Arbuscular Mycorrhizal Fungi Effectively Enhances the Growth of Gleditsia sinensis Lam. Seedlings under Greenhouse Conditions

Abstract
:
1. Introduction
2. Material and Methods
2.1. Plant Seeds, AMF Inocula and Soil
2.2. Experimental Design
2.3. Seedling Growth Parameters
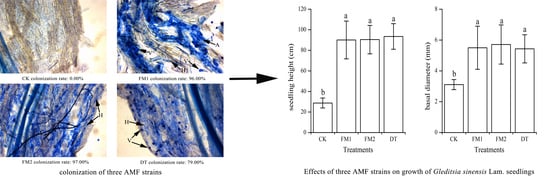
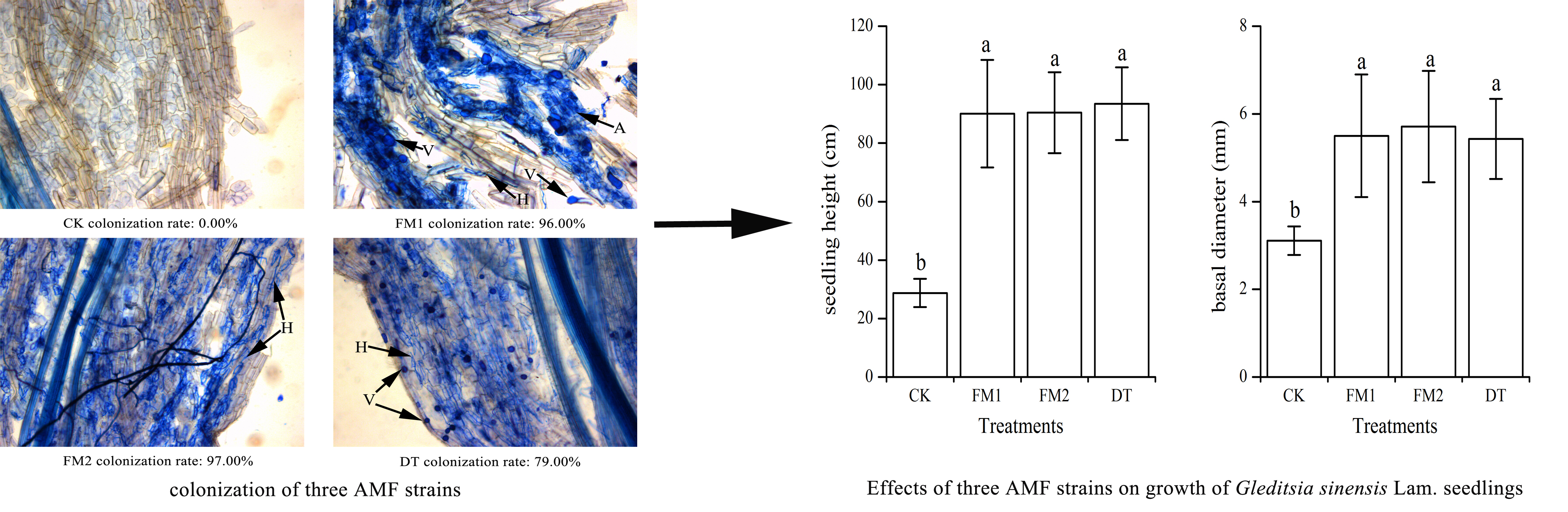
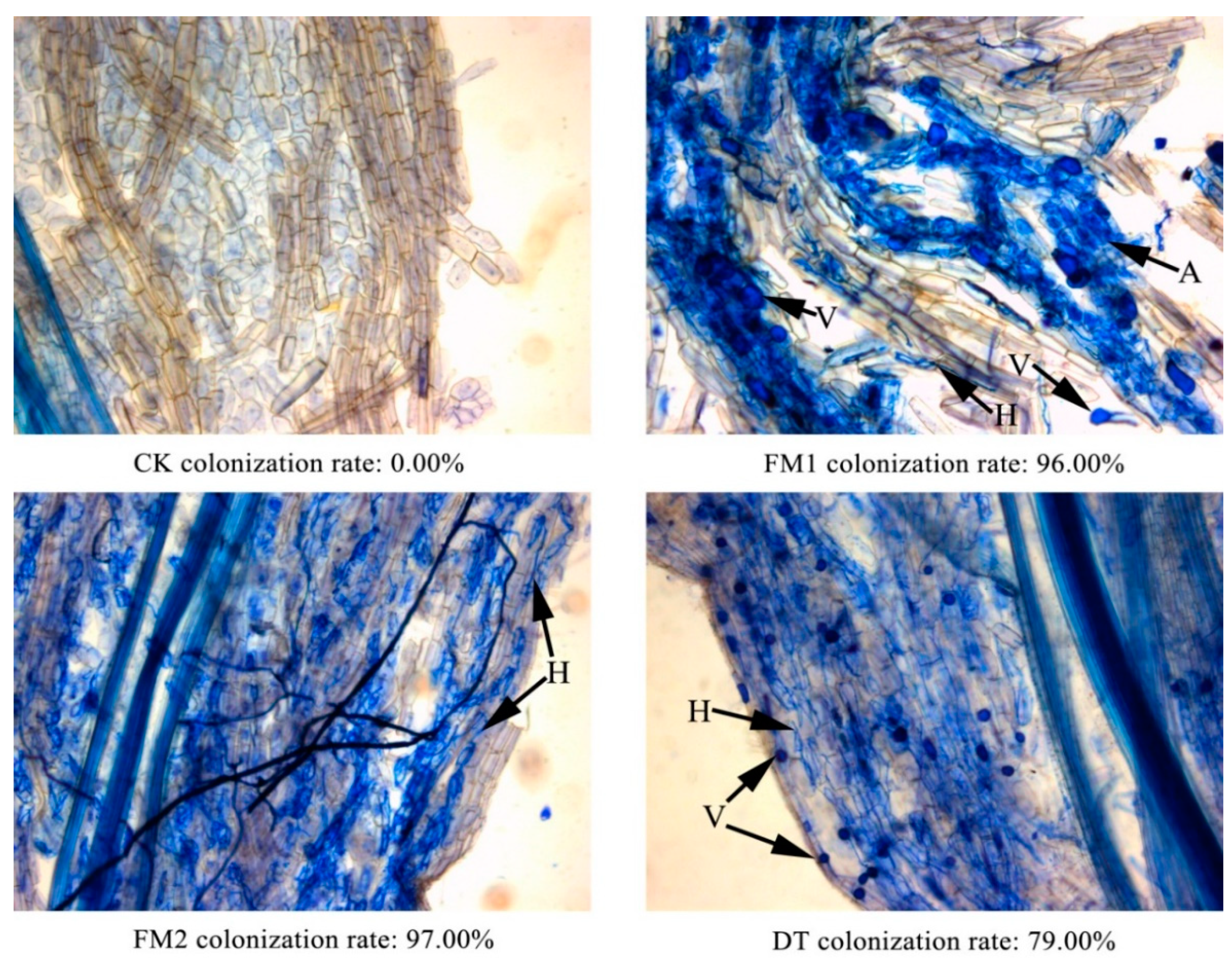
2.4. Root Mycorrhizal Colonization
2.5. Photosynthetic Pigments and Photosynthetic Rate
2.6. Nutrient Contents in Different Organs
2.7. Statistical Analysis
3. Results
3.1. Mycorrhizal Colonization and Plant Growth Parameters
3.2. Plant Biomass and Mycorrhizal Dependence
3.3. Photosynthetic Pigments and Photosynthesis
3.4. Nitrogen, Phosphorus and Potassium Contents
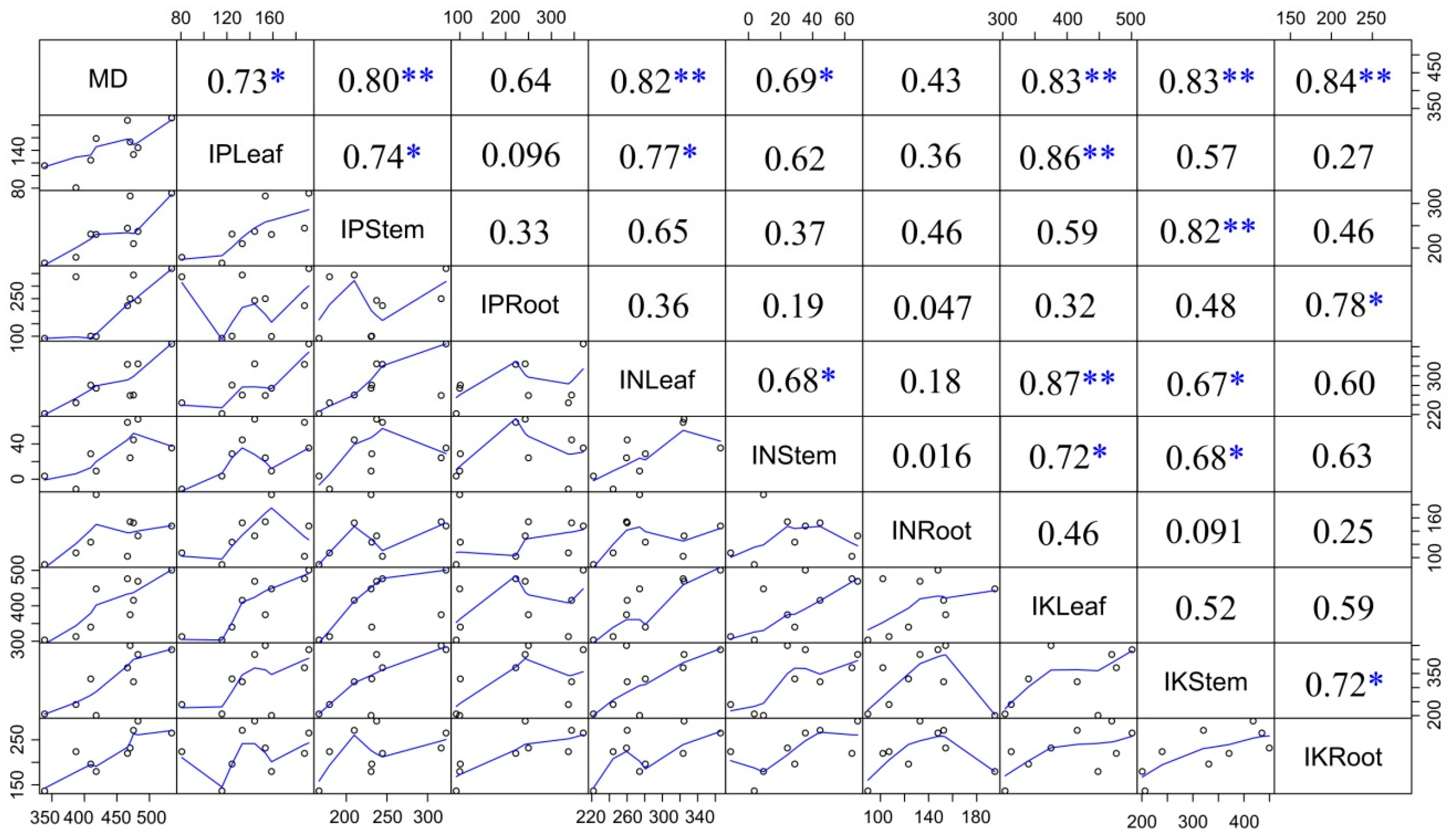
3.5. Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, J.P.; Tian, X.H.; Yang, Y.X.; Liu, Q.X.; Wang, Q.; Chen, L.P.; Li, H.L.; Zhang, W.D. Gleditsia species: An ethnomedical, phytochemical and pharmacological review. J. Ethnopharmacol. 2016, 178, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Lan, Y.P.; Zhou, L.D.; Li, S.Y.; Cao, Q.C.; Lan, W.Z. Advances in research of Gleditsia and its prospect of industrializational development. World For. Res. 2004, 17, 17–21. (In Chinese) [Google Scholar]
- Mei, X.; Ju, L.X.; Fang, C.H.; Li, J.H.; Bi, S.Q.; Chen, Y.; Zou, C.; Zhou, C.; Ma, L.; Miao, L.T. Promoting the hydrolysis and acidification of rice straw by adding Gleditsia sinensis pod powder. Ind. Crops Prod. 2017, 100, 35–40. [Google Scholar] [CrossRef]
- Wu, J.; Li, J.; Zhu, Z.; Li, J.; Huang, G.; Tang, Y.; Gao, X. Protective effects of echinocystic acid isolated from Gleditsia sinensis Lam. against acute myocardial ischemia. Fitoterapia 2010, 81, 8–10. [Google Scholar] [CrossRef]
