The Importance of Moisture for Brown Rot Degradation of Modified Wood: A Critical Discussion

{kind=link}
Abstract
1. Introduction
1.1. Brown Rot Wood Degradation
1.2. Wood Modification
1.3. Wood-Water Relationships in Modified Wood
1.4. Aim of the Review
2. Fungal Response to Wood Modification and Inhibition of Decay
3. Diffusion in Modified Wood
3.1. Nanopore Blocking
3.2. Media of Transport and Percolating Network
3.2.1. Capillary Water Formation in Nanopores
3.2.2. Plasticised Hemicelluloses
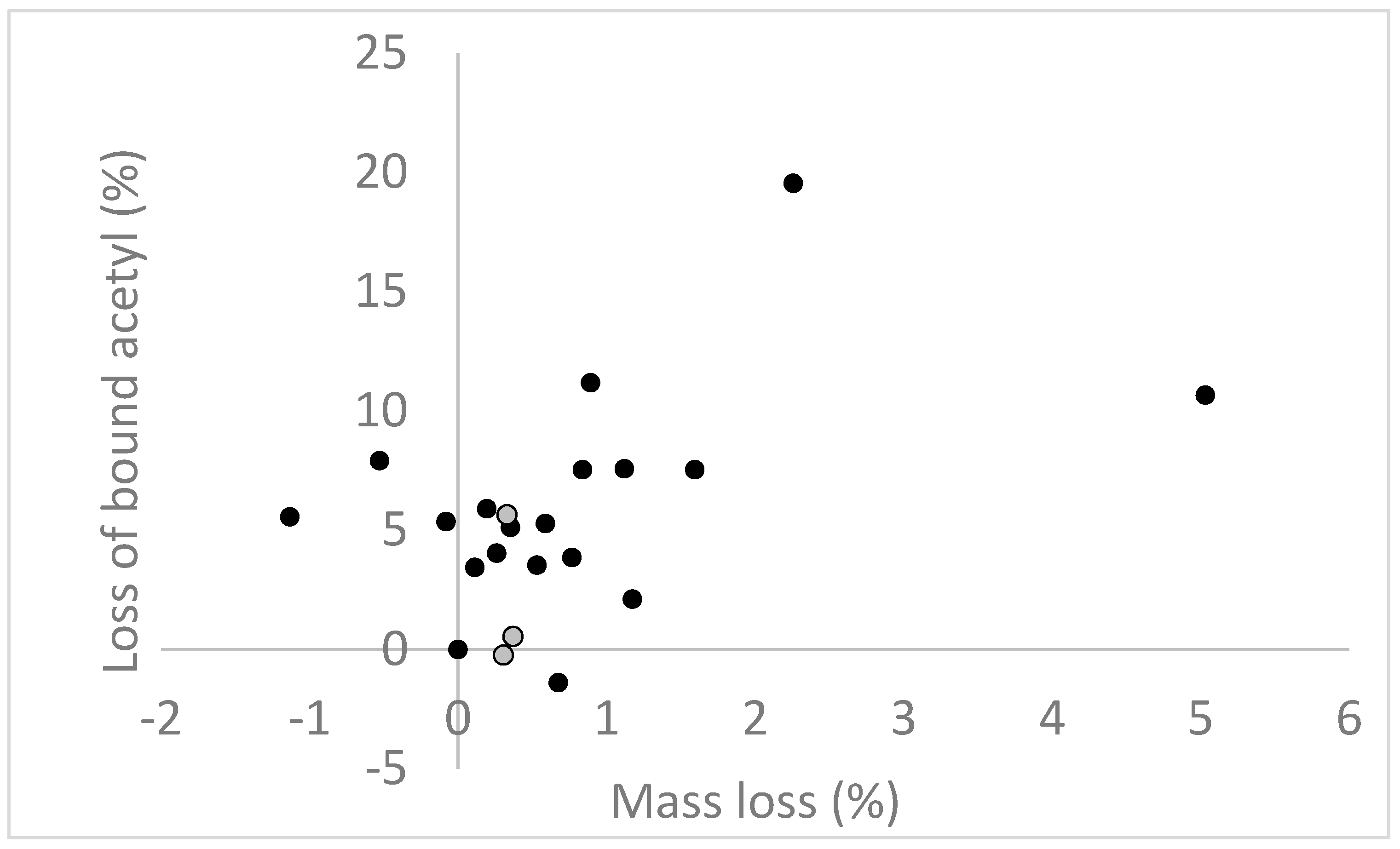
4. Moisture Relationships in Modified Wood during Decay
5. Controlling the Moisture Conditions during Decay Tests: A Challenge
6. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Eaton, R.A.; Hale, M.D.C. Wood: Decay, Pests and Protection; Chapman & Hall: Cambridge, UK, 1993. [Google Scholar]
- Ibach, R. Biological properties. In Wood Chemistry and Wood Composites; Rowell, R.M., Ed.; Taylor & Francis: Boca Raton, FL, USA, 2005. [Google Scholar]
- Morrell, J.J. Protection of wood based materials. In Handbook of Environmental Degradation of Materials; Kutz, M., Ed.; William Andrew: Norwich, NY, USA, 2005; pp. 299–317. [Google Scholar]
- IPCC. Climate Change 2013; IPCC: Budapest, Hungary, 2013. [Google Scholar]
- Alfredsen, G.; Solheim, H.; Jenssen, K. Råtesopp i norske bygninger. Agarica 2006, 26, 78–86. [Google Scholar]
- Gabriel, J.; Švec, K. Occurrence of indoor wood decay basidiomycetes in Europe. Fungal Biol. Rev. 2017, 31, 212–217. [Google Scholar] [CrossRef]
- Schwarze, F.W.M.R. Wood decay under the microscope. Fungal Biol. Rev. 2007, 21, 133–170. [Google Scholar] [CrossRef]
- Yelle, D.J.; Wei, D.; Ralph, J.; Hammel, K.E. Multidimensional NMR analysis reveals truncated lignin structures in wood decayed by the brown rot basidiomycete Postia Placenta. Environ. Microbiol. 2011, 13, 1091–1100. [Google Scholar] [CrossRef] [PubMed]
- Arantes, V.; Milagres, A.M.F.; Filley, T.R.; Goodell, B. Lignocellulosic polysaccharides and lignin degradation by wood decay fungi: The relevance of nonenzymatic Fenton-based reactions. J. Ind. Microbiol. Biotechnol. 2011, 38, 541–555. [Google Scholar] [CrossRef] [PubMed]
- Winandy, J.E.; Morrell, J.J. Relationship between incipient decay, strength, and chemical composition of wood during incipient brown rot decay. Wood Fiber Sci. 1992, 25, 278–288. [Google Scholar]
- Schultze-Dewitz, G. Relations between elasticity and static and impact bending strength of pinewood after exposure to basidiomycetes. Holz Als Roh-Und Werkst. 1966, 24, 506–512. [Google Scholar] [CrossRef]
- Bariska, M.; Osuk, A.; Bosshard, H.H. Änderung der mechanischen Eigenschaften von Holz nach Abbau durch Basidiomyceten. Holz Als Roh-Und Werkst. 1983, 41, 241–245. [Google Scholar] [CrossRef]
- Venäläinen, M.; Partanen, H.; Harju, A. The strength loss of Scots pine timber in an accelerated soil contact test. Int. Biodeterior. Biodegrad. 2013, 86, 150–152. [Google Scholar] [CrossRef]
- Goodell, B.; Jellison, J.; Liu, G.; Daniel, A.; Paszcynski, F.; Fekete, S.; Krishnamurthy, L.; Jun, L.; Xu, G. Low molecular weight chelators and phenolic compounds isolated from wood decay fungi and their role in the fungal biodegradation of wood. J. Biotechnol. 1997, 53, 133–162. [Google Scholar] [CrossRef]
- Arantes, V.; Jellison, J.; Goodell, B. Peculiarities of brown-rot fungi and biochemical Fenton reaction with regard to their potential as a model for bioprocessing biomass. Appl. Microbiol. Biotechnol. 2012, 94, 323–338. [Google Scholar] [CrossRef] [PubMed]
- Baldrian, P.; Valaskova, V. Degradation of cellulose by basidiomycetous fungi. Fems Microbiol. Rev. 2008, 32, 501–521. [Google Scholar] [CrossRef] [PubMed]
- Goodell, B.; Zhu, Y.; Kim, S.; Kafle, K.; Eastwood, D.; Daniel, G.; Jellison, J.; Yoshida, M.; Groom, L.; Pingali, S.; et al. Biotechnology for Biofuels Modification of the nanostructure of lignocellulose cell walls via a non-enzymatic lignocellulose deconstruction system in brown rot wood—Decay fungi. Biotechnol. Biofuels 2017, 10, 179. [Google Scholar] [CrossRef] [PubMed]
- Dix, N.J. Fungal Ecology; Chapman & Hall: London, UK, 1995. [Google Scholar]
- Engelund, E.T.; Thygesen, L.G.; Svensson, S.; Hill, C.A.S. A critical discussion of the physics of wood–water interactions. Wood Sci. Technol. 2012, 47, 141–161. [Google Scholar] [CrossRef]
- Stone, J.E.; Scallan, A.M. A structural model for the cell wall of swollen wood pulp fibres based on accessibility to macromolecules. Cellul. Chem. Technol. 1968, 2, 343–358. [Google Scholar]
