Responses of Soil and Microbial C:N:P Stoichiometry to Vegetation Succession in a Karst Region of Southwest China



Abstract
1. Introduction
2. Materials and Methods
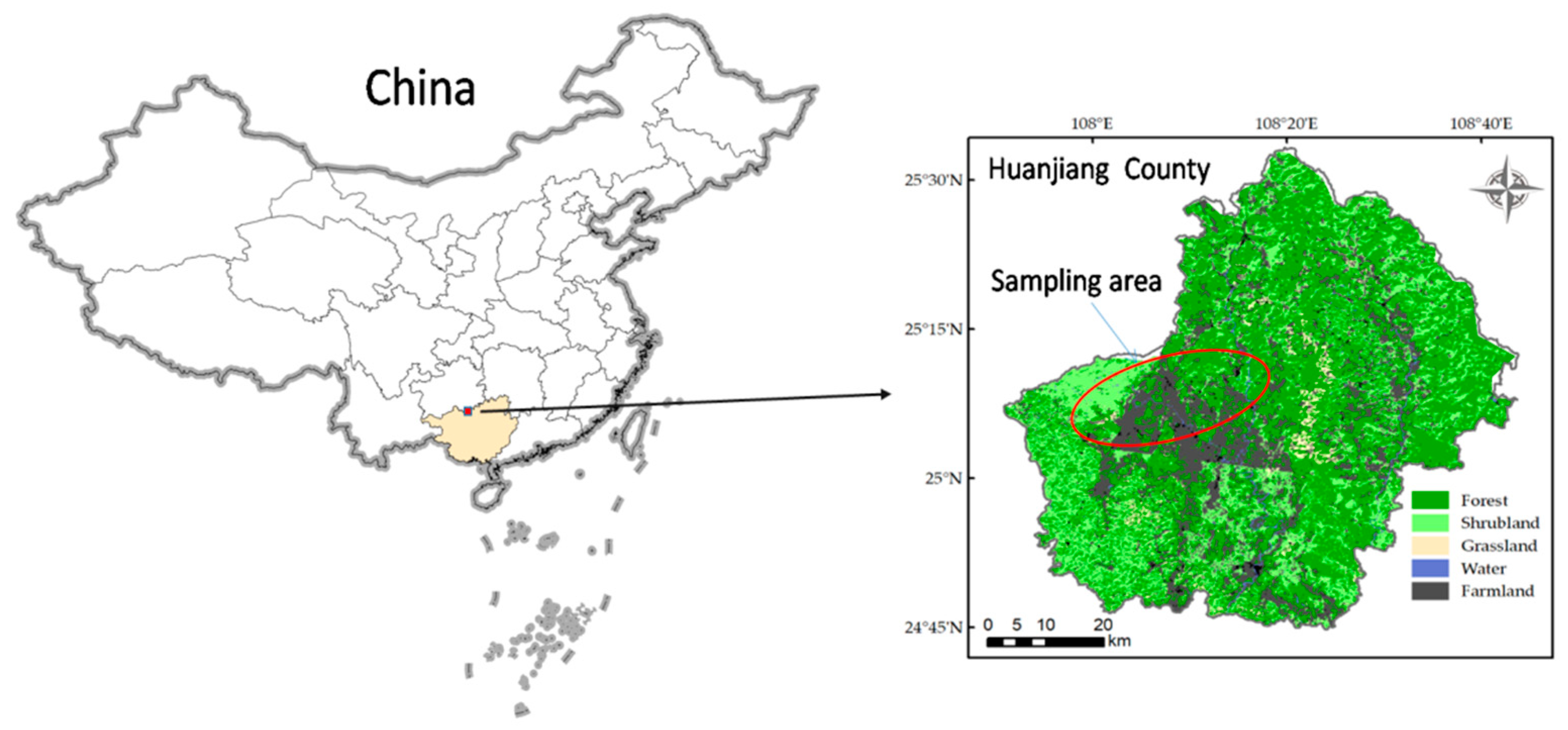
2.1. Study Region
2.2. Soil Sampling
2.3. Chemical Analysis
2.4. Data Analysis
3. Results
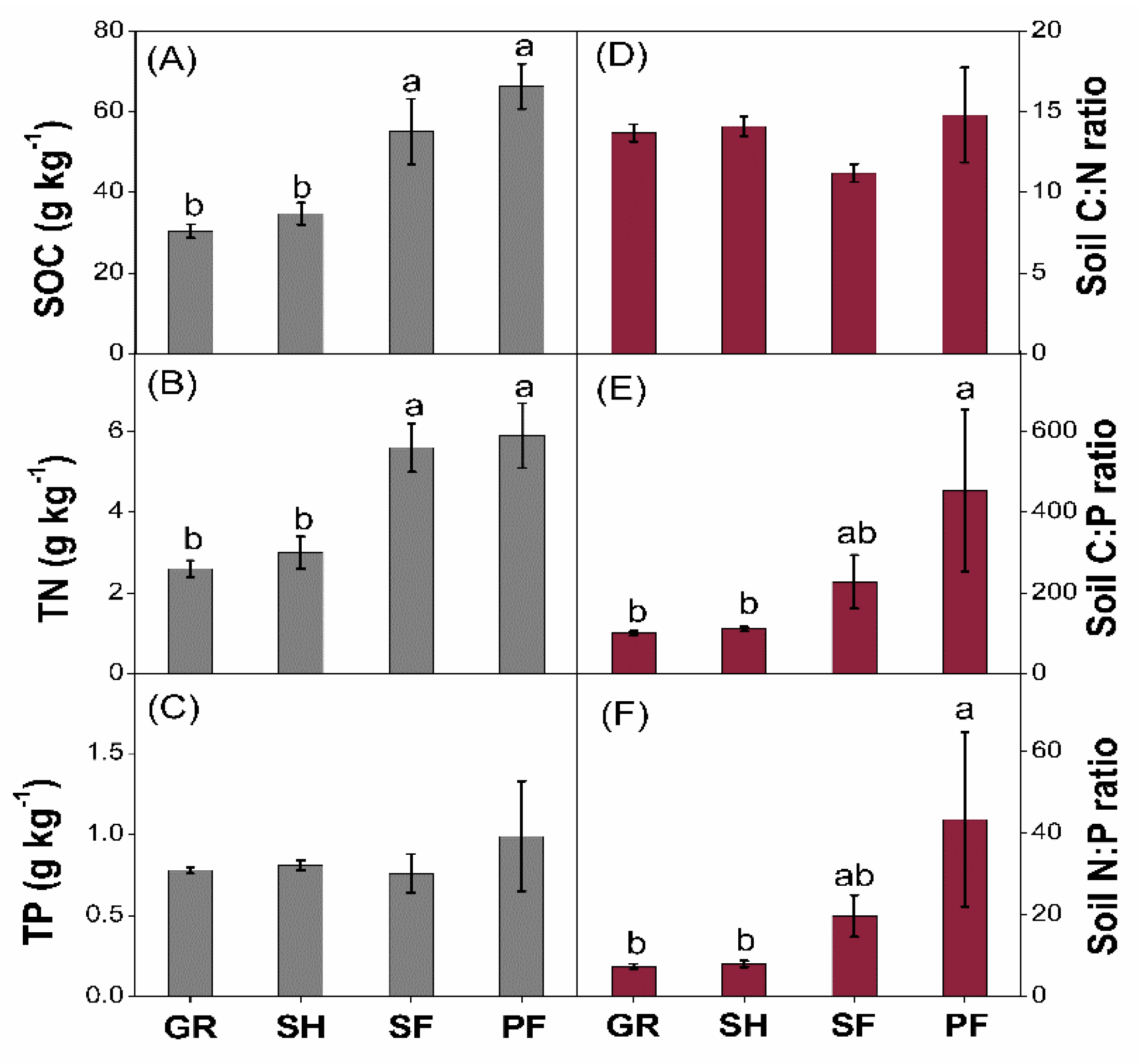
3.1. Soil C:N:P Stoichiometry
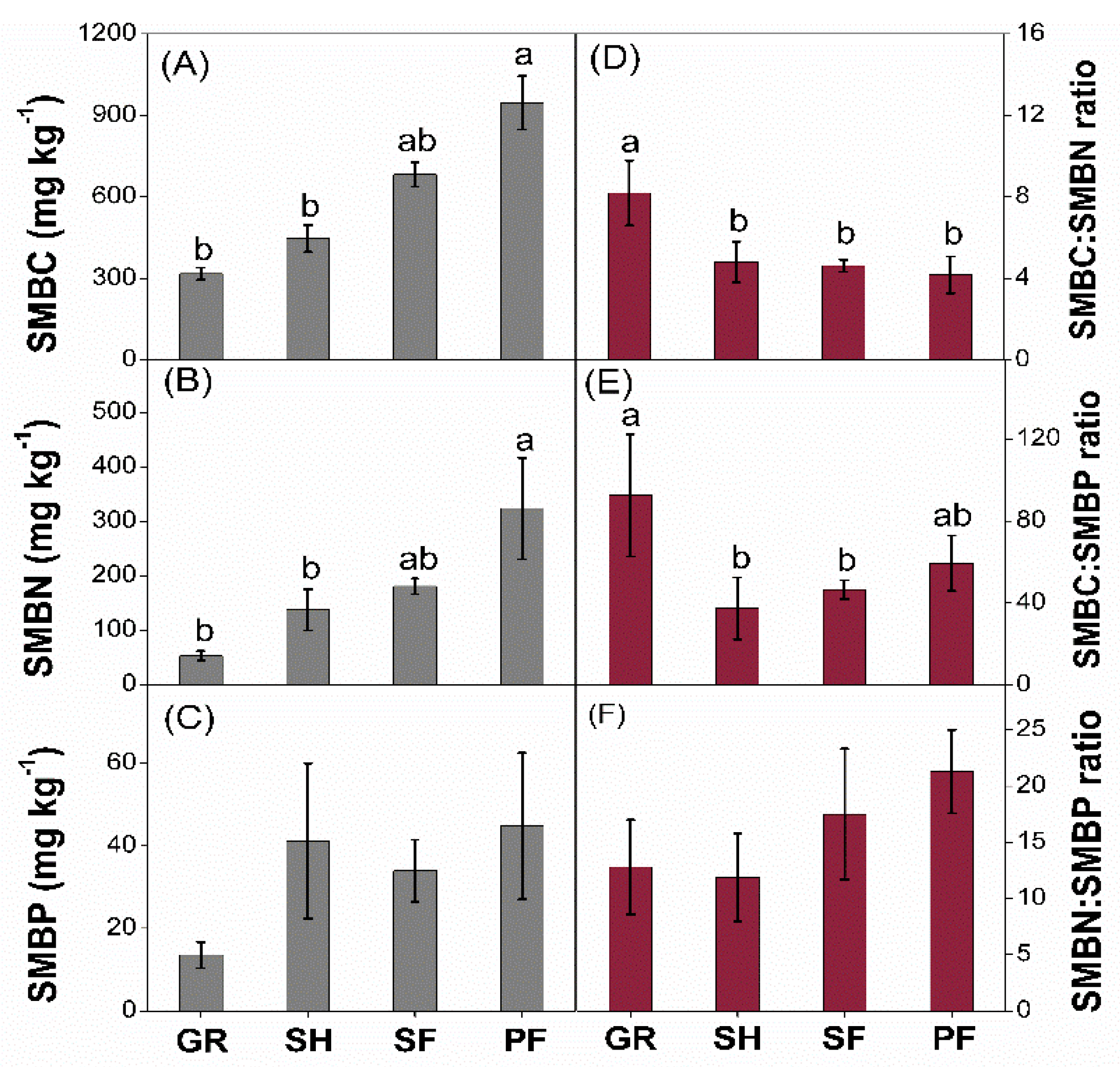
3.2. Microbial C:N:P Stoichiometry
3.3. Relationships between Soil and Microbial C:N:P Stoichiometry
4. Discussion
4.1. Vegetation Succession Effects on Soil C:N:P Stoichiometry
4.2. Vegetation Succession Effects on Microbial C:N:P Stoichiometry
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Redfield, A.C. The biological control of chemical factors in the environment. Am. Sci. 1958, 46, 205–221. [Google Scholar]
- Cleveland, C.C.; Liptzin, D. C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass? Biogeochemistry 2007, 85, 235–252. [Google Scholar] [CrossRef]
- Tian, H.; Chen, G.; Zhang, C.; Melillo, J.M.; Hall, C.A.S. Pattern and variation of C:N:P ratios in China’s soils: A synthesis of observational data. Biogeochemistry 2010, 98, 139–151. [Google Scholar] [CrossRef]
- Mooshammer, M.; Wanek, W.; Schnecker, J.; Wild, B.; Leitner, S.; Hofhansl, F.; Bloechl, A.; Haemmerle, I.; Frank, A.H.; Fuchslueger, L.; et al. Stoichiometric controls of nitrogen and phosphorus cycling in decomposing beech leaf litter. Ecology 2012, 93, 770–782. [Google Scholar] [CrossRef] [PubMed]
- Penuelas, J.; Sardans, J.; Rivas-Ubach, A.; Janssens, I.A. The human-induced imbalance between C, N and P in Earth’s life system. Glob. Chang. Biol. 2012, 18, 3–6. [Google Scholar] [CrossRef]
- Sardans, J.; Rivas-Ubach, A.; Penuelas, J. The C:N:P stoichiometry of organisms and ecosystems in a changing world: A review and perspectives. Perspect. Plant. Ecol. 2012, 14, 33–47. [Google Scholar] [CrossRef]
