Reasons for the Extremely Small Population of putative hybrid Sonneratia × hainanensis W.C. Ko (Lythraceae)



Abstract
1. Introduction
2. Materials and Methods
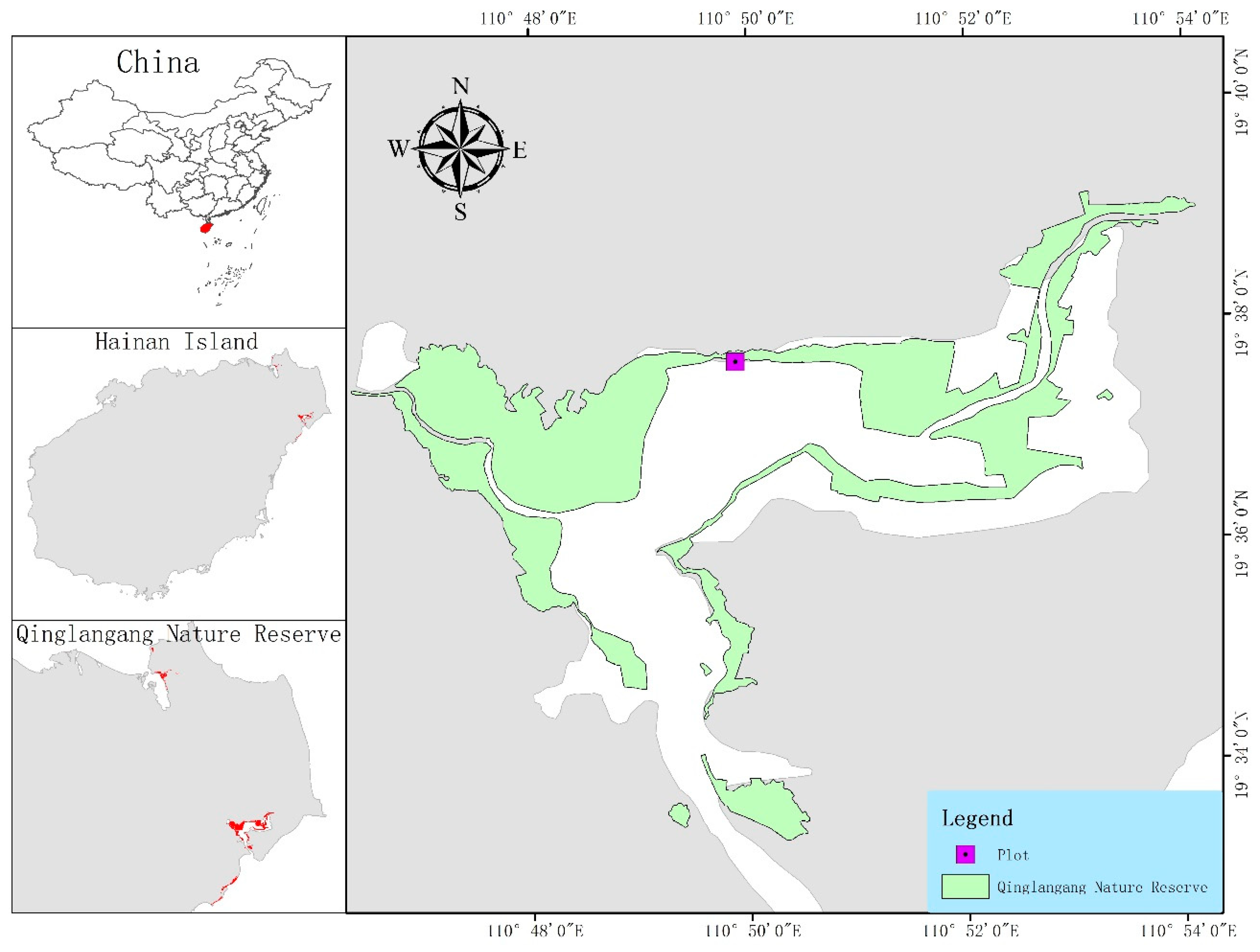
2.1. An Overview of the Sample Plot
2.2. Study Object
2.3. Determination of the Reproductive System
2.3.1. Estimating the Outcrossing Index (OCI)
2.3.2. Pollen to Ovule (P/O) ratio
2.4. Determination of the Reproductive System
2.4.1. Changes in Stigma Receptivity and Pollen Vitality with Time
2.4.2. Artificial Pollination and Bagging Experiments
2.5. Seed Germination and Seedling Survival Experiment
2.5.1. Dynamic Changes in S. × hainanensis Seeds under Natural Conditions
2.5.2. The Relationship between S. × hainanensis Seed Germination and Environmental Factors
2.6. Data Analysis Method
3. Results
3.1. Determination of the Reproductive System
3.2. Exploring Dysgenesis in S. × hainanensis
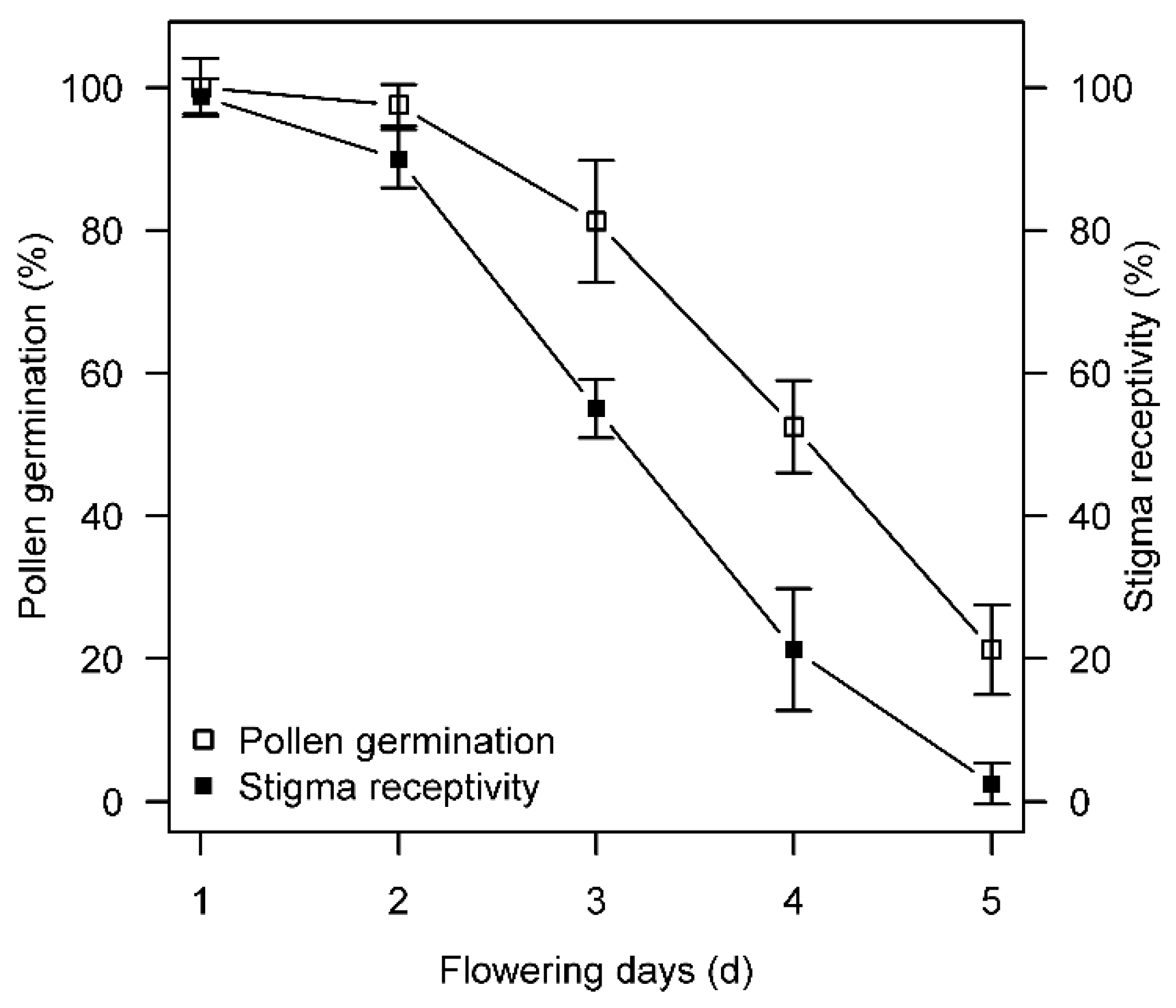
3.2.1. Pollen Germination Rate and Stigma Receptivity
3.2.2. Artificial Pollination Experiment
3.3. Relationships among Seed Germination, Survival of Seedlings, and Environmental Factors
3.3.1. Seed Germination Rate at Different Fruit Dropping Times
3.3.2. Dynamic Observation and Verification of Seeds under Natural Conditions
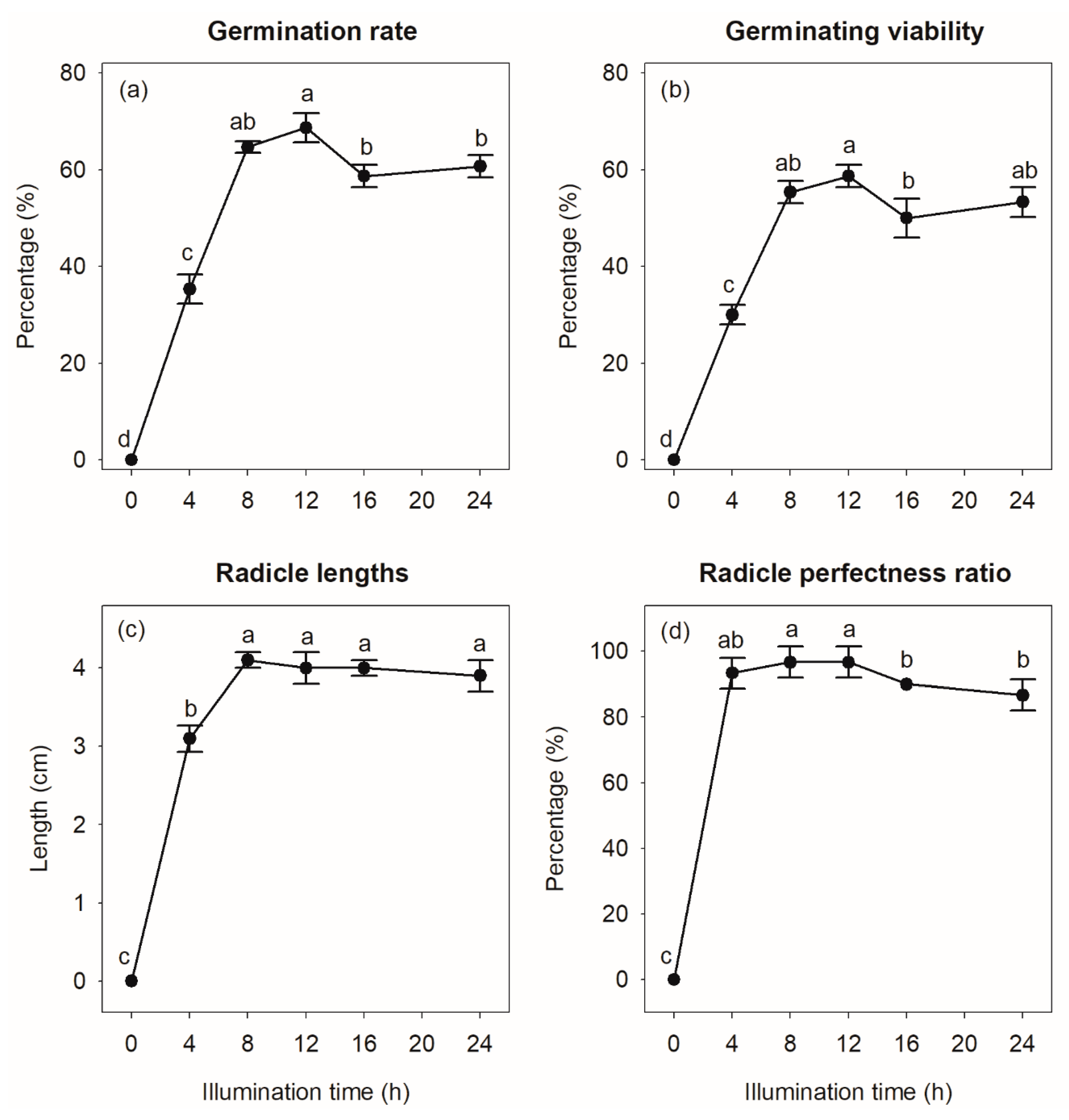
3.3.3. Effects of Light Cycle Duration on S. × hainanensis Seed Sprouting
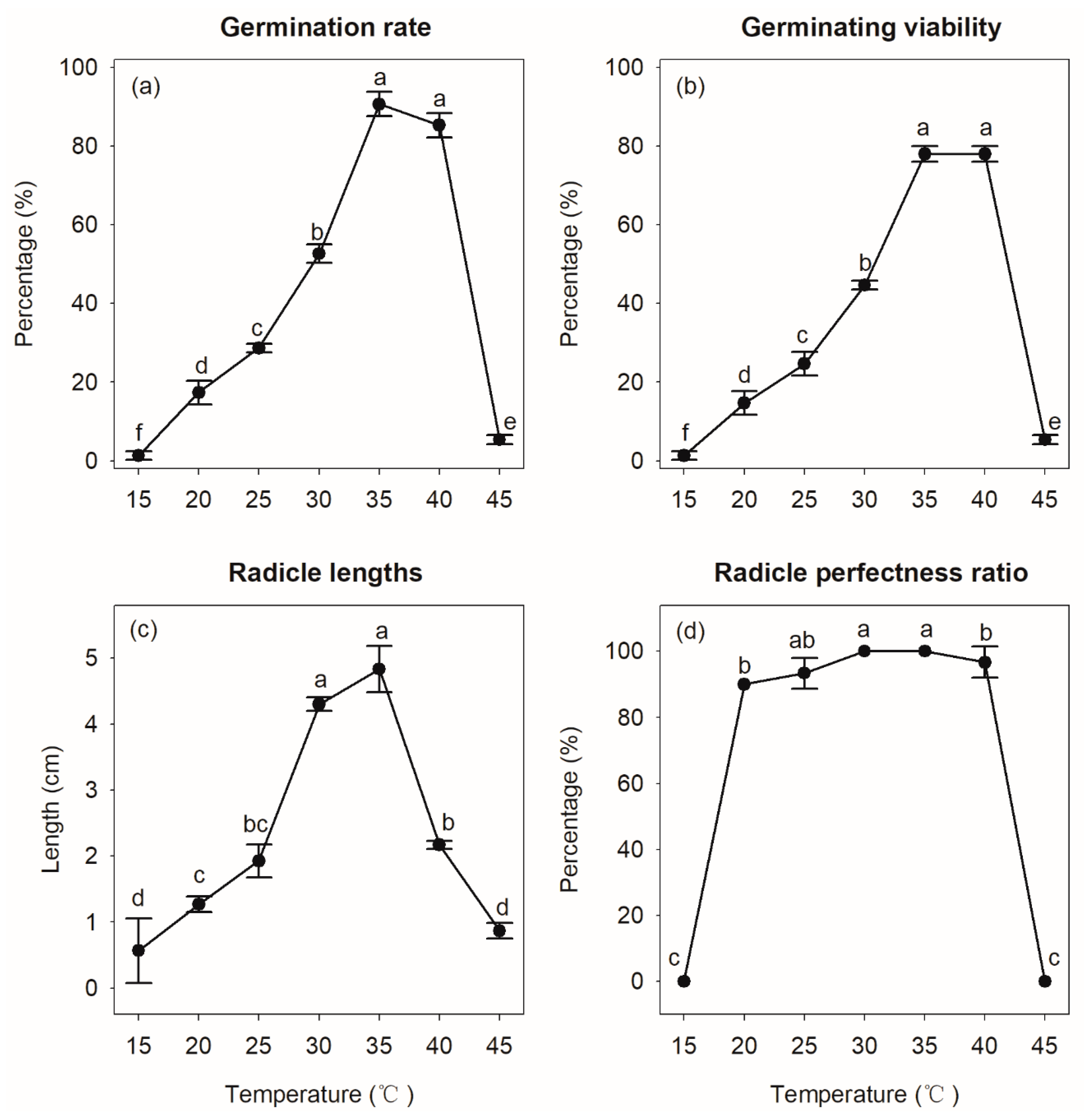
3.3.4. Effect of Temperature on S. × hainanensis Sprouting
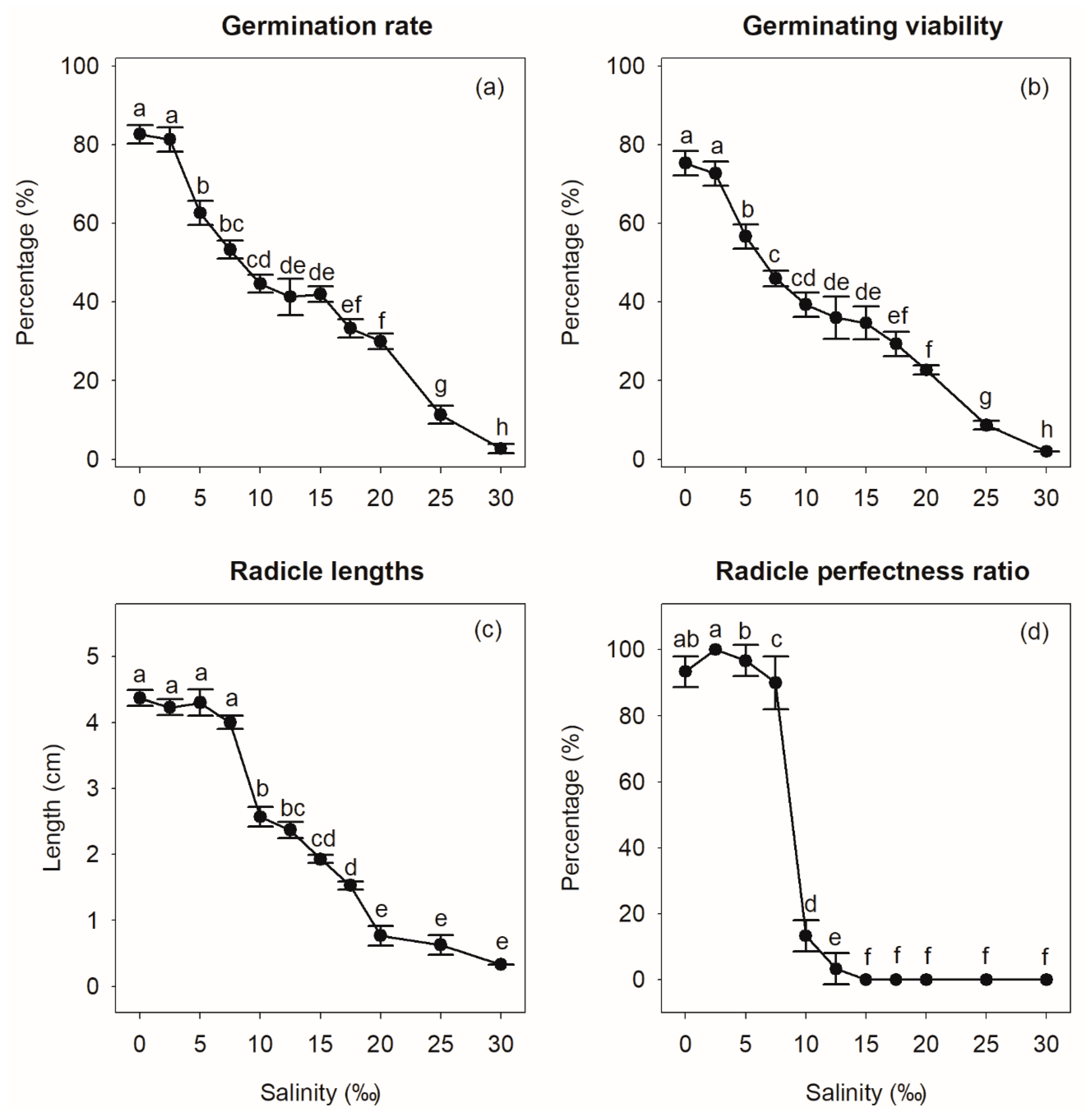
3.3.5. Effect of Salinity on S. × hainanensis Seed Germination
4. Discussion
4.1. Reproductive System of S. × hainanensis
4.2. Effects of Natural Environment on S. × hainanensis Seed Germination
4.3. Effects of Simulated Environmental Factors on S. × hainanensis Seed Germination