- Shaheen, U.; Ragab, E.A.; Abdalla, A.N.; Bader, A. Triterpenoidal saponins from the fruits of Gleditsia caspica with proapoptotic properties. Phytochemistry 2018, 145, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Larrainzar, E.; Wienkoop, S. A proteomic view on the role of legume symbiotic interactions. Front. Plant Sci. 2017, 8, 1267. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; Huang, Y.; Zhu, C.; Zeng, H.; Shen, C.; Liu, C.; Zhao, Y.; Pi, E. Rhizobia-inoculation enhances the soybean’s tolerance to salt stress. Plant Soil 2016, 400, 209–222. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Srinivas, V.; Samineni, S. Nitrogen fixation, plant growth and yield enhancements by diazotrophic growth-promoting bacteria in two cultivars of chickpea (Cicer arietinum L.). Biocatal. Agric. Biotechnol. 2017, 11, 116–123. [Google Scholar] [CrossRef]
- Pagano, M. Mycorrhiza: Occurrence in Natural and Restored Environments; Nova Science: New York, NY, USA, 2010; pp. 147–163. [Google Scholar]
- Marschner, H.; Dell, B. Nutrient uptake in mycorrhizal symbiosis. Plant Soil 1994, 159, 89–102. [Google Scholar] [CrossRef]
- Schüßler, A.; Schwarzott, D.; Walker, C. A new fungal phylum, the Glomeromycota: Phylogeny and evolution. Mycol. Res. 2001, 105, 1413–1421. [Google Scholar] [CrossRef]
- Bulgarelli, R.G.; Marcos, F.C.C.; Ribeiro, R.V.; der Andrade, S.A.L. Mycorrhizae enhance nitrogen fixation and photosynthesis in phosphorus-starved soybean (Glycine max L. Merrill). Environ. Exp. Bot. 2017, 140, 26–33. [Google Scholar] [CrossRef]
- Evelin, H.; Giri, B.; Kapoor, R. Contribution of Glomus intraradices inoculation to nutrient acquisition and mitigation of ionic imbalance in NaCl-stressed Trigonella foenum-graecum. Mycorrhiza 2012, 22, 203–217. [Google Scholar] [CrossRef] [PubMed]
- Garg, N.; Pandey, R. Effectiveness of native and exotic arbuscular mycorrhizal fungi on nutrient uptake and ion homeostasis in salt-stressed Cajanus cajan L. (Millsp.) genotypes. Mycorrhiza 2015, 25, 165–180. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.H.; Syvertsen, J.P. Host determinants of mycorrhizal dependency of citrus rootstock seedlings. New Phytol. 2010, 101, 667–676. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161, IN16–IN18. [Google Scholar] [CrossRef]