- Fan, H.; Wu, J.; Liu, W.; Yuan, Y.; Hu, L.; Cai, Q. Linkages of plant and soil C:N:P stoichiometry and their relationships to forest growth in subtropical plantations. Plant. Soil 2015, 392, 127–138. [Google Scholar] [CrossRef]
- Lambers, H.; Mougel, C.; Jaillard, B.; Hinsinger, P. Plant-microbe-soil interactions in the rhizosphere: An evolutionary perspective. Plant. Soil 2009, 321, 83–115. [Google Scholar] [CrossRef]
- Abbott, L.K.; Tang, C.; Reuter, D. Soil-plant-microbe interactions from microscopy to field practice. Plant. Soil 2011, 348, 1–5. [Google Scholar] [CrossRef][Green Version]
- Bui, E.N.; Henderson, B.L. C:N:P stoichiometry in Australian soils with respect to vegetation and environmental factors. Plant. Soil 2013, 373, 553–568. [Google Scholar] [CrossRef]
- Zechmeister-Boltenstern, S.; Keiblinger, K.M.; Mooshammer, M.; Penuelas, J.; Richter, A.; Sardans, J.; Wanek, W. The application of ecological stoichiometry to plant-microbial-soil organic matter transformations. Ecol. Monogr. 2015, 85, 133–155. [Google Scholar] [CrossRef]
- Zhou, Y.; Boutton, T.W.; Wu, X.B. Soil C:N:P stoichiometry responds to vegetation change from grassland to woodland. Biogeochemistry 2018, 140, 341–357. [Google Scholar] [CrossRef]
- Zhao, F.; Sun, J.; Ren, C.; Kang, D.; Deng, J.; Han, X.; Yang, G.; Feng, Y.; Ren, G. Land use change influences soil C, N, and P stoichiometry under ‘Grain-to-Green Program’ in China. Sci. Rep. 2015, 5, 10195. [Google Scholar]
- Van der Heijden, M.G.A.; Bardgett, R.D.; van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Buchkowski, R.W.; Schmitz, O.J.; Bradford, M.A. Microbial stoichiometry overrides biomass as a regulator of soil carbon and nitrogen cycling. Ecology 2015, 96, 1139–1149. [Google Scholar] [CrossRef] [PubMed]
- Tischer, A.; Potthast, K.; Hamer, U. Land-use and soil depth affect resource and microbial stoichiometry in a tropical mountain rainforest region of southern Ecuador. Oecologia 2014, 175, 375–393. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.L.; Chen, L.Y.; Peng, Y.F.; Ding, J.Z.; Li, F.; Yang, G.B.; Kou, D.; Liu, L.; Fang, K.; Zhang, B.B.; et al. Linking microbial C:N:P stoichiometry to microbial community and abiotic factors along a 3500-km grassland transect on the Tibetan Plateau. Glob. Ecol. Biogeogr. 2016, 25, 1416–1427. [Google Scholar] [CrossRef]
