribution of Leaf Area of European Larch (Larix decidua Mill.) and Norway Spruce (Picea abies (L.) Karst.) in Pure and Mixed Stands. Forests 2019, 10, 570. [Google Scholar] [CrossRef]
- Metz, J.; Seidel, D.; Schall, P.; Scheffer, D.; Schulze, E.D.; Ammer, C. Crown modeling by terrestrial laser scanning as an approach to assess the effect of aboveground intra-and interspecific competition on tree growth. For. Ecol. Manag. 2013, 310, 275–288. [Google Scholar] [CrossRef]
- Silva Pedro, M.; Rammer, W.; Seidl, R. Disentangling the effects of compositional and structural diversity on forest productivity. J. Veg. Sci. 2017, 28, 649–658. [Google Scholar] [CrossRef]
- Zeller, L.; Pretzsch, H. Effect of forest structure on stand productivity in Central European forests depends on developmental stage and tree species diversity. For. Ecol. Manag. 2019, 434, 193–204. [Google Scholar] [CrossRef]
- Sterba, H. Assmanns Theorie der Grundflächenhaltung und die “Competition-Density-Rule” der Japaner Kira, Ando und Tadaki. Cbl für das ges Forstwes. 1975, 92, 46–62. [Google Scholar]
- Sterba, H. Natürlicher Bestockungsgrad und Reinekes SDI. Cbl für das ges Forstwes 1981, 98, 101–116. [Google Scholar]
- Pretzsch, H.; Steckel, M.; Heym, M.; Biber, P.; Ammer, C.; Ehbrecht, M.; Bielak, K.; Bravo, F.; Ordóñez, C.; Collet, C.; et al. Stand growth and structure of mixed-species and monospecific stands of Scots pine (Pinus sylvestris L.) and oak (Q. robur L., Quercus petraea (MATT.) LIEBL.) analysed along a productivity gradient through Europe. Eur. J. For. Res. 2019. under review. [Google Scholar]
- Pretzsch, H.; del Río, M. Density regulation of mixed and mono-specific forest stands as a continuum. A concept based on species-specific coefficients of density equivalence and density modification. Forestry 2019. under review. [Google Scholar]
- Sterba, H.; Monserud, R.A. The maximum density concept applied to uneven-aged mixed-species stands. For. Sci. 1993, 39, 432–452. [Google Scholar]
- Fatichi, S.; Leuzinger, S.; Körner, C. Moving beyond photosynthesis: From carbon source to sink-driven vegetation modelling. N. Phytol. 2014, 201, 1086–1095. [Google Scholar] [CrossRef]
- Jucker, T.; Caspersen, J.; Chave, J.; Antin, C.; Barbier, N.; Bongers, F.; Higgins, S.I. Allometric equations for integrating remote sensing imagery into forest monitoring programmes. Glob. Chang. Biol. 2017, 23, 177–190. [Google Scholar] [CrossRef]
- Grams, T.E.; Lüttge, U. Space as a resource. In Progress in Botany 72; Springer: Berlin/Heidelberg, Germany, 2010; pp. 349–370. [Google Scholar]
- Meng, S.X.; Rudnicki, M.; Lieffers, V.J.; Reid, D.E.; Silins, U. Preventing crown collisions increases the crown cover and leaf area of maturing lodgepole pine. J. Ecol. 2006, 94, 681–686. [Google Scholar] [CrossRef]
- Kelty, M.J. Comparative productivity of monocultures and mixed-species stands. In The Ecology and Silviculture of Mixed-Species Forests; Springer: Dordrecht, The Netherlands, 1992; pp. 125–141. [Google Scholar]
- Richards, A.E.; Forrester, D.I.; Bauhus, J.; Scherer-Lorenzen, M. The influence of mixed tree plantations on the nutrition of individual species: A review. Tree Physiol. 2010, 30, 1192–1208. [Google Scholar] [CrossRef]
- Bauhus, J.; Khanna, P.K.; Menden, N. Aboveground and belowground interactions in mixed plantations of Eucalyptus globulus and Acacia mearnsii. Can. J. For. Res. 2000, 30, 1886–1894. [Google Scholar] [CrossRef]
- Schmid, I.; Kazda, M. Vertical distribution and radial growth of coarse roots in pure and mixed stands of Fagus sylvatica and Picea abies. Can. J. For. Res. 2001, 31, 539–548. [Google Scholar] [CrossRef]
- Gale, M.R.; Grigal, D.F. Vertical root distributions of northern tree species in relation to successional status. Can. J. For. Res. 1987, 17, 829–834. [Google Scholar] [CrossRef]
- Stone, E.L.; Kalisz, P.J. On the maximum extent of tree roots. For. Ecol. Manag. 1991, 46, 59–102. [Google Scholar] [CrossRef]
- Williams, L.J.; Paquette, A.; Cavender-Bares, J.; Messier, C.; Reich, P.B. Spatial complementarity in tree crowns explains overyielding in species mixtures. Nat. Ecol. Evol. 2017, 1, 0063. [Google Scholar] [CrossRef]
- Pretzsch, H.; Biber, P. Tree species mixing can increase maximum stand density. Can. J. For. Res. 2016, 46, 1179–1193. [Google Scholar] [CrossRef]
- Pretzsch, H. Ertragstafel-Korrekturfaktoren für Umwelt- und Mischungseffekte. Available online: https://www.semanticscholar.org/paper/Ertragstafel-Korrekturfaktoren-f%C3%BCr-Umwelt-und-Pretzsch/93cead166218f3c96f2ed502a064d234e0d800cc (accessed on 23 July 2019).