- Biermann, B.; Linderman, R.G. Quantifying vesicular-arbuscular mycorrhizae: A proposed method towards standardization. New Phytol. 1981, 87, 63–67. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, M.; Ding, Y.; Zhou, P.; Fang, Y.M. Composition of photosynthetic pigments and photosynthetic characteristics in green and yellow sectors of the variegated Aucuba japonica ‘Variegata’ leaves. Flora 2018, 240, 25–33. [Google Scholar] [CrossRef]
- Wang, J.P.; Fu, Z.Y.; Ren, Q.; Zhu, L.J.; Lin, J.; Zhang, J.C.; Cheng, X.F.; Ma, J.Y.; Yue, J.M. Effects of arbuscular mycorrhizal fungi on growth, photosynthesis, and nutrient uptake of Zelkova serrata (Thunb.) Makino seedlings under salt stress. Forests 2019, 10, 186. [Google Scholar] [CrossRef]
- Allen, M.F. Mycorrhizae and rehabilitation of disturbed arid soils: Processes and practices. Arid Soil Res. Rehabil. 1989, 3, 229–241. [Google Scholar] [CrossRef]
- Giri, B.; Kapoor, R.; Mukerji, K.G. Influence of arbuscular mycorrhizal fungi and salinity on growth, biomass, and mineral nutrition of Acacia auriculiformis. Biol. Fertil. Soils 2003, 38, 170–175. [Google Scholar] [CrossRef]
- Giri, B.; Kapoor, R.; Mukerji, K.G. Improved tolerance of Acacia nilotica to salt stress by arbuscular mycorrhiza, Glomus fasciculatum may be partly related to elevated K/Na ratios in root and shoot tissues. Microb. Ecol. 2007, 54, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.F.; Zhang, J.C.; Xu, G.P.; Zhou, L.W.; Li, Y.Q. Arbuscular mycorrhizal fungi improve the growth and drought tolerance of Zenia insignis seedlings under drought stress. New For. 2018, 1–12. [Google Scholar] [CrossRef]