- Fanin, N.; Fromin, N.; Buatois, B.; Haettenschwiler, S. An experimental test of the hypothesis of non-homeostatic consumer stoichiometry in a plant littermicrobe system. Ecol. Lett. 2013, 16, 764–772. [Google Scholar] [CrossRef]
- Wang, S.J.; Liu, Q.M.; Zhang, D.F. Karst rocky desertification in southwestern China: Geomorphology, landuse, impact and rehabilitation. Land Degrad. Dev. 2004, 15, 115–121. [Google Scholar] [CrossRef]
- Yin, R.; Yin, G. China’s Primary Programs of Terrestrial Ecosystem Restoration: Initiation, Implementation, and Challenges. Environ. Manag. 2010, 45, 429–441. [Google Scholar] [CrossRef]
- Shi, S.; Han, P. Estimating the soil carbon sequestration potential of China’s Grain for Green Project. Glob. Biogeochem. Cycles 2014, 28, 1279–1294. [Google Scholar] [CrossRef]
- Lu, F.; Hu, H.; Sun, W.; Zhu, J.; Liu, G.; Zhou, W.; Zhang, Q.; Shi, P.; Liu, X.; Wu, X.; et al. Effects of national ecological restoration projects on carbon sequestration in China from 2001 to 2010. Proc. Natl. Acad. Sci. USA 2018, 115, 4039–4044. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Brandt, M.; Yue, Y.; Horion, S.; Wang, K.; Keersmaecker, W.D.; Feng, T.; Schurgers, G.; Xiao, X.; Luo, Y. Increased vegetation growth and carbon stock in China karst via ecological engineering. Nat. Sustain. 2018, 1, 44–50. [Google Scholar] [CrossRef]
- Zhang, J.T. Succession analysis of plant communities in abandoned croplands in the eastern Loess Plateau of China. J. Arid Environ. 2005, 63, 458–474. [Google Scholar] [CrossRef]
- Wen, L.; Song, T.; Du, H.; Wang, K.; Peng, W.; Zeng, F.; Zeng, Z.; He, T. The succession characteristics and its driving mechanism of plant community in karst region, Southwest China. Acta Ecol. Sin. 2015, 35, 5822–5833. [Google Scholar]