- Hoffmeyer, P.; Engelund, E.T.; Thygesen, L.G. Equilibrium moisture content (EMC) in Norway spruce during the first and second desorptions. Holzforschung 2011, 65, 875–882. [Google Scholar] [CrossRef]
- Simpson, W. Sorption theories applied to wood. Wood Fiber 1980, 12, 183–195. [Google Scholar]
- Christensen, G.N.; Kelsey, K.E. The rate of sorption of water vapor by wood. Holz Roh Werkst 1959, 17, 178–188. [Google Scholar] [CrossRef]
- Thybring, E.E.; Thygesen, L.G.; Burgert, I. Hydroxyl accessibility in wood cell walls as affected by drying and re-wetting procedures. Cellulose 2017, 24, 2375–2384. [Google Scholar] [CrossRef]
- Maréchal, Y.; Chanzy, H. The hydrogen bond network in Iβ cellulose as observed by infrared spectrometry. J. Mol. Struct. 2000, 523, 183–196. [Google Scholar] [CrossRef]
- Hofstetter, K.; Hinterstoisser, B.; Salmén, L. Moisture uptake in native cellulose—The roles of different hydrogen bonds: A dynamic FT-IR study using Deuterium exchange. Cellulose 2006, 13, 131–145. [Google Scholar] [CrossRef]
- Fredriksson, M.; Thybring, E.E. Scanning or desorption isotherms? Characterising sorption hysteresis of wood. Cellulose 2018, 25, 4477–4485. [Google Scholar] [CrossRef]
- Cardias Williams, F.; Hale, M.D. The resistance of wood chemically modified with isocyanates: The role of moisture content in decay suppression. Int. Biodeterior. Biodegrad. 2003, 52, 215–221. [Google Scholar] [CrossRef]
- Hill, C.A.S.; Ormondroyd, G.A. Dimensional changes in Corsican pine (Pinus nigra Arnold) modified with acetic anhydride measured using a helium pycnometer. Holzforschung 2004, 58, 544–547. [Google Scholar] [CrossRef]
- Thybring, E.E. The decay resistance of modified wood influenced by moisture exclusion and swelling reduction. Int. Biodeterior. Biodegrad. 2013, 82, 87–95. [Google Scholar] [CrossRef]
- Kirker, G.; Zelink, S.; Gleber, S.-G.; Vine, D.; Finney, L.; Chen, S.; Hong, Y.P.; Uyarte, O.; Vogt, S.; Jellison, J.; et al. Synchrotron-based X-ray fluorescence microscopy enables multiscale spatial visualization of ions involved in fungal lignocellulose deconstruction. Sci. Rep. 2017, 7, 41798. [Google Scholar] [CrossRef] [PubMed]
- Zelinka, S.L.; Gleber, S.-C.; Vogt, S.; Rodriguez Lopez, G.M.; Jakes, J. Threshold for ion movements in wood cell walls below fiber saturation observed by X-ray fluorescence microscopy (XFM). Holzforschung 2015, 69, 441–448. [Google Scholar] [CrossRef]
- Schmidt, O. Wood and Tree Fungi: Biology, Damage, Protection and Use; Springer: Berlin, Germany, 2006. [Google Scholar]
- Meyer, L.; Brischke, C. Fungal decay at different moisture levels of selected European-grown wood species. Int. Biodeterior. Biodegrad. 2015, 103, 23–29. [Google Scholar] [CrossRef]
- KIFS. Kemikalieinspektionens föreskrifter (KIFS 1998:8) om kemiska produkter och biotekniska organismer; KIFS: Sundbyberg, Sweden, 1998. [Google Scholar]
- Hingston, J.A.; Collins, C.D.; Murphy, R.J.; Lester, J.N. Leaching of chromated copper arsenate wood preservatives: A review. Environ. Pollut. 2000, 111, 53–66. [Google Scholar] [CrossRef]
- Townsend, T.; Dubey, B.; Tolaymat, T.; Solo-Gabriele, H. Preservative leaching from weathered CCA-treated wood. J. Environ. Manag. 2005, 75, 105–113. [Google Scholar] [CrossRef]
- Hill, C. Wood Modifications: Chemical, Thermal, and Other Processes; John Wiley and Sons, Ltd.: Hoboken, NJ, USA, 2006. [Google Scholar]
- Van der Lugt, P.; Vogtländer, J.G. Wood acetylation: A potential route towards climate change mitigation. Wit Trans. Built Environ. 2014, 142, 241–252. [Google Scholar]
- Militz, H. Die Verbesserung das Schwind-und Quellverhaltens und der Daurhaftigkeit von Holz mittels Behnadlung mit unkatalysiertem Essigsäureanhydrid. Holz Als Roh-Und Werkst. 1991, 49, 147–152. [Google Scholar] [CrossRef]
- Larsson Brelid, P.; Simonson, R.; Bergman, Ö.; Nilsson, T. Resistance of acetylated wood to biological degradation. Holz Als Roh-Und Werkst. 2000, 58, 331–337. [Google Scholar] [CrossRef]
- Schneider, M.H. New cell wall and cell lumen wood polymer composites. J. Wood Sci. Technol. 1995, 36, 429–433. [Google Scholar] [CrossRef]
- Lande, S.; Eikenes, M.; Westin, M.; Schneider, M. Furfurylation of wood: Chemistry; properties and commerzialisation. Dev. Commer. Wood Preserv. 2008, 982, 337–355. [Google Scholar]
- Bryne, L.E.; Wålinder, M.E.P. Ageing of modified wood. Part 1: Wetting properties of acetylated, furfurylated, and thermally modified wood. Holzforschung 2010, 64, 295–304. [Google Scholar] [CrossRef]
