Susceptibility of Small-Diameter Norway Spruce Understory Stumps to Heterobasidion Spore Infection

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
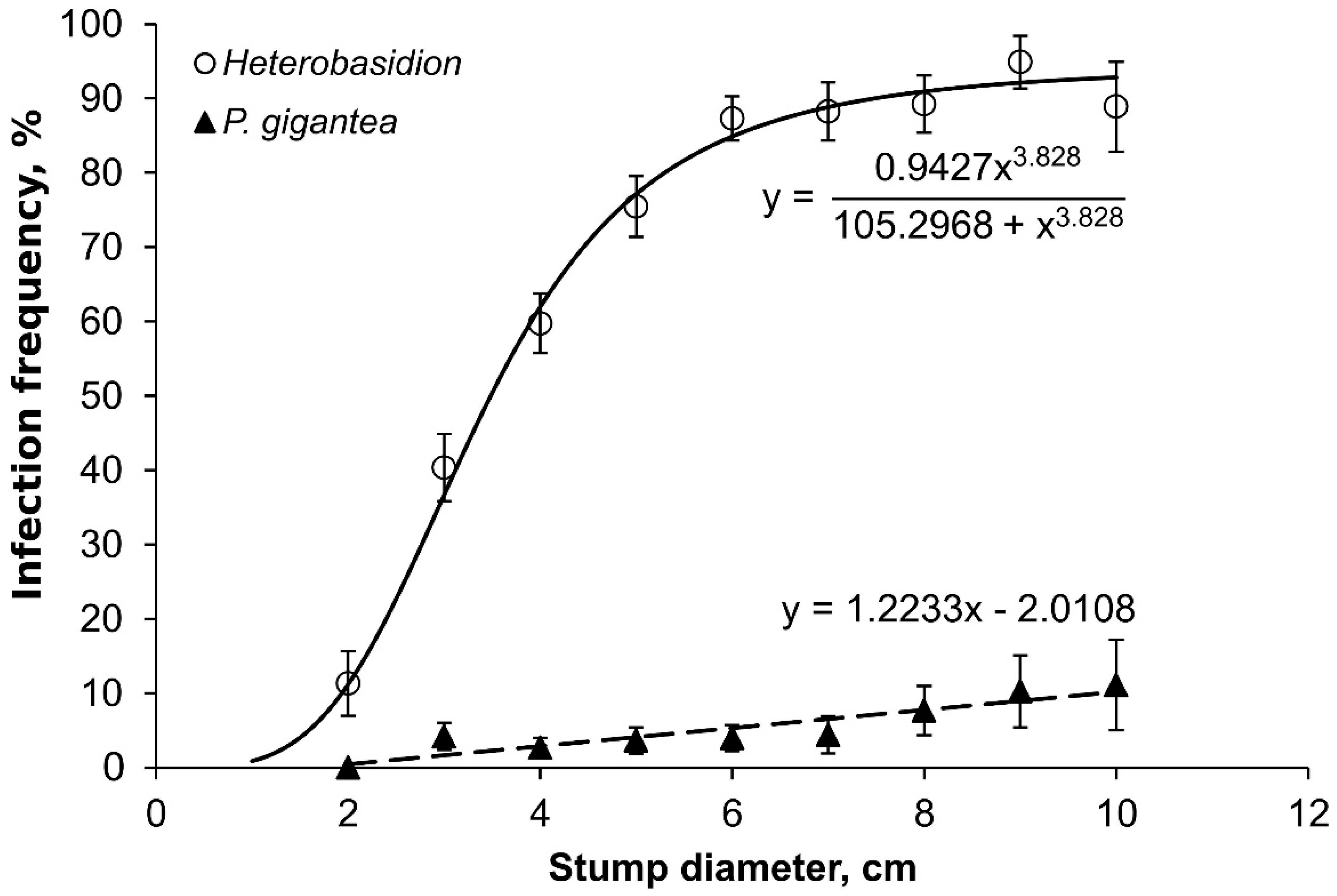
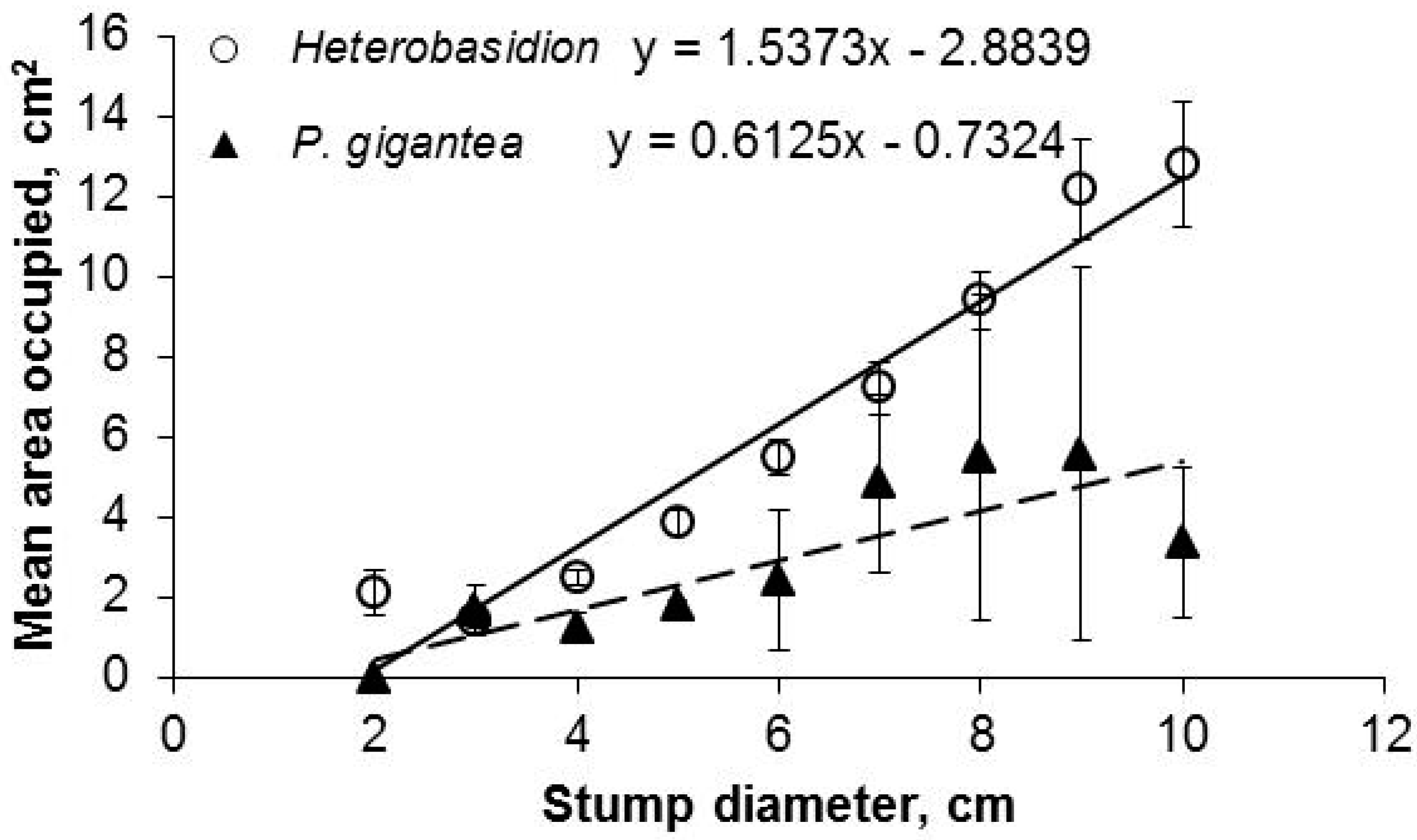
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Redfern, D.B.; Stenlid, J. Spore dispersal and infection. In Heterobasidion annosum: Biology, Ecology, Impact and Control; Woodward, S., Stenlid, J., Karjalainen, R., Hüttermann, A., Eds.; CAB International: Wallingford, UK, 1998; pp. 105–141. [Google Scholar]
- Korhonen, K.; Holdenrieder, O. Neue erkenntnisse über den wurzelschwamm (heterobasidion annosum s.l.). eine literaturübersicht. Forst. Und. Holz. 2005, 60, 206–211. [Google Scholar]
- Asiegbu, F.O.; Adomas, A.; Stenlid, J. Conifer root and butt rot caused by Heterobasidion Annosum (Fr.) Bref. s.l. Mol. Plant Pathol. 2005, 6, 395–409. [Google Scholar] [CrossRef] [PubMed]
- Stenlid, J.; Wästerlund, I. Estimating the frequency of stem rot in Picea abies using an increment borer. Scand. J. Forest. Res. 1986, 1, 303–308. [Google Scholar] [CrossRef]
- Vasiliauskas, R.; Stenlid, J. Spread of S and P group isolates of Heterobasidion annosum within and among Picea abies trees in central Lithuania. Can. J. For. Res. 1998, 28, 961–966. [Google Scholar] [CrossRef]
- Tamminen, P. Butt rot in Norway spruce in Southern Finland. Commun. Inst. For. Fenn. 1985, 127, 1–52. [Google Scholar]
- Bendz-Hellgren, M.; Stenlid, J. Long-term reduction in the diameter growth of butt rot affected Norway spruce, Picea abies. For. Ecol. Manag. 1995, 74, 239–243. [Google Scholar] [CrossRef]
- Bendz-Hellgren, M.; Stenlid, J. Decreased volume growth of Picea abies in response to Heterobasidion annosum infection. Can. J. For. Res. 1997, 27, 1519–1524. [Google Scholar] [CrossRef]
- Bendz-Hellgren, M.; Lipponen, K.; Solheim, H.; Thomsen, I.M. The nordic countries. In Heterobasidion annosum: Biology, Ecology, Impact and Control; Woodward, S., Stenlid, J., Karjalainen, R., Hüttermann, A., Eds.; CAB International: Wallingford, UK, 1998; pp. 333–345. [Google Scholar]