4.3.1. Effect of Light on S. × hainanensis Seed Germination
4.3.2. Effect of Temperature on S. × hainanensis Seed Germination
4.3.3. Effect of Salinity on S. × hainanensis Seed Germination
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Arroyo, J.; Thompson, J.D. Plant reproductive ecology and evolution in a changing Mediterranean climate. Plant Boil. 2018, 20, 3–7. [Google Scholar] [CrossRef]
- Sheng, M.Y.; Shen, C.Z.; Chen, X.; Tian, X.J. Resource status and protection countermeasures of endangered wild plants in China. Chin. J. Nat. 2011, 33, 455–467. [Google Scholar]
- Liu, K.Q.; Deng, H.P. Floral biology and breeding system of endangered plant Scutellaria tsinyunensis endemic to Chongqing, China. Bull. Bot. Res. 2011, 31, 403–407. [Google Scholar]
- Sigrist, M.R.; Sazima, M. Phenology, reproductive biology and diversity of buzzing bees of sympatric Dichorisandra species (Commelinaceae): breeding system and performance of pollinators. Plant Syst. Evol. 2014, 301, 1005–1015. [Google Scholar] [CrossRef]
- Tsuji, K.; Ohgushi, T. Florivory indirectly decreases the plant reproductive output through changes in pollinator attraction. Ecol. Evol. 2018, 8, 2993–3001. [Google Scholar] [CrossRef]
- Waelti, M.O.; Muhlemann, J.K.; Widmer, A.; Schiestl, F.P. Floral odour and reproductive isolation in two species of Silene. J. Revolut. Biol. 2008, 21, 111–121. [Google Scholar] [CrossRef]
- Gross, C.L.; Bartier, F.V.; Mulligan, D.R. Floral structure, breeding system and fruit-set in the threatened sub-shrub Tetratheca juncea Smith (Tremandraceae). Ann. Bot. 2003, 92, 771–777. [Google Scholar] [CrossRef][Green Version]