- Niinemets, U.; Kull, K. Co-limitation of plant primary productivity by nitrogen and phosphorus in a species-rich wooded meadow on calcareous soils. Acta Oecol. 2005, 28, 345–356. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, W.; Wang, K.; Pan, F.; Yang, S.; Shu, S. Factors controlling accumulation of soil organic carbon along vegetation succession in a typical karst region in Southwest China. Sci. Total Environ. 2015, 521, 52–58. [Google Scholar] [CrossRef]
- Xiao, K.; He, T.; Chen, H.; Peng, W.; Song, T.; Wang, K.; Li, D. Impacts of vegetation restoration strategies on soil organic carbon and nitrogen dynamics in a karst area, Southwest China. Ecol. Eng. 2017, 101, 247–254. [Google Scholar] [CrossRef]
- Wen, L.; Li, D.; Yang, L.; Luo, P.; Chen, H.; Xiao, K.; Song, T.; Zhang, W.; He, X.; Chen, H.; et al. Rapid recuperation of soil nitrogen following agricultural abandonment in a karst area, southwest China. Biogeochemistry 2016, 129, 341–354. [Google Scholar] [CrossRef]
- Zhu, H.; He, X.; Wang, K.; Su, Y.; Wu, J. Interactions of vegetation succession, soil bio-chemical properties and microbial communities in a Karst ecosystem. Eur. J. Soil Biol. 2012, 51, 1–7. [Google Scholar] [CrossRef]
- Zhao, C.; Long, J.; Liao, H.; Zheng, C.; Li, J.; Liu, L.; Zhang, M. Dynamics of soil microbial communities following vegetation succession in a karst mountain ecosystem, Southwest China. Sci. Rep. 2019, 9, 2160. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Cheng, X.; Dang, H.; Ye, C.; Zhang, Y.; Zhang, Q. Linking litter production, quality and decomposition to vegetation succession following agricultural abandonment. Soil Biol. Biochem. 2013, 57, 803–813. [Google Scholar] [CrossRef]
- Xiao, K.; Li, D.; Wen, L.; Yang, L.; Luo, P.; Chen, H.; Wang, K. Dynamics of soil nitrogen availability during post-agricultural succession in a karst region, southwest China. Geoderma 2018, 314, 184–189. [Google Scholar] [CrossRef]