- Thygesen, L.G.; Barsberg, S.; Venås, T.M. The fluorescence characteristics of furfurylated wood studied by fluorescence spectroscopy and confocal laser scanning microscopy. Wood Sci. Technol. 2010, 44, 51–65. [Google Scholar] [CrossRef]
- Tjeerdsma, B.F.; Boonstra, M.; Pizzi, A.; Militz, H. Characterisation of thermally modified wood: Molecular reasons for wood performance improvement. Holz Als Roh-Und Werkst. 1998, 56, 149–153. [Google Scholar] [CrossRef]
- Welzbacher, C.R. Verhalten Von Nach Neuen Thermischen Modifikationsverfahren Behandelter Fichte und Kiefer Unter Besonderer Berücksichtigung der Dauerhaftigkeit Gegenüber Holzzerstörenden Mikroorganismen—Performance of Spruce and Pine Timber, Treated by Novel Thermal Modification. Ph.D. Thesis, University of Hamburg, Hamburg, Germany, 2007. [Google Scholar]
- Windeisen, E.; Bächle, H.; Zimmer, B.; Wegener, G. Relations between chemical changes and mechanical properties of thermally treated wood. Holzforschung 2009, 63, 773–778. [Google Scholar]
- Pfriem, A.; Zauer, M.; Wagenführ, A. Alteration of the unsteady sorption behaviour of maple (Acer pseudoplatanus L.) and spruce (Picea abies (L.) Karst.) due to thermal modification. Holzforschung 2010, 64, 235–241. [Google Scholar] [CrossRef]
- Ibach, R.; Rowell, R.M. Improvements in Decay Resistance Based on Moisture Exclusion. Mol. Cryst. Liq. Cryst. 2000, 353, 22–33. [Google Scholar] [CrossRef]
- Epmeier, H.; Westin, M.; Rapp, A. Differently modified wood: Comparison of some selected properties. Scand. J. For. Res. 2004, 19, 31–37. [Google Scholar] [CrossRef]
- Rowell, R.M.; Ibach, R.; Nilsson, T. Influence of moisture on brown-rot fungal attack on wood. In Proceedings of the 3rd meeting of the Nordic-Baltic Network in Wood Material Science and Engineering (WSE), Helsinki, Finland, 29–30 October 2007; pp. 65–69. [Google Scholar]
- Rapp, A.O.; Brischke, C.; Welzbacher, C.R.; Jazayeri, L. Increased resistance of thermally modiied norway spruce timber (TMT) against brown rot decay by Oligoporus placenta—A study on the mode of protective action. Wood Res. 2008, 53, 13–26. [Google Scholar]
- Nordiska träskyddsföreningen. Conditions for Approval of Industrially Protected Wood in the Nordic Countries; No 2; Nordiska Träskyddsföreningen: Stockholm, Sweden, 2017. [Google Scholar]
- Hill, C.; Popescu, C.; Rautkari, L.; Ormondroyd, G.; Xie, Y.; Jalaludin, Z. The role of hydroxyl groups in determining the sorption properties of modified wood. In Proceedings of the Seventh European Conference on Wood Modification, Lisbon, Portugal, 10–12 March 2014. [Google Scholar]
- Popescu, C.-M.; Hill, C.A.S.; Curling, S.; Ormondroyd, G.; Xie, Y. The water vapour sorption behaviour of acetylated birch wood: How acetylation affects the sorption isotherm and accessible hydroxyl content. J. Mater. Sci. 2013, 49, 2362–2371. [Google Scholar] [CrossRef]
- Rowell, R.M.; Ibach, R.E.; McSweeny, J.; Nilsson, T. Understanding decay resistance, dimensional stability and strength changes in heat-treated and acetylated wood. Wood Mater. Sci. Eng. 2009, 4, 14–22. [Google Scholar] [CrossRef]
- Popescu, C.-M.; Hill, C.A.S.; Popescu, M.-C. Water adsorption in acetylated birch wood evaluated through near infrared spectroscopy. Int. Wood Prod. J. 2016, 7, 61–65. [Google Scholar] [CrossRef]
- Passarini, L.; Zelinka, S.L.; Glass, S.V.; Hunt, C.G. Effect of weight percent gain and experimental method on fiber saturation point of acetylated wood determined by differential scanning calorimetry. Wood Sci. Technol. 2017, 51, 1291–1305. [Google Scholar] [CrossRef]
- Beck, G.; Thybring, E.E.; Thygesen, L.G.; Hill, C. Characterization of moisture in acetylated and propionylated radiata pine using low-field nuclear magnetic resonance (LFNMR) relaxometry. Holzforschung 2017, 72, 225–233. [Google Scholar] [CrossRef]
- Thygesen, L.G.; Tang Engelund, E.; Hoffmeyer, P. Water sorption in wood and modified wood at high values of relative humidity. Part I: Results for untreated, acetylated, and furfurylated Norway spruce. Holzforschung 2010, 64, 315–323. [Google Scholar] [CrossRef]
- Hill, C.A.S.; Forster, S.C.; Farahani, M.R.M.; Hale, M.D.C.; Ormondroyd, G.A.; Williams, G.R. An investigation of cell wall micropore blocking as a possible mechanism for the decay resistance of anhydride modified wood. Int. Biodeterior. Biodegrad. 2005, 55, 69–76. [Google Scholar] [CrossRef]
- Hill, C.A.S.; Curling, S.F.; Kwon, J.H.; Marty, V. Decay resistance of acetylated and hexanoylated hardwood and softwood species exposed to Coniophora puteana. Holzforschung 2009, 63, 619–625. [Google Scholar] [CrossRef]