- Hamston, T.J.; Wilson, R.J.; De Vere, N.; Rich, T.C.G.; Stevens, J.R.; Cresswell, J.E. Breeding system and spatial isolation from congeners strongly constrain seed set in an insect–pollinated apomictic tree: Sorbus subcuneata (Rosaceae). Sci. Rep. 2017, 7, 45122. [Google Scholar] [CrossRef]
- Sawyer, N.W. Reproductive Ecology of Trillium recurvatum (Trilliaceae) in Wisconsin. Am. Midl. Nat. 2010, 163, 146–160. [Google Scholar] [CrossRef]
- Opedal, Ø.H.; Armbruster, W.S.; Pélabon, C. Inbreeding effects in a mixed–mating vine: effects of mating history, pollen competition and stress on the cost of inbreeding. AoB PLANTS 2015, 7, 133. [Google Scholar] [CrossRef]
- Takebayashi, N.; Morrell, P.L. Is self–fertilization an evolutionary dead end? Revisiting an old hypothesis with genetic theories and a macro–evolutionary approach. Am. J. Bot. 2001, 88, 1143–1150. [Google Scholar] [CrossRef] [PubMed]
- Bebawi, F.F.; Campbell, S.D.; Mayer, R.J. Seed ecology of Captain Cook tree [Cascabela thevetia (L.) Lippold] – germination and longevity. Rangel. J. 2017, 39, 307. [Google Scholar] [CrossRef]
- Oakley, C.G.; Ågren, J.; Schemske, D.W. Heterosis and outbreeding depression in crosses between natural populations of Arabidopsis thaliana. Heredity 2015, 115, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Prill, N.; Bullock, J.M.; Van Dam, N.M.; Leimu, R. Loss of heterosis and family–dependent inbreeding depression in plant performance and resistance against multiple herbivores under drought stress. J. Ecol. 2014, 102, 1497–1505. [Google Scholar] [CrossRef]
- Jiménez-Alfaro, B.; Silveira, F.A.; Fidelis, A.; Poschlod, P.; Commander, L.E. Seed germination traits can contribute better to plant community ecology. J. Veg. Sci. 2016, 27, 637–645. [Google Scholar] [CrossRef]
- Phartyal, S.S.; Rosbakh, S.; Poschlod, P. Seed germination ecology in Trapa natans L., a widely distributed freshwater macrophyte. Aquat. Bot. 2018, 147, 18–23. [Google Scholar] [CrossRef]