- Song, M.; He, T.; Chen, H.; Wang, K.; Li, D. Dynamics of soil gross nitrogen transformations during post-agricultural succession in a subtropical karst region. Geoderma 2019, 341, 1–9. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, J.; Pan, F.; Li, D.; Chen, H.; Wang, K. Changes in nitrogen and phosphorus limitation during secondary succession in a karst region in southwest China. Plant. Soil 2015, 391, 77–91. [Google Scholar] [CrossRef]
- Gusewell, S. N:P ratios in terrestrial plants: Variation and functional significance. New Phytol. 2004, 164, 243–266. [Google Scholar] [CrossRef]
- Hu, N.; Li, H.; Tang, Z.; Li, Z.; Li, G.; Jiang, Y.; Hu, X.; Lou, Y. Community size, activity and C:N stoichiometry of soil microorganisms following reforestation in a Karst region. Eur. J. Soil Biol. 2016, 73, 77–83. [Google Scholar] [CrossRef]
- WRB. World Reference Base for Soil Resources; FAO, ISRIC, ISSS: Rome, Italy, 2006. [Google Scholar]
- Lu, R. Methods of Soil Science and Agricultural Chemistry Analysis; China Agriculture Press: Beijing, China, 2000. (In Chinese) [Google Scholar]
- John, M.K. Colorimetric determination of phosphorus in soil and plant materials with ascorbic acid. Soil Sci. 1970, 109, 214–220. [Google Scholar] [CrossRef]
- Brookes, P.C.; Powlson, D.S.; Jenkinson, D.S. Measurement of microbial biomass phosphorus in soil. Soil Biol. Biochem. 1982, 14, 319–329. [Google Scholar] [CrossRef]
- Wu, J.; Joergensen, R.G.; Pommerening, B.; Chaussod, R.; Brookes, P.C. Measurement of soil microbial biomass C by fumigation-extraction-an automated procedure. Soil Biol. Biochem. 1990, 22, 1167–1169. [Google Scholar] [CrossRef]
- Wang, B.; Liu, G.B.; Xue, S.; Zhu, B. Changes in soil physico-chemical and microbiological properties during natural succession on abandoned farmland in the Loess Plateau. Environ. Earth Sci. 2011, 62, 915–925. [Google Scholar] [CrossRef]