- Oliva, J.; Samils, N.; Johansson, U.; Bendz-Hellgren, M.; Stenlid, J. Urea treatment reduced Heterobasidion annosum s.l. root rot in Picea abies after 15 years. For. Ecol. Manag. 2008, 7, 2876–2882. [Google Scholar] [CrossRef]
- Klavina, D.; Menkis, A.; Gaitnieks, T.; Velmala, S.; Lazdins, A.; Rajala, T.; Pennanen, T. Analysis of Norway spruce dieback phenomenon in Latvia—A belowground perspective. Scand. J. Forest. Res. 2016, 31, 156–165. [Google Scholar] [CrossRef]
- Arhipova, N.; Gaitnieks, T.; Donis, J.; Stenlid, J.; Vasaitis, R. Butt rot incidence, causal fungi and related yield loss in Picea abies stands of Latvia. Can. J. For. Res. 2011, 41, 2337–2345. [Google Scholar] [CrossRef]
- Gaitnieks, T.; Šica, L.; Indriksons, A.; Grantina, L. Root-rot detection in Norway spruce stands using pulsed electronic tool. In Proceedings of the 11th International Conference Root and Butt Rots of Forest Trees, Poznan – Bialowieza, Poland, 16–22 August 2004; pp. 379–385. [Google Scholar]
- Korhonen, K.; Bobko, I.; Hanso, S.; Piri, T.; Vasilauskas, A. Intersterility groups of Heterobasidion annosum in some spruce and pine stands in Byelorussia, Lithuania and Estonia. Eur. J. Plant Pathol. 1992, 22, 384–392. [Google Scholar] [CrossRef]
- Korhonen, K.; Piri, T. The main hosts and distribution of the S and P groups of Heterobasidion annosum in Finland. In Proceedings of the 8th IUFRO Conference on Root and Butt Rots, Wik, Sweden and Haikko, Finland, 9–16 August 1993; Johansson, M., Stenlid, J., Eds.; Swedish University of Agricultural Sciences: Uppsala, Sweden, 1994; pp. 260–268. [Google Scholar]
- Isomäki, A.; Kallio, T. Consequences of injury caused by timber harvesting machines on the growth and decay of spruce (Picea abies (L.) Karst.). Acta For. Fenn. 1974, 136, 1–25. [Google Scholar] [CrossRef]