- Zhang, S.X. A preliminary study on cutting propagation techniques for old plants of Cinnamomum micranthum. J. Fujian Sci. Tech. 2000, 27, 69–71. [Google Scholar]
- Xie, Z.Q.; Chen, W.L. The endangered causes and protection measures of Cathaya argyrophylla, an endemic to China. Acta Phytoecol. Sin. 2003, 27, 661–666. [Google Scholar]
- Gao, Y.Z. Sonneratiaceae: The Flora of China; Science Press: Beijing, China, 1983; pp. 114–116. [Google Scholar]
- Lin, P.; Lu, C.Y. Mangrove community in Hainan Island. J. Xiamen Univ. 1985, 24, 1117–1119. [Google Scholar]
- Li, H.S.; Chen, G.Z. Biological characteristics and protection of Chinese endemic plant S. × hainanensis. J. Guangdong Educ. Inst. 2003, 23, 48–51. [Google Scholar]
- Zhang, Z.; Guo, Y.; He, J.-S.; Tang, Z. Conservation status of wild plant species with extremely small populations in China. Biodivers. Sci. 2018, 26, 572–577. [Google Scholar] [CrossRef]
- Duke, N.C. A mangrove hybrid, Sonneratia × gulngai (Sonneratiaceae) from north-eastern Australia. Austrobaileya 1984, 2, 103–105. [Google Scholar]
- Tomlinson, P.B. The botany of Mangroves; Cambridge University Press: Cambridge, UK, 1986. [Google Scholar]
- Zhang, Y.L.; Wang, K.F.; Li, Z. Study on pollen morphology(Sonneratiaceae) in China and Its paleoecological significance. Mar. Geol. Quat. Geol. 1997, 17, 2993–3001. [Google Scholar]
- Renchao, Z. Natural Hybridization and Speciation in Sonneratia. Ph.D. Thesis, Zhongshan University, Guangzhou, China, 2006. [Google Scholar]
- Li, H.S.; Chen, G.Z.; Si, S.H. ISSR study on genetic diversity of Hainan Sanghai. Acta Sci. Nat. Univ. Sunyatseni 2004, 43, 68–70. [Google Scholar]
- Dafni, A. Pollination Ecology; Oxford University Press: New York, NY, USA, 1992. [Google Scholar]