- Drössler, L.; Agestam, E.; Bielak, K.; Dudzinska, M.; Koricheva, J.; Liziniewicz, M.; Löf, M.; Mason, B.; Pretzsch, H.; Valkonen, S.; et al. Over- and Underyielding in Time and Space in Experiments with Mixed Stands of Scots Pine and Norway Spruce. Forests 2018, 9, 495. [Google Scholar] [CrossRef]
- Liang, J.; Crowther, T.W.; Picard, N.; Wiser, S.; Zhou, M.; Alberti, G.; Schulze, E.D.; McGuire, A.D.; Bozzato, F.; Pretzsch, H.; et al. Positive biodiversity-productivity relationship predominant in global forests. Science 2016, 354, 1–12. [Google Scholar] [CrossRef]
- Bravo-Oviedo, A.; Pretzsch, H.; del Río, M. Dynamics, Silviculture and Management of Mixed Forests, Managing Forest Ecosystems 31; Springer Nature: Cham, Germany, 2018; 420p. [Google Scholar]
- Pretzsch, H. Growth and Structure in Mixed-Species stands compared with monocultures: Review and perspectives. In Dynamics, Silviculture and Management of Mixed Forests, Managing Forest Ecosystems 31; Bravo-Oviedo, A., Pretzsch, H., del Río, M., Eds.; Springer Nature: Cham, Germany, 2018; pp. 131–183. [Google Scholar]
- Cole, W.G.; Lorimer, C.G. Predicting tree growth from crown variables in managed northern hardwood stands. For. Ecol. Manag. 1994, 67, 159–175. [Google Scholar] [CrossRef]
- Maguire, D.A.; Brissette, J.C.; Gu, L. Crown structure and growth efficiency of red spruce in uneven-aged, mixed-species stands in Maine. Can. J. For. Res. 1998, 28, 1233–1240. [Google Scholar] [CrossRef]
- Pretzsch, H.; Biber, P.; Ďurský, J. The single tree-based stand simulator SILVA: Construction. application and evaluation. For. Ecol. Manag. 2002, 162, 3–21. [Google Scholar] [CrossRef]
- Thorpe, H.C.; Astrup, R.; Trowbridge, A.; Coates, K.D. Competition and tree crowns: A neighborhood analysis of three boreal tree species. For. Ecol. Manag. 2010, 259, 1586–1596. [Google Scholar] [CrossRef]
- Bravo, F.; Fabrika, M.; Ammer, C.; Barreiro, S.; Bielak, K.; Coll, L.; Pach, M. Modelling approaches for mixed forests dynamics prognosis. Research gaps and opportunities. For. Syst. 2019, 28, eR002. [Google Scholar] [CrossRef]
- Pretzsch, H.; Zenner, E.K. Toward managing mixed-species stands: From parametrization to prescription. For. Ecosyst. 2017, 4, 19. [Google Scholar] [CrossRef]
- Pretzsch, H. Grundlagen der Waldwachstumsforschung; Springer: Berlin, Germany, 2019; 664p. [Google Scholar]
- del Río, M.; Bravo-Oviedo, A.; Ruiz-Peinado, R.; Condés, S. Tree allometry variation in response to intra-and inter-specific competitions. Trees 2019, 33, 121–138. [Google Scholar] [CrossRef]
- Biber, P.; Borges, J.; Moshammer, R.; Barreiro, S.; Botequim, B.; Brodrechtova, Y.; Eriksson, L. How sensitive are ecosystem services in European forest landscapes to silvicultural treatment? Forests 2015, 6, 1666–1695. [Google Scholar] [CrossRef]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Species | Variables | n | a0 | se (a0) | p-Value | a1 | se (a1) | p-Value | a2 | se (a2) | p-Value | a3 | se (a3) | p-Value |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | N. sp | cpa~h | 11784 | 1.210 | 0.100 | 0.000 | 0.832 | 0.033 | 0.000 | ||||||