- Papadopoulos, A.N.; Hill, C.A.S. The biological effectiveness of wood modified with linear chain carboxylic acid anhydrides against Coniophora Puteana. Holz Als Roh-Und Werkst. 2002, 60, 329–332. [Google Scholar] [CrossRef]
- Papadopoulos, A.N.; Hill, C.A.S. The sorption of water vapour by anhydride modified softwood. Wood Sci. Tech. 2003, 37, 221–231. [Google Scholar] [CrossRef]
- Beck, G.; Strohbusch, S.; Larnøy, E.; Militz, H.; Hill, C. Accessibility of hydroxyl groups in anhydride modified wood as measured by deuterium exchange and saponification. Holzforschung 2017, 72, 17–23. [Google Scholar] [CrossRef]
- Meyer, L.; Brischke, C.; Pilgård, A. Moisture performance based wood durability testing. In Proceedings of the IRG Annual meeting (ISSN 2000-8953), Kuala Lumpur, Malaysia, 6–10 May 2012. IRG/WP 12-20495. [Google Scholar]
- Venås, T.M. A Study of Mechanisms Related to the Fungal Decay Protection Rendered by Wood Furfurylation. Ph.D. Thesis, University of Copenhagen, Copenhagen, Denmark, 2008. [Google Scholar]
- Thygesen, L.G.; Elder, T. Moisture in Untreated, Acetylated, and Furfurylated Norway Spruce Monitored During Drying Below Fiber Saturation Using Time Domain NMR. Wood Fiber Sci. 2009, 41, 194–200. [Google Scholar]
- Dieste, A.; Krause, A.; Mai, C.; Sèbe, G.; Grelier, S.; Militz, H. Modification of Fagus sylvatica L. with 1,3-dimethylol-4,5-dihydroxy ethylene urea (DMDHEU). Part 2: Pore size distribution determined by differential scanning calorimetry. Holzforschung 2009, 63, 89–93. [Google Scholar] [CrossRef]
- Dieste, A.; Krause, A.; Mai, C.; Militz, H. The calculation of EMC for the analysis of wood/water relations in Fagus sylvatica L. modified with 1,3-dimethylol-4,5-dihydroxyethyleneurea. Wood Sci. Technol. 2009, 44, 597–606. [Google Scholar] [CrossRef][Green Version]
- Phuong, L.X.; Takayama, M.; Shida, S.; Matsumoto, Y.; Aoyagi, T. Determination of the accessible hydroxyl groups in heat-treated Styrax tonkinensis (Pierre) Craib ex Hartwich wood by hydrogen-deuterium exchange and 2H NMR spectroscopy. Holzforschung 2007, 61, 488–491. [Google Scholar] [CrossRef]
- Hakkou, M.; Pétrissans, M.; Gérardin, P.; Zoulalian, A. Investigations of the reasons for fungal durability of heat-treated beech wood. Polym. Degrad. Stab. 2006, 91, 393–397. [Google Scholar] [CrossRef]
- Altgen, M.; Willems, W.; Hosseinpourpia, R.; Rautkari, L. Hydroxyl accessibility and dimensional changes of Scots pine sapwood affected by alterations in the cell wall ultrastructure during heat-treatment. Polym. Degrad. Stab. 2018, 52, 244–252. [Google Scholar] [CrossRef]
- Hoffmeyer, P.; Jensen, S.K.; Jones, D.; Klinke, H.B.; Felby, C. Sorption properties of steam treated wood and plant fibres. In Proceedings of the First European Conference on Wood Modification, Ghent, Belgium, 3–4 April 2003; pp. 177–189. [Google Scholar]
- Scheiding, W.; Direske, M.; Zauer, M. Water absorption of untreated and thermally modified sapwood and heartwood of Pinus sylvestris L. Eur. J. Wood Wood Prod. 2016, 74, 585–589. [Google Scholar] [CrossRef]
- Zauer, M.; Pfriem, A.; Wagenführ, A. Toward improved understanding of the cell-wall density and porosity of wood determined by gas pycnometry. Wood Sci. Technol. 2013, 6, 1197–1211. [Google Scholar] [CrossRef]
- Biziks, V.; Andersons, B.; Sansonetti, E.; Andersone, I.; Militz, H.; Grinins, J. One-stage thermo-hydro treatment (THT) of hardwoods: An analysis of form stability after five soaking-drying cycles. Holzforschung 2014, 69, 563–571. [Google Scholar] [CrossRef]
- Ringman, R.; Pilgard, A.; Brischke, C.; Richter, K. Mode of action of brown rot decay resistance in modified wood: A review. Holzforschung 2014, 68, 239–246. [Google Scholar] [CrossRef]
- Boonstra, M.J.; Tjeerdsma, B. Chemical analysis of heat treated softwoods. Holz Als Roh-Und Werkst. 2006, 64, 204–211. [Google Scholar] [CrossRef]
- Boonstra, M.J.; Van Acker, J.; Kegel, E.; Stevens, M. Optimisation of a two-stage heat treatment process: durabilityaspects. Wood Sci. Technol. 2007, 41, 31–57. [Google Scholar] [CrossRef]
- Rowell, R.M. Chemical modification of wood. In Wood Chemistry and Wood Composites; Taylor & Francis: Boca Raton, FL, USA, 2005. [Google Scholar]
- Schmöllerl, B.; Alfredsen, G.; Fossdal, C.G.; Westin, M.; Steitz, A. Molecular investigation of Postia placenta growing in modified wood. In Proceedings of the International Research Group on Wood Protection, Queenstown, New Zealand, 8–12 May 2011. Document No. IRG/WP 11-10756. [Google Scholar]