- Cruden, R.W. Pollen-ovule ratios: A conservative indicator of reproductive systems in flowering plants. Evolution 1977, 31, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Holsinger, K.E. Pollination biology and the evolution of mating systems in flowering plants. Evol. Biol. 1996, 29, 107–149. [Google Scholar]
- Morgan, J.W. Effects of population size on seed production and germinability in an endangered, fragmented grassland plant. Conserv. Boil. 1999, 13, 266–273. [Google Scholar] [CrossRef]
- Bie, P.F.; Tang, T.; Hu, J.Y.; Jiang, W. Flowering phenology and breeding system of an endangered and rare species Urophysa rockii(Ranunculaceae). Acta Ecol. Sin. 2018, 38, 3899–3908. [Google Scholar]
- Lande, R. Risk of population extinction from fixation of new deleterious mutations. Evolution 1995, 48, 1460–1469. [Google Scholar] [CrossRef]
- Kéry, M.; Matthies, D.; Spillmann, H.H. Reduced fecundity and offspring performance in small populations of the declining grassland plants Primula veris and Gentiana lutea. J. Ecol. 2000, 88, 17–30. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Y.H.; Zhang, X.N.; Yang, Y. Flower phenology and breeding system of endangered mangrove Lumnitzera littorea (Jack.) Voigt. Chin. J. Appl. Env. Biol. 2017, 23, 0077–0081. [Google Scholar]
- Zhang, J.H.; Liu, B.W. Patterns of seed predation and removal of Mongolian oak by rodents. Acta Ecol. Sin. 2014, 34, 1201–1211. [Google Scholar]
- Yue, H.J.; Tong, C.; Zhu, J.M.; Huang, J.F. Seed rain and soil seed bank of endangered Taxus chinensis var mairei in Fujian, China. Acta Ecol. Sin. 2010, 30, 4389–4400. [Google Scholar]
- Wu, D.R.; Wang, B.S. Seed and seedl ing ecology of the endangered Phoebe bournei (Lauraceae). Acta Ecol. Sin. 1997, 11, 1752–1757. [Google Scholar]