- Stenlid, J.; Redfern, D.B. Spread within the tree and stand. In Heterobasidion annosum: Biology, Ecology, Impact and Control; Woodward, S., Stenlid, J., Karjalainen, R., Hüttermann, A., Eds.; CAB International: Wallingford, UK, 1998; pp. 125–141. [Google Scholar]
- Korhonen, K. Simulated stump treatment experiments for monitoring the efficacy of Phlebiopsis gigantea against Heterobasidion. Root and butt rots of forest trees. In Proceedings of the 10th International Conference on Root and Butt Rots, Québec, QC, Canada, 16–22 September 2003; Laflamme, G., Bérubé, J.A., Bussiéres, G., Eds.; Canadian Forest Service: Chalk River, ON, Canada, 2003; pp. 206–210. [Google Scholar]
- Thor, M. Optional stump treatment against Heterobasidion annosum in European forestry—Current situation. In Proceedings of the 10th International Conference on Root and Butt Rots, Québec, QC, Canada, 16–22 September 2003; Laflamme, G., Bérubé, J.A., Bussiéres, G., Eds.; Canadian Forest Service: Chalk River, ON, Canada, 2003; pp. 170–175. [Google Scholar]
- Semenkova, I.G. Attack of spruce understory by Heterobasidion annosum. Nauchn. Tr. Mosk. Lesotek. Inst. 1971, 38, 150–166. [Google Scholar]
- Schönhar, S. Untersuchungen über die infection der fichte (Picea abies Karst.) durch Heterobasidion Annosum (Fr.) Bref. Allgemeine Forst-Und Jagdzeitung 1995, 166, 14–17. [Google Scholar]
- Piri, T. Early development of root rot in young Norway spruce planted on sites infected by Heterobasidion in Southern Finland. Can. J. For. Res. 2003, 33, 604–611. [Google Scholar] [CrossRef]
- Gruduls, K.; Gaitnieks, T.; Donis, J.; Liepa, I. Heterobasidion spp. in Picea abies understory: Incidence and impact on radial growth of trees. Res. For. 2012, 2, 21–24. [Google Scholar]
- Piri, T.; Korhonen, K. Infection of advance regeneration of Norway spruce by Heterobasidion parviporum. Can. J. For. Res. 2001, 31, 937–942. [Google Scholar] [CrossRef]
- Piri, T.; Valkonen, S. Incidence and spread of Heterobasidion root rot in uneven-aged Norway spruce stands. Can. J. For. Res. 2013, 43, 872–877. [Google Scholar] [CrossRef]