- Alfredsen, G.; Pilgård, A.; Fossdal, C.G. Characterisation of Postia placenta colonisation during 36 weeks in acetylated southern yellow pine sapwood at three acetylation levels including genomic DNA and gene expression quantification of the fungus. Holzforschung 2016, 70, 1055–1065. [Google Scholar] [CrossRef]
- Ringman, R.; Pilgård, A.; Kölle, M.; Brischke, C.; Richter, K. Effects of thermal modification on Postia placenta wood degradation dynamics: Measurements of mass loss, structural integrity and gene expression. Wood Sci. Technol. 2016, 50, 385–397. [Google Scholar] [CrossRef]
- Ringman, R.; Pilgård, A.; Richter, K. Effect of wood modification on gene expression during incipient Postia placenta decay. Int. Biodeterior. Biodegrad. 2014, 86, 86–91. [Google Scholar] [CrossRef]
- Beck, G.; Hegnar, O.A.; Fossdal, C.G.; Alfredsen, G. Acetylation of Pinus radiata delays hydrolytic depolymerisation by the brown-rot fungus Rhondonia placenta. Int. Biodeterior. Biodegrad. 2018, 135, 39–52. [Google Scholar] [CrossRef]
- Alfredsen, G.; Fossdal, C.G.; Nagy, N.E.; Jellison, J.; Goodell, B. Furfurylated wood: Impact on Postia placenta gene expression and oxalate crystal formation. Holzforschung 2016, 70, 947–962. [Google Scholar] [CrossRef]
- Ringman, R.; Pilgård, A.; Richter, K. In vitro oxidative and enzymatic degradation of modified wood. Int. Wood Prod. J. 2015, 6, 36–39. [Google Scholar] [CrossRef]
- Verma, P.; Mai, C. Hydrolysis of cellulose and wood powder treated with DMDHEU by a hydrolase enzyme complex, Fenton’s reagent, and in a liquid culture of Trametes Versicolor. Holzforschung 2010, 64, 69–75. [Google Scholar] [CrossRef]
- Hosseinpourpia, R.; Mai, C. Mode of action of brown rot decay resistance of thermally modified wood: Resistance to Fenton’s reagent. Holzforschung 2016, 70, 691–697. [Google Scholar] [CrossRef]
- Xie, Y.; Xiao, Z.; Mai, C. Degradation of chemically modified Scots pine (Pinus sylvestris L.) with Fenton reagent. Holzforschung 2014, 69, 153–161. [Google Scholar] [CrossRef]
- Hegnar, O.A.; Goodell, B.; Felby, C.; Johansson, L.; Labbé, N.; Kim, K.; Eijsink, V.G.H.; Alfredsen, G.; Várnai, A. Challenges and opportunities in mimicking non-enzymatic brown-rot decay mechanisms for pretreatment of Norway spruce. Wood Sci. Technol. 2019, 53, 291–311. [Google Scholar] [CrossRef]
- Curling, S.F.; Clausen, C.A.; Winandy, J.E. Relationships between mechanical properties, weight loss and chemical composition of wood during incipient brown-rot decay. For. Prod. J. 2002, 52, 34–39. [Google Scholar]
- Ringman, R.; Pilgård, A.; Brischke, C.; Windeisen, E.; Richter, K. Incipient brown rot decay in modified wood: Patterns of mass loss, structural integrity, moisture and acetyl content in high resolution. Int. Wood Prod. J. 2017, 8, 172–182. [Google Scholar] [CrossRef]
- Thybring, E.E.; Kymäläinen, M.; Rautkari, L. Moisture in modified wood and its relevance for fungal decay. IForest 2018, 11, 418–422. [Google Scholar] [CrossRef]
- Zelinka, S.L.; Ringman, R.; Pilgård, A.; Thybring, E.E.; Jakes, J.E.; Richter, K. The role of chemical transport in the brown-rot decay resistance of modified wood. Int. Wood Prod. J. 2016, 7, 35–43. [Google Scholar] [CrossRef]
- Jellison, J.; Chandhoke, V.; Goodell, B.; Fekete, F. A The isolation and immunolocalization of iron-binding compounds produced by Gloeophyllum trabeum. Appl. Microbiol. Biotechnol. 1991, 35, 805–809. [Google Scholar] [CrossRef]
- Kim, Y.S.; Wi, A.G.; Lee, K.H.; Singh, S.P. Cytochemical Localization of Hydrogen Peroxide Production during Wood Decay by Brown-Rot Fungi Tyromyces palustris and Coniophora puteana. Holzforschung 2002, 56, 7–12. [Google Scholar] [CrossRef]
- Rowell, R.M. Chemical modification of wood: A short review. Wood Mater. Sci. Eng. 2006, 1, 29–33. [Google Scholar] [CrossRef]
- Hunt, C.G.; Zelinka, S.L.; Frihart, C.R.; Lorenz, L.; Yelle, D.; Gleber, S.-C.; Vogt, S.; Jakes, J.E. Acetylation increases relative humidity threshold for ion transport in wood cell walls—A means to understanding decay resistance. Int. Biodeterior. Biodegrad. 2018, 133, 230–237. [Google Scholar] [CrossRef]
- Hosseinpourpia, R.; Mai, C. Mode of action of brown rot decay resistance of acetylated wood: Resistance to Fenton’s reagent. Wood Sci. Technol. 2016, 50, 413–426. [Google Scholar] [CrossRef]
- Zelinka, S.L.; Glass, S.V.; Stone, D.S. A percolation model for electrical conduction in wood with implications for wood-water relations. Wood Fiber Sci. 2008, 40, 544–552. [Google Scholar]