- Manfred, J.; Lesley, P.; Birgitte, S. Habitat specificity, seed germination and experimental translocation of the endangered herb Brachycome muelleri (Asteraceae). Biol. Conserv. 2004, 116, 251–267. [Google Scholar]
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed dormancy and the control of germination. New Phytol. 2006, 171, 501–523. [Google Scholar] [CrossRef]
- Zhang, J.J.; Chai, S.F.; Wei, X.; Lv, S.H.; Wu, S.H. Germination characteristics of the seed of a Rare and endangered plant, Garcinia paucinervis Australia. Sci. Silvae Sin. 2018, 54, 175–178. [Google Scholar]
- Yang, Y.; Zhong, C.R.; Li, Y.H.; Zhang, Y. The morphological structure and germination characters of seed of endangered Mangrove Lumnitzera littorea (Jack.) Voigt. Mol. Plant Breed. 2016, 14, 2851–2858. [Google Scholar]
- Liao, B.W.; Zheng, D.Z.; Zheng, S.F.; Li, Y. Seed germination conditions of Sonneratia caseolaris of mangrove. J. Cent. South For. Univ. 1997, 17, 25–27. [Google Scholar]
- Xu, X.L. Effects of Cold Storage and Temperature on Seed Germination Characteristics of Two Common Plants in Alpine Meadow. Ph.D. Thesis, Lanzhou University, Lanzhou, China, 2007. [Google Scholar]
- Li, W.L.; Zhang, X.P.; Hao, C.Y.; Zhang, H.; Holsinger, K.E. Characteristics of seed germination of the rare plant Cercidiphyllum japonicum. Acta Ecol. Sin. 2008, 28, 107–149. [Google Scholar]
- Chai, S.F.; Jiang, J.S.; Wei, X.; Wang, M.L.; Li, L.; Qi, X.X. Seed germination characteristics of endangered plant Sinia rhodoleuca. Chin. J. Ecol. 2010, 29, 233–237. [Google Scholar]
- Zheng, D.J.; Wu, Y.J.; Yun, Y.; Jiang, D.Q.; Chen, X.; Zhang, Z.L. Seed germination and its environment adaptability of endangered tree Dracaena cambodiana. J. Trop. Subtrop. Bot. 2016, 24, 71–79. [Google Scholar]