- Lu, N.; Zhou, X.; Cui, M.; Yu, M.; Zhou, J.X.; Qin, Y.S.; Li, Y. Colonization with arbuscular mycorrhizal fungi promotes the growth of Morus alba L. seedlings under greenhouse conditions. Forests 2015, 6, 734–747. [Google Scholar] [CrossRef]
- Smith, F.A.; Smith, S.E. Mutualism and parasitism: Diversity in function and structure in the “Arbuscular” (VA) mycorrhizal symbiosis. Adv. Bot. Res. 1996, 22, 1–43. [Google Scholar]
- Jiang, J.; Moore, J.A.M.; Priyadarshi, A.; Classen, A.T. Plant-mycorrhizal interactions mediate plant community coexistence by altering resource demand. Ecology 2017, 98, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Gerdemann, J.W. Vesicular-arbuscular mycorrhizae. In The Development and Function of Roots; Torrey, J.G., Clarkson, D.T., Eds.; Academic Press: London, UK, 1975; pp. 575–591. [Google Scholar]
- Lu, Y.W.; Wang, G.Q.; Meng, Q.J.; Zhang, W.H.; Duan, B.L. Growth and physiological responses to arbuscular mycorrhizal fungi and salt stress in dioecious plant Populus tomentosa. Can. J. For. Res. 2014, 44, 1020–1031. [Google Scholar] [CrossRef]
- Frosi, G.; Barros, V.A.; Oliveira, M.T.; Santos, M.; Ramos, D.G.; Maia, L.C.; Santos, M.G. Arbuscular mycorrhizal fungi and foliar phosphorus inorganic supply alleviate salt stress effects in physiological attributes, but only arbuscular mycorrhizal fungi increase biomass in woody species of a semiarid environment. Tree Physiol. 2018, 38, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Koltai, H.; Kapulnik, Y. Arbuscular Mycorrhizas: Physiology and Function; Springer: Dordrecht, The Netherlands, 2010; pp. 57–71. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Treatments | Seedling Height (cm) | Basal Diameter (mm) | Dry Weight (g/pot) | Mycorrhizal Dependence | ||||
|---|---|---|---|---|---|---|---|---|
| July | September | July | September | Shoot | Root | Total | ||
| CK | 13.24 ± 2.69b | 28.73 ± 4.82b | 2.34 ± 0.27b | 3.11 ± 0.32b | 3.47 ± 0.53b | 2.46 ± 0.31b | 5.92 ± 0.72c | 0 ± 0c |