- Jakes, J.E.; Plaza, N.; Stone, D.S.; Hunt, C.G.; Glass, S.V.; Zelinka, S.L. Mechanism of Transport Through Wood Cell Wall Polymers. J. For. Prod. Ind. 2013, 2, 10–13. [Google Scholar]
- Hearle, J.W.S. The electrical resistance of textile materials: IV. Theory. J. Text. Inst. Trans. 1953, 44, T177–T198. [Google Scholar] [CrossRef]
- Obataya, E.; Minato, K. Potassium acetate-catalyzed acetylation of wood: Extraordinarily rapid acetylation at 120 degrees C. Wood Sci. Technol. 2008, 42, 567–677. [Google Scholar] [CrossRef]
- Lande, S.; Westin, M.; Schneider, M. Properties of furfurylated wood. Scand. J. For. Res. 2004, 19, 22–30. [Google Scholar] [CrossRef]
- Yasuda, R.; Minato, K.; Norimoto, M. Chemical modification of wood by non-formaldehyde cross-linking reagents. Wood Sci. Technol. 1994, 28, 209–218. [Google Scholar] [CrossRef]
- Kwon, J.H.; Hill, C.A.S.; Ormondroyd, G.A.; Karim, S. Changes in the cell wall volume of a number of wood species due to reaction with acetic anhydride. Holzforschung 2007, 61, 138–142. [Google Scholar] [CrossRef]
- Farahani, M.R.M. Decay Resistance of Modified Wood. Ph.D. Thesis, University of Wales, Bangor, UK, 2003. [Google Scholar]
- Forster, S.C. The Decay Resistance of Chemically Modified Softwood. Ph.D. Thesis, University of Wales, Bangor, UK, 1999. [Google Scholar]
- Allan, G.G.; Ritzenthaler, P. The microporosity of pulp: The nature of the pore size distribution. Tappi J. 1991, 74, 205–212. [Google Scholar]
- Alince, B. Comments on porosity or swollen pulp fibers analyzed by solute-exclusion. Tappi J. 1991, 74, 200–202. [Google Scholar]
- Rowland, S.P.; Bertoniere, N.R. Some interactions of water-soluble solutes with cellulose and Sephadex. Text. Res. J. 1976, 46, 770–775. [Google Scholar] [CrossRef]
- Thybring, E.E.; Kymäläinen, M.; Rautkari, L. Experimental techniques for characterising water in wood covering the range from dry to fully water-saturated. Wood Sci. Technol. 2018, 52, 297–329. [Google Scholar] [CrossRef]
- Flournoy, D.S.; Kirk, K.; Highley, T.L. Wood Decay by Brown-Rot Fungi: Changes in Pore Structure and Cell Wall Volume. Holzforschung 1991, 45, 383–388. [Google Scholar] [CrossRef]
- Hosseinpourpia, R.; Mai, C. Mode of action of brown rot decay resistance in phenol-formaldehyde-modified wood: Resistance to Fenton’s reagent. Holzforschung 2015, 70, 253–259. [Google Scholar] [CrossRef]
- Zelinka, S.L.; Lambrecht, M.J.; Glass, S.V.; Wiedenhoeft, A.C.; Yelle, D.J. Examination of water phase transitions in Loblolly pine and cell wall components by differential scanning calorimetry. Thermochim. Acta 2012, 533, 39–45. [Google Scholar] [CrossRef]
- Thomson, W. On the Equilibrium of Vapour at a Curved Surface of Liquid. Proc. R. Soc. Edinb. 1872, 7, 63–68. [Google Scholar] [CrossRef]
- Griffin, D.M. Water Potential and Wood-Decay Fungi. Annu. Rev. Phytopathol. 1977, 15, 319–329. [Google Scholar] [CrossRef]
- Clarke, R.W.; Jennings, D.H.; Coggings, C.R. Growth of Serpula lacrimans in relation to water potential of substrate. Trans. Br. Mycol. Soc. 1980, 75, 271–280. [Google Scholar] [CrossRef]
- Viitanen, H. Modelling the Time Factor in the Development of Mould Fungi-the Effect of Critical Humidity and Temperature Conditions on Pine and Spruce Sapwood. Holzforschung 1997, 51, 6–14. [Google Scholar] [CrossRef]
- Schmidt, O. Indoor wood-decay basidiomycetes: Damage, causal fungi, physiology, identification and characterization, prevention and control. Mycol. Prog. 2007, 6, 261–279. [Google Scholar] [CrossRef]
- Mantanis, G.I.; Young, R.A. Wetting of wood. Wood Sci. Technol. 1997, 31, 339. [Google Scholar] [CrossRef]
- Zauer, M.; Hempel, S.; Pfriem, A.; Mechtcherine, V.; Wagenführ, A. Investigations of the pore-size distribution of wood in the dry and wet state by means of mercury intrusion porosimetry. Wood Sci. Technol. 2014, 48, 1229–1240. [Google Scholar] [CrossRef]
- Wang, W.; Yan, N. Characterizing Liquid Resin Penetration In Wood Using A Mercury Intrusion Porosimeter. Wood Fiber Sci. 2005, 37, 505–513. [Google Scholar]
- Zauer, M.; Kretzschmar, J.; Großmann, L.; Pfriem, A.; Wagenführ, A. Analysis of the pore-size distribution and fiber saturation point of native and thermally modified wood using differential scanning calorimetry. Wood Sci. Technol. 2013, 48, 177–193. [Google Scholar] [CrossRef]
- Bastani, A.; Adamopoulos, S.; Militz, H. Water uptake and wetting behaviour of furfurylated, N-methylol melamine modified and heat-treated wood. Eur. J. Wood Wood Prod. 2015, 73, 627–634. [Google Scholar] [CrossRef]