- Paludan, F. Infection and spread of Fomes annosus in young Norway spruce. Det Forstl. Forsøgsvæs. Dan. 1966, 30, 21–47. [Google Scholar]
- Solheim, H. Infeksjon av rotkjuke på granstubber til ulike årstider og effekten av ureabehandling. Nor. Inst. Skogforsk. Rapp. Skogforsk 1994, 3, 1–10. [Google Scholar]
- Berglund, M.; Carlsson, T.; Rönnberg, J. Infection of Heterobasidion spp. in late pre-commercial thinnings of Picea abies in southern Sweden. In Proceedings of the 12th International Conference on Root and Butt Rots, Berkeley, California, Medford, OR, USA, 12–19 August 2007; Garbelotto, M., Gonthier, P., Eds.; The University of California: Berkeley, CA, USA, 2008; pp. 221–225. [Google Scholar]
- Gunulf, A.; Mc Carthy, R.; Rönnberg, J. Control efficacy of stump treatment and influence of stump height on natural spore infection by Heterobasidion spp. of precommercial thinning stumps of Norway spruce and birch. Silva Fenn. 2012, 46, 655–665. [Google Scholar] [CrossRef]
- Gaitnieks, T.; Brauners, I.; Kenigsvalde, K.; Zaļuma, A.; Brūna, L.; Jansons, J.; Burņeviča, N.; Lazdiņš, A.; Vasaitis, R. Infection of pre-commercially cut stumps of Picea abies and Pinus sylvestris by Heterobasidion spp.—A comparative study. Silva Fenn. 2018, 52. [Google Scholar] [CrossRef]
- Gunulf, A.; Wang, L.; Englund, J.E.; Rönnberg, J. Secondary spread of Heterobasidion parviporum from small Norway spruce stumps to adjacent trees. Forest Ecol. Manag. 2013, 287, 1–8. [Google Scholar] [CrossRef]
- Sellin, A. Sapwood amount in Picea abies (L.) Karst. determined by tree age and radial growth rate. Holzforschung 1996, 50, 291–296. [Google Scholar] [CrossRef]
- Rishbeth, J. Dispersal of Fomes annosus Fr. and Peniophora gigantea (Fr.) Massee. Trans. Br. Mycol. Soc. 1959, 42, 243–260. [Google Scholar] [CrossRef]
- Yde-Andersen, A. Seasonal incidence of stump infection in Norway spruce by air-borne Fomes annosus spores. For. Sci. 1962, 8, 98–103. [Google Scholar]
- Stenlid, J. Homokaryotic Heterobasidion annosum mycelia