| FM1 | 42.34 ± 17.16a | 90.06 ± 18.39a | 3.50 ± 0.85a | 5.50 ± 1.40a | 21.29 ± 4.60a | 9.04 ± 0.63a | 30.33 ± 4.82ab | 463.82 ± 73.78ab |
| FM2 | 44.83 ± 12.53a | 90.43 ± 13.85a | 3.64 ± 0.61a | 5.71 ± 1.27a | 21.52 ± 1.65a | 9.52 ± 1.95a | 31.04 ± 0.53a | 474.68 ± 8.08a |
| DT | 45.71 ± 9.15a | 93.48 ± 12.44a | 3.37 ± 0.55a | 5.43 ± 0.92a | 17.76 ± 1.56a | 7.67 ± 1.45a | 25.43 ± 2.85b | 388.86 ± 43.55b |
| Treatments | Chl a (mg/g) | Chl b (mg/g) | Chl (mg/g) | Pn (µmol m−2 s−1) | Gs (mmol m−2 s−1) | Tr (mmol m−2 s−1) |
|---|---|---|---|---|---|---|
| CK | 1.82 ± 0.40a | 0.538 ± 0.206a | 2.35 ± 0.58a | 11.54 ± 2.03a | 0.127 ± 0.029a | 2.36 ± 0.37b |
| FM1 | 2.06 ± 0.29a | 0.446 ± 0.062a | 2.50 ± 0.35a | 13.32 ± 2.95a | 0.132 ± 0.053a | 3.63 ± 1.28a |
| FM2 | 2.10 ± 0.15a | 0.474 ± 0.087a | 2.58 ± 0.24a | 13.90 ± 1.88a | 0.169 ± 0.058a | 3.94 ± 1.02a |
| DT | 2.04 ± 0.17a | 0.416 ± 0.050a | 2.46 ± 0.21a | 12.92 ± 2.98a | 0.130 ± 0.067a | 3.64 ± 1.47a |
| Nutrient | Organ | CK | FM1 | FM2 | DT | |||
|---|---|---|---|---|---|---|---|---|
| (mg per pot) | (mg per pot) | Increase (%) | (mg per pot) | Increase (%) | (mg per pot) | Increase (%) | ||
| P | Leaf | 2.82 ± 0.17b | 6.82 ± 1.58a | 141.83 ± 56.16 | 7.20 ± 0.81a | 155.02 ± 28.67 | 6.57 ± 0.64a | 132.98 ± 22.79 |
| Stem | 2.20 ± 0.24b | 8.20 ± 1.78a | 272.88 ± 81.05 | 7.27 ± 0.40a | 230.52 ± 18.30 | 6.81 ± 0.82a | 209.42 ± 37.19 | |
| Root | 2.37 ± 0.16c | 9.95 ± 1.47a | 318.95 ± 62.02 | 8.78 ± 1.55a | 269.72 ± 65.34 | 4.69 ± 0.11b | 97.386 ± 4.79 | |
| Total | 7.40 ± 0.35c | 24.98 ± 3.54a | 237.67 ± 47.79 | 23.25 ± 0.75a | 214.30 ± 10.19 | 18.07 ± 1.43b | 144.29 ± 19.33 | |
| N | Leaf | 41.60 ± 1.73b | 162.14 ± 27.56a | 289.80 ± 66.26 | 167.53 ± 15.35a | 302.76 ± 36.90 | 149.31 ± 13.42a | 258.95 ± 32.25 |
| Stem | 75.65 ± 1.37b | 87.88 ± 18.40b | 16.17 ± 24.32 | 120.31 ± 9.55a | 59.04 ± 12.63 | 86.17 ± 10.02b | 13.906 ± 13.24 | |