- Dieste, A.; Krause, A.; Militz, H. Modification of Fagus sylvatica (L.) with 1,3-dimethylol-4,5-dihydroxyethylene urea (DMDHEU): Part 1. Estimation of heat adsorption by the isosteric method (Hailwood-Horrobin model) and by solution calorimetry. Holzforschung 2008, 62, 577–583. [Google Scholar] [CrossRef]
- Hill, C.A.S.; Papadopoulos, A.N. A Review of Methods Used to Determine the Size of the Cell Wall Microvoids of Wood. J. Inst. Wood Sci. 2001, 15, 337–345. [Google Scholar]
- Plötze, M.; Niemz, P. Porosity and pore size distribution of different wood types as determined by mercury intrusion porosimetry. Eur. J. Wood Wood Prod. 2011, 69, 649–657. [Google Scholar] [CrossRef]
- Park, S.; Venditti, R.A.; Jameel, H.; Pawlak, J.J. Changes in pore size distribution during the drying of cellulose fibers as measured by differential scanning calorimetry. Carbohydr. Polym. 2006, 66, 97–103. [Google Scholar] [CrossRef]
- Gao, X.; Zhuang, S.; Jin, J.; Cao, P. Bound Water Content and Pore Size Distribution in Swollen Cell Walls Determined by NMR Technology. BioResources 2015, 10, 8208–8224. [Google Scholar] [CrossRef]
- Weatherwax, R.C. Collapse of cell-wall pores during drying of cellulose. J. Colloid Interface Sci. 1977, 62, 432–446. [Google Scholar] [CrossRef]
- Weatherwax, R.C.; Caulfield, D.F. Cellulose aerogels: An improved method for preparing a highly expanded form of dry cellulose. Tappi J. 1971, 54, 985–986. [Google Scholar]
- Kang, K.-Y.; Hwang, K.-R.; Park, J.-Y.; Lee, J.-P.; Kim, J.-S.; Lee, J.-S. Critical Point Drying: An Effective Drying Method for Direct Measurement of the Surface Area of a Pretreated Cellulosic Biomass. Polymers 2018, 10, 676. [Google Scholar] [CrossRef]
- Cox, J.; McDonald, P.J.; Gardiner, B.A. A study of water exchange in wood by means of 2D NMR relaxation correlation and exchange. Holzforschung 2010, 64, 259–266. [Google Scholar] [CrossRef]
- Martínez-Sanz, M.; Gidley, M.J.; Gilbert, E.P. Application of X-ray and neutron small angle scattering techniques to study the hierarchical structure of plant cell walls: A review. Carbohydr. Polym. 2015, 125, 120–134. [Google Scholar]
- Xu, P.; Donaldson, L.A.; Gergely, Z.R.; Staehelin, L.A. Dual-axis electron tomography: A new approach for investigating the spatial organization of wood cellulose microfibrils. Wood Sci. Technol. 2006, 41, 101–116. [Google Scholar] [CrossRef]
- Torstensen, J.Ø.; Liu, M.; Jin, S.-A.; Deng, L.; Hawaru, A.I.; Syverud, K.; Spontak, R.J.; Gregersen, Ø.W. Swelling and free-volume characteristics of TEMPO-oxidized cellulose nanofibril films. Biomacromolecules 2018, 19, 1016–1025. [Google Scholar] [CrossRef] [PubMed]
- Kulasinski, K.; Guyer, R.A. Quantification of nanopore networks: Application to amorphous polymers. J. Phys. Chem. C 2016, 120, 28144–28151. [Google Scholar] [CrossRef]
- Zauer, M.; Meissner, F.; Plagge, R.; Wagenführ, A. Capillary pore-size distribution and equilibrium moisture content of wood determined by means of pressure plate technique. Holzforschung 2015, 70, 137–143. [Google Scholar] [CrossRef]
- Fredriksson, M.; Johansson, S.A. Method for Determination of Absorption Isotherms at High Relative Humidity Levels: Measurements on Lime-Silica Brick and Norway Spruce. Dry. Technol. 2016, 34, 132–141. [Google Scholar] [CrossRef]
- Katz, J.R. Die Quellung. In Ergebnisse der Exakten Naturwissenschaften; Springer: Berlin, Germany, 1924; Volume 3, p. 316. [Google Scholar]
- Cousins, W.J. Young’s modulus of hemicellulose as related to moisture content. Wood Sci. Technol. 1978, 12, 161–167. [Google Scholar] [CrossRef]
- Kelley, S.S.; Rials, T.G.; Glasser, W.G.; Al, E. Relaxation behaviour of the amorphous components of wood. J. Mater. Sci. 1987, 22, 617–624. [Google Scholar] [CrossRef]
- Olsson, A.M.; Salmén, L. The softening behavior of hemicelluloses related to moisture. ACS Symp. Ser. 2004, 864, 184–197. [Google Scholar]
- Paul, W.; Ohlmeyer, M.; Leithoff, H.; Boonstra, M.J.; Pizzi, A. Optimising the properties of OSB by a one-step heat pre-treatment process. Holz Als Roh-Und Werkst. 2006, 64, 227–234. [Google Scholar] [CrossRef]
- Åkerholm, M.; Salmen, L. Softening of wood polymers induced by moisture studied by dynamic FTIR spectroscopy. J. Appl. Polym. Sci. 2004, 94, 2032–2040. [Google Scholar] [CrossRef]
- Gröndahl, M.; Teleman, A.; Gatenholm, P. Effect of acetylation on the material properties of glucuronoxylan from aspen wood. Carbohydr. Polym. 2003, 52, 359–366. [Google Scholar] [CrossRef]