| 1 | S. pi | cpa~h | 2734 | −0.992 | 0.850 | 0.243 | 1.489 | 0.280 | 0.000 | ||||||
| 1 | D-fir | cpa~h | 1480 | −0.377 | 0.196 | 0.054 | 1.507 | 0.068 | 0.000 | ||||||
| 1 | E. be | cpa~h | 11802 | 1.458 | 0.531 | 0.006 | 1.081 | 0.170 | 0.000 | ||||||
| 1 | s. oak | cpa~h | 3920 | −1.900 | 0.638 | 0.003 | 2.057 | 0.189 | 0.000 | ||||||
| 2 | N. sp | cpa~d | 4734 | −2.017 | 0.108 | 0.000 | 1.396 | 0.013 | 0.000 | ||||||
| 2 | S. pi | cpa~d | 2734 | −3.691 | 0.091 | 0.000 | 1.963 | 0.029 | 0.000 | ||||||
| 2 | D-fir | cpa~d | 1480 | −1.606 | 0.140 | 0.000 | 1.341 | 0.218 | 0.000 | ||||||
| 2 | E. be | cpa~d | 2830 | −0.434 | 0.080 | 0.000 | 1.160 | 0.014 | 0.000 | ||||||
| 2 | s. oak | cpa~d | 692 | −4.408 | 0.322 | 0.000 | 2.204 | 0.075 | 0.000 | ||||||
| 3 | N. sp | hcb~h | 11783 | −1.890 | 0.054 | 0.000 | 1.354 | 0.016 | 0.000 | ||||||
| 3 | S. pi | hcb~h | 4483 | −0.927 | 0.037 | 0.000 | 1.155 | 0.012 | 0.000 | ||||||
| 3 | D-fir | hcb~h | 1689 | −2.836 | 0.129 | 0.000 | 1.582 | 0.038 | 0.000 | ||||||
| 3 | E. be | hcb~h | 11802 | −2.347 | 0.046 | 0.000 | 1.422 | 0.014 | 0.000 | ||||||
| 3 | s. oak | hcb~h | 3919 | −0.953 | 0.035 | 0.000 | 1.109 | 0.011 | 0.000 | ||||||
| 4 | various species | cpa~R | 14 | −17.87 | 38.65 | 0.652 | 177.36 | 67.31 | 0.022 | ||||||
| 4 | various species | cr~R | 14 | 2.090 | 1.097 | 0.081 | 5.243 | 1.911 | 0.018 | ||||||
| 6 | S. pi mono-mix | cpa~d | 3801 | −2.937 | 0.168 | 0.000 | 1.693 | 0.046 | 0.000 | −0.717 | 0.235 | 0.006 | 0.207 | 0.067 | 0.002 |
| 6 | s. oak mono-mix | cpa~d | 3098 | −1.899 | 0.144 | 0.000 | 1.515 | 0.041 | 0.000 | 1.289 | 0.184 | 0.000 | −0.303 | 0.055 | 0.000 |
| 6 | S. pi mono-mix | cpa~d | 1382 | −3.340 | 0.184 | 0.000 | 1.795 | 0.050 | 0.000 | −0.152 | 0.069 | 0.039 | |||
| 6 | E. be mono-mix | cpa~d | 1818 | −0.323 | 0.136 | 0.017 | 1.127 | 0.032 | 0.000 | 0.245 | 0.055 | 0.000 | |||
| 7 | S. pi mono-mix | hcb~h | 3152 | −0.665 | 0.089 | 0.000 | 1.081 | 0.025 | 0.000 | 0.758 | 0.124 | 0.000 | −0.261 | 0.038 | 0.000 |
| 7 | s. oak mono-mix | hcb~h | 2486 | −0.981 | 0.144 | 0.000 | 1.097 | 0.043 | 0.000 | 0.486 | 0.162 | 0.007 | −0.195 | 0.052 | 0.000 |
| 7 | S. pi mono-mix | hcb~h | 1194 | 0.227 | 0.105 | 0.032 | 0.795 | 0.031 | 0.000 | 0.053 | 0.022 | 0.024 | |||
| 7 | E. be mono-mix | hcb~h | 1335 | −3.038 | 0.296 | 0.000 | 1.670 | 0.093 | 0.000 | 1.604 | 0.380 | 0.000 | −0.526 | 0.120 | 0.000 |
| 8 | various species | SDI~cpa | 14 | 8637.4 | 2622.5 | 0.006 | 8637.4 | 2622.5 | 0.006 | ||||||
| 9 | various species | N~cpa | 1910 | 10.212 | 0.055 | 0.000 | −1.294 | 0.018 | 0.000 | −0.351 | 0.106 | 0.000 | 0.196 | 0.034 | 0.000 |
| 10 | N.sp (N.sp, E. be) | N~dq | 178 | 12.814 | 0.163 | 0.000 | −1.841 | 0.048 | 0.000 | ||||||
| 10 | E.be (N.sp, E. be) | N~dq | 178 | 11.410 | 0.101 | 0.000 | −1.533 | 0.033 | 0.000 | ||||||
| 10 | S.pi (S.pi, E. be) | N~dq | 32 | 13.594 | 0.527 | 0.000 | −2.143 | 0.161 | 0.000 | ||||||