| Root | 50.29 ± 3.15b | 118.88 ± 12.82a | 136.37 ± 25.49 | 115.24 ± 12.81a | 129.13 ± 25.47 | 118.69 ± 27.11a | 135.99 ± 53.91 | |
| Total | 167.54 ± 3.71b | 368.89 ± 53.16a | 120.19 ± 31.73 | 403.08 ± 17.34a | 140.59 ± 10.35 | 354.16 ± 41.41a | 111.39 ± 24.71 | |
| K | Leaf | 22.81 ± 1.08b | 113.20 ± 21.82a | 396.35 ± 95.67 | 126.30 ± 7.58a | 453.79 ± 33.24 | 105.79 ± 17.12a | 363.88 ± 75.08 |
| Stem | 16.07 ± 0.51b | 76.30 ± 18.83a | 374.80 ± 117.20 | 75.49 ± 7.82a | 369.77 ± 48.67 | 55.60 ± 11.84a | 245.97 ± 73.70 | |
| Root | 32.64 ± 3.52c | 111.06 ± 7.15a | 240.23 ± 21.92 | 117.81 ± 12.07a | 260.91 ± 36.97 | 88.41 ± 10.06b | 170.85 ± 30.81 | |
| Total | 71.52 ± 4.01c | 300.56 ± 44.01ab | 320.25 ± 61.54 | 319.60 ± 18.67a | 346.88 ± 26.10 | 249.80 ± 27.15b | 249.28 ± 37.96 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Zhong, H.; Zhu, L.; Yuan, Y.; Xu, L.; Wang, G.G.; Zhai, L.; Yang, L.; Zhang, J. Arbuscular Mycorrhizal Fungi Effectively Enhances the Growth of Gleditsia sinensis Lam. Seedlings under Greenhouse Conditions. Forests 2019, 10, 567. https://doi.org/10.3390/f10070567
Wang J, Zhong H, Zhu L, Yuan Y, Xu L, Wang GG, Zhai L, Yang L, Zhang J. Arbuscular Mycorrhizal Fungi Effectively Enhances the Growth of Gleditsia sinensis Lam. Seedlings under Greenhouse Conditions. Forests. 2019; 10(7):567. https://doi.org/10.3390/f10070567
Chicago/Turabian StyleWang, Jinping, Huini Zhong, Lingjun Zhu, Yingdan Yuan, Linhao Xu, G. Geoff Wang, Lu Zhai, Lu Yang, and Jinchi Zhang. 2019. "Arbuscular Mycorrhizal Fungi Effectively Enhances the Growth of Gleditsia sinensis Lam. Seedlings under Greenhouse Conditions" Forests 10, no. 7: 567. https://doi.org/10.3390/f10070567
APA StyleWang, J., Zhong, H., Zhu, L., Yuan, Y., Xu, L., Wang, G. G., Zhai, L., Yang, L., & Zhang, J. (2019). Arbuscular Mycorrhizal Fungi Effectively Enhances the Growth of Gleditsia sinensis Lam. Seedlings under Greenhouse Conditions. Forests, 10(7), 567. https://doi.org/10.3390/f10070567