- Li, Y.; Wu, J.; Liu, S.; Shen, J.; Huang, D.; Su, Y.; Wei, W.; Syers, J.K. Is the C:N:P stoichiometry in soil and soil microbial biomass related to the landscape and land use in southern subtropical China? Glob. Biogeochem. Cycles 2012, 26, GB4002. [Google Scholar] [CrossRef]
- Du, Y.; Pan, G.; Li, L.; Hu, Z.; Wang, X. Leaf N/P ratio and nutrient reuse between dominant species and stands: Predicting phosphorus deficiencies in Karst ecosystems, southwestern China. Environ. Earth Sci. 2011, 64, 299–309. [Google Scholar] [CrossRef]
- Heuck, C.; Weig, A.; Spohn, M. Soil microbial biomass C:N:P stoichiometry and microbial use of organic phosphorus. Soil Biol. Biochem. 2015, 85, 119–129. [Google Scholar] [CrossRef]
- Ren, C.; Zhao, F.; Kang, D.; Yang, G.; Han, X.; Tong, X.; Feng, Y.; Ren, G. Linkages of C:N:P stoichiometry and bacterial community in soil following afforestation of former farmland. For. Ecol. Manag. 2016, 376, 59–66. [Google Scholar] [CrossRef]
- Strickland, M.S.; Rousk, J. Considering fungal: Bacterial dominance in soils—Methods, controls, and ecosystem implications. Soil Biol. Biochem. 2010, 42, 1385–1395. [Google Scholar] [CrossRef]
- Wilson, W.A.; Roach, P.J.; Montero, M.; Baroja-Fernandez, E.; Jose Munoz, F.; Eydallin, G.; Viale, A.M.; Pozueta-Romero, J. Regulation of glycogen metabolism in yeast and bacteria. FEMS Microbiol. Rev. 2010, 34, 952–985. [Google Scholar] [CrossRef]
- Achbergerova, L.; Nahalka, J. Polyphosphate-an ancient energy source and active metabolic regulator. Microb. Cell Fact. 2011, 10, 63. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Vegetation Type | Canopy Cover (%) | Community Height (m) | Dominant Species | Altitude (m) | Slope (o) |
|---|---|---|---|---|---|
| Grassland | 74.2 ± 5.5 | 1.6 ± 0.5 | Ischaemum indicum, Imperata cylindrical, Murdannia triquetra | 287–307 | 26.3 ± 5.5 |
| Shrubland | 82.5 ± 2.7 | 2.2 ± 0.4 | Vitex negundo, Rhus chinensis | 294–309 | 28.9 ± 6.8 |
| Secondary forest | 77.5 ± 4.1 | 6.8 ± 0.7 | Alangium chinense, Itoa orientalis | 292–356 | 30.7 ± 7.2 |