- Chen, L.Z.; Wang, W.Q.; Zhang, Y.H.; Huang, L.; Zhao, C.L.; Yang, S.C. Damage to mangroves from extreme cold in early 2008 in sourthern China. Chin. J. Plant Ecol. 2010, 34, 186–194. [Google Scholar]
- You, H.-M. Adaptability of mangrove Kandelia obovata seedlings to salinity–waterlogging. Ying yong sheng tai xue bao = J. Appl. Ecol. 2015, 26, 675–680. [Google Scholar]
- Tang, M.; Li, K.; Xiang, H.Y.; Dong, X.; Jin, H.X.; Wang, Y.; Yang, H.J.; Zhang, Z.X. Research on ecological, physiological and morphological adaptability of two mangrove species to salt stress. Ecol. Sci. 2014, 33, 513–519. [Google Scholar]
- Zhang, Y.; Ye, Y.; Lu, C.Y. Seed germination and seedling growth of mangrove Excoecaria agallocha under different salinities. J. Xiamen Univ. 2010, 49, 145–148. [Google Scholar]
- Khan, M.A.; Gul, B.; Weber, D.J. Germination responses to Salicornia rubra to temperature and salinity. J. Arid Env. 2000, 45, 207–214. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, D.Z.; Liao, B.W.; Zheng, S.F.; Song, X.Y. Effect of salinity and temperature on seed germination of mangrove Sonneratia apetala Buch. Ham. For. Res. 1997, 10, 137–142. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items of Observation | Number of Pollen in Each Simple Flower (P) | Number of Ovules in Each Simple Flower (O) | P/O Ratio | Types of Breeding System |
|---|---|---|---|---|
| Results | ≈12500 ± 2000 | 34 ± 10 | 354 | Xenogamy |
| Pollination Method | Number of Flowers | Fruit Dropping Time (d) | Number of Fruits | Fruiting Rate (%) | Seed Setting Number in a Single Flower |
|---|---|---|---|---|---|
| Net isolation with castration | 40 | 7 | 4 | 10 | 16.25 |
| Bagging with castration | 40 | 0 | 0 | 0 | 0 |
| No bagging treatment | 40 | 7 | 7 | 17.5 | 19 |
| Natural pollination | 40 | 7 | 18 | 45 | 30.4 |