| 10 | E.be(S.pi, E. be) | N~dq | 32 | 12.494 | 0.454 | 0.000 | −1.856 | 0.143 | 0.000 | ||||||
| 10 | D-fir (D-fir, E. be) | N~dq | 18 | 11.062 | 0.425 | 0.000 | −1.326 | 0.116 | 0.000 | ||||||
| 10 | E.be (D-fir, E. be) | N~dq | 18 | 11.831 | 0.539 | 0.000 | −1.672 | 0.170 | 0.000 | ||||||
| 10 | s.oak (s.oak, E. be) | N~dq | 254 | 11.869 | 0.207 | 0.000 | −1.820 | 0.062 | 0.000 | ||||||
| 10 | E.be (s.oak, E. be) | N~dq | 254 | 11.979 | 0.178 | 0.000 | −1.823 | 0.052 | 0.000 | ||||||
| 10 | S.pi (S.pi, s. oak) | N~dq | 36 | 12.599 | 0.730 | 0.000 | −1.808 | 0.219 | 0.000 | ||||||
| 10 | s.oak (S.pi, s. oak) | N~dq | 36 | 13.114 | 0.473 | 0.000 | −2.033 | 0.146 | 0.000 | ||||||
| 10 | N.sp & E. be, exp. | N~dq | 178 | 12.081 | 0.129 | 0.000 | −1.685 | 0.040 | 0.000 | ||||||
| 10 | S.pi & E. be, exp. | N~dq | 32 | 12.568 | 0.554 | 0.000 | −1.841 | 0.173 | 0.000 | ||||||
| 10 | D-fir & E. be, exp. | N~dq | 18 | 11.607 | 0.666 | 0.000 | −1.530 | 0.203 | 0.000 | ||||||
| 10 | s.oak & E.be, exp. | N~dq | 254 | 10.661 | 0.148 | 0.000 | −1.428 | 0.049 | 0.000 | ||||||
| 10 | S.pi & s. oak, exp. | N~dq | 36 | 12.817 | 0.821 | 0.000 | −1.884 | 0.247 | 0.000 | ||||||
| 10 | N.sp & E. be, obs. | N~dq | 178 | 11.316 | 0.129 | 0.000 | −1.505 | 0.043 | 0.000 | ||||||
| 10 | S.pi & E. be, obs. | N~dq | 32 | 12.055 | 0.557 | 0.000 | −1.691 | 0.181 | 0.000 | ||||||
| 10 | D-fir & E. be, obs. | N~dq | 18 | 11.017 | 0.669 | 0.000 | −1.384 | 0.222 | 0.000 | ||||||
| 10 | s.oak & E.be, obs. | N~dq | 254 | 11.497 | 0.176 | 0.000 | −1.648 | 0.061 | 0.000 | ||||||
| 10 | S.pi & s. oak, obs | N~dq | 36 | 13.360 | 0.572 | 0.000 | −2.073 | 0.179 | 0.000 |
| Species Combination | N. spruce E. beech | s. oak E. beech | S. pine E. beech | Douglas-fir E. beech | S. pine s. oak |
|---|---|---|---|---|---|
| n | 178 | 254 | 32 | 18 | 36 |
| mean | 1.020 | 1.257 | 1.130 | 1.281 | 1.137 |
| se | 0.015 | 0.046 | 0.054 | 0.141 | 0.040 |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pretzsch, H. The Effect of Tree Crown Allometry on Community Dynamics in Mixed-Species Stands versus Monocultures. A Review and Perspectives for Modeling and Silvicultural Regulation. Forests 2019, 10, 810. https://doi.org/10.3390/f10090810
Pretzsch H. The Effect of Tree Crown Allometry on Community Dynamics in Mixed-Species Stands versus Monocultures. A Review and Perspectives for Modeling and Silvicultural Regulation. Forests. 2019; 10(9):810. https://doi.org/10.3390/f10090810
Chicago/Turabian StylePretzsch, Hans. 2019. "The Effect of Tree Crown Allometry on Community Dynamics in Mixed-Species Stands versus Monocultures. A Review and Perspectives for Modeling and Silvicultural Regulation" Forests 10, no. 9: 810. https://doi.org/10.3390/f10090810
APA StylePretzsch, H. (2019). The Effect of Tree Crown Allometry on Community Dynamics in Mixed-Species Stands versus Monocultures. A Review and Perspectives for Modeling and Silvicultural Regulation. Forests, 10(9), 810. https://doi.org/10.3390/f10090810