- Willems, W. Glassy state of wood polymers in native and thermally modified wood: Effects on long-term material performance in service. Int. Wood Prod. J. 2016, 7, 71–75. [Google Scholar] [CrossRef]
- Hill, C.A.S.; Ramsay, J.; Keating, B.; Laine, K.; RAutkari, L.; Hughes, M.; Constant, B. The water vapour sorption properties of thermally modified and densified wood. J. Mater. Sci. 2012, 47, 3191–3197. [Google Scholar] [CrossRef]
- Salmén, L.; Possler, H.; Stevanic, J.S.; Stanzl-Tschegg, S.E. Analysis of thermally treated wood samples using dynamic FT-IR-spectroscopy. Holzforschung 2008, 62, 676–678. [Google Scholar] [CrossRef]
- Krause, A.; Jones, D.; van der Zee, M.; Militz, H. Interlacement treatment—wood modification with n-methylol compounds. In Proceedings of the First European Conference on Wood Modification, Ghent, Belgium, 3–4 April 2003; pp. 317–327. [Google Scholar]
- Nordstierna, L.; Lande, S.; Westin, M.; Karlsson, O.; Furó, I. Towards novel wood-based materials: Chemical bonds between lignin-like model molecules and poly(furfuryl alcohol) studied by NMR. Holzforschung 2008, 62, 709–713. [Google Scholar] [CrossRef]
- Ehmcke, G.; Pilgård, A.; Koch, G.; Richter, K. Topochemical analyses of furfuryl alcohol-modified radiata pine (Pinus radiata) by UMSP, light microscopy and SEM. Holzforschung 2017, 71, 821–831. [Google Scholar] [CrossRef]
- Herold, N.; Grigsby, W.J.; Franich, R.A.; Pfriem, A. Investigations of wood veneer during furfuryl alcohol modification using DMTA. Eur. J. Wood Wood Prod. 2015, 73, 693–695. [Google Scholar] [CrossRef]
- Ganser, C.; Hirn, U.; Rohm, S.; Schennach, R. AFM nanoindentation of pulp fibers and thin cellulose films at varying relative humidity. Holzforschung 2013, 68, 53–60. [Google Scholar] [CrossRef]
- Beck, G.; Thybring, E.E.; Thygesen, L.G. Brown-rot fungal degradation and de-acetylation of acetylated wood. Int. Biodeterior. Biodegrad. 2018, 135, 62–70. [Google Scholar] [CrossRef]
- Verma, P.; Junga, U.; Militz, H.; Mai, C. Protection mechanisms of DMDHEU treated wood against white and brown rot fungi. Holzforschung 2009, 63, 371–378. [Google Scholar] [CrossRef]
- Thybring, E.E. Water relations in untreated and modified wood under brown-rot and white-rot decay. Int. Biodeterior. Biodegrad. 2017, 118, 134–142. [Google Scholar] [CrossRef]
- Eastwood, D.C.; Floudas, D.; Binder, M.; Majcherczyk, A.; Schneider, P.; Aerts, A.; Asiegbu, F.O.; Baker, S.E.; Barry, K.; Bendiksby, M.; et al. The plant cell wall-decomposing machinery underlies the functional diversity of forest fungi. Science 2011, 333, 762–765. [Google Scholar] [CrossRef] [PubMed]
- EN 113. Wood Preservatives—Test Method for Determining the Protective Effectiveness Against Wood Destroying Basidiomycetes: Determination of Toxic Values; European Committee for Standardization (CEN): Brussels, Belgium, 1996; p. 31. [Google Scholar]
- AWPA E10. American Wood-Preservers’ Association Standard E10-91. Standard Method for Testing Wood Preservatives by Laboratory Soil-Block Cultures; AWPA: Birmingham, AL, USA, 1991. [Google Scholar]
- Zhang, J.; Presley, G.N.; Hammel, K.E.; Ryu, J.-S.; Menke, J.R.; Figueroa, M.; Hu, D.; Orr, G.; Schilling, J.S. Localizing gene regulation reveals a staggered wood decay mechanism for the brown rot fungus Postia placenta. Proc. Natl. Acad. Sci. USA 2016, 113, 10968–10973. [Google Scholar] [CrossRef] [PubMed]
- Alfredsen, G.; Ringman, R.; Pilgård, A.; Fossdal, C.G. New insight regarding mode of action of brown rot decay of modified wood based on DNA and gene expression studies: A review. Int. Wood Prod. J. 2015, 6, 2008–2013. [Google Scholar] [CrossRef]
- Esteves, B.M.; Pereira, H.M. Wood modification by heat treatment: A review. BioResources 2009, 4, 370–404. [Google Scholar]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ringman, R.; Beck, G.; Pilgård, A. The Importance of Moisture for Brown Rot Degradation of Modified Wood: A Critical Discussion. Forests 2019, 10, 522. https://doi.org/10.3390/f10060522
Ringman R, Beck G, Pilgård A. The Importance of Moisture for Brown Rot Degradation of Modified Wood: A Critical Discussion. Forests. 2019; 10(6):522. https://doi.org/10.3390/f10060522
Chicago/Turabian StyleRingman, Rebecka, Greeley Beck, and Annica Pilgård. 2019. "The Importance of Moisture for Brown Rot Degradation of Modified Wood: A Critical Discussion" Forests 10, no. 6: 522. https://doi.org/10.3390/f10060522
APA StyleRingman, R., Beck, G., & Pilgård, A. (2019). The Importance of Moisture for Brown Rot Degradation of Modified Wood: A Critical Discussion. Forests, 10(6), 522. https://doi.org/10.3390/f10060522