| Artificial self-pollination | 40 | 15 | 17 | 42.5 | 24.2 |
| Artificial geitonogamy | 40 | 15 | 20 | 50 | 34.6 |
| Artificial xenogamy | 40 | 18 | 25 | 62.5 | 56.2 |
| Dropping Time | 5 d | 10 d | 15 d | 20 d | 25 d | 30 d | 35d |
|---|---|---|---|---|---|---|---|
| Fruit dropping amount | 14.67 ± 4.51f | 25.67 ± 4.04e | 39.33 ± 9.24d | 58.00 ± 8.00c | 70.33 ± 4.51ab | 84.67 ± 6.43a | 0 ± 0g |
| Seed germination rate (%) | 0 ± 0d | 0 ± 0d | 2 ± 0bc | 9 ± 0b | 43 ± 3a | 45 ± 3a | 0 ± 0d |
| Sampling Point | Fruit Predation Rate (%) | Seed Germination Rate (%) | Seed Damage Rate (%) | Seed Remaining Rate (%) |
|---|---|---|---|---|
| 1 m | 100 | 0 | 83 ± 10 | 17 ± 10 |
| 3 m | 100 | 0 | 85 ± 0 | 15 ± 0 |
| 5 m | 100 | 0 | 80 ± 6 | 20 ± 6 |
| 10 m | 100 | 0 | 82 ± 13 | 18 ± 13 |
| Average | 100 | 0 | 82.5 ± 2.08 | 17.5 ± 2.08 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Yang, X.; Long, W.; Li, D.; Lv, X. Reasons for the Extremely Small Population of putative hybrid Sonneratia × hainanensis W.C. Ko (Lythraceae). Forests 2019, 10, 526. https://doi.org/10.3390/f10060526
Zhang M, Yang X, Long W, Li D, Lv X. Reasons for the Extremely Small Population of putative hybrid Sonneratia × hainanensis W.C. Ko (Lythraceae). Forests. 2019; 10(6):526. https://doi.org/10.3390/f10060526
Chicago/Turabian StyleZhang, Mengwen, Xiaobo Yang, Wenxing Long, Donghai Li, and Xiaobo Lv. 2019. "Reasons for the Extremely Small Population of putative hybrid Sonneratia × hainanensis W.C. Ko (Lythraceae)" Forests 10, no. 6: 526. https://doi.org/10.3390/f10060526
APA StyleZhang, M., Yang, X., Long, W., Li, D., & Lv, X. (2019). Reasons for the Extremely Small Population of putative hybrid Sonneratia × hainanensis W.C. Ko (Lythraceae). Forests, 10(6), 526. https://doi.org/10.3390/f10060526