| Primary forest | 88.4 ± 3.8 | 8.5 ± 0.5 | Biota orientalis, Sinosideroxylon pedunculatum | 376–578 | 34.5 ± 4.5 |
| SOC | TN | TP | SMBC | SMBN | SMBP | Soil C:N | Soil C:P | Soil N:P | SMBC:SMBN | SMBC:SMBP | SMBN:SMBP | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SOC | 1 | 0.78 ** | 0.36 | 0.36 | 0.68 ** | 0.36 | 0.22 | 0.33 | 0.28 | −0.45 * | −0.33 | 0.06 |
| TN | 1 | −0.02 | 0.01 | 0.22 | −0.01 | −0.43 * | 0.55 ** | 0.55 ** | −0.49 * | −0.38 | 0.21 | |
| TP | 1 | 0.77 ** | 0.58 ** | 0.46 * | 0.53 ** | −0.57 ** | −0.58 ** | 0.18 | 0.10 | −0.26 | ||
| SMBC | 1 | 0.65 ** | 0.33 | 0.56 ** | −0.18 | −0.20 | 0.23 | 0.32 | −0.02 | |||
| SMBN | 1 | 0.73 ** | 0.66 ** | 0.01 | −0.03 | −0.36 | −0.16 | 0.07 | ||||
| SMBP | 1 | 0.55 ** | −0.15 | −0.19 | −0.28 | −0.39 | −0.42 * | |||||
| Soil C:N | 1 | −0.31 | −0.39 | 0.13 | 0.21 | −0.19 | ||||||
| Soil C:P | 1 | 0.99 ** | −0.32 | −0.14 | 0.31 | |||||||
| Soil N:P | 1 | −0.30 | −0.13 | 0.54 * | ||||||||
| SMBC:SMBN | 1 | 0.48 * | −0.30 | |||||||||
| SMBC:SMBP | 1 | 0.41 * | ||||||||||
| SMBN:SMBP | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, M.; Peng, W.; Du, H.; Xu, Q. Responses of Soil and Microbial C:N:P Stoichiometry to Vegetation Succession in a Karst Region of Southwest China. Forests 2019, 10, 755. https://doi.org/10.3390/f10090755
Song M, Peng W, Du H, Xu Q. Responses of Soil and Microbial C:N:P Stoichiometry to Vegetation Succession in a Karst Region of Southwest China. Forests. 2019; 10(9):755. https://doi.org/10.3390/f10090755
Chicago/Turabian StyleSong, Min, Wanxia Peng, Hu Du, and Qingguo Xu. 2019. "Responses of Soil and Microbial C:N:P Stoichiometry to Vegetation Succession in a Karst Region of Southwest China" Forests 10, no. 9: 755. https://doi.org/10.3390/f10090755
APA StyleSong, M., Peng, W., Du, H., & Xu, Q. (2019). Responses of Soil and Microbial C:N:P Stoichiometry to Vegetation Succession in a Karst Region of Southwest China. Forests, 10(9), 755. https://doi.org/10.3390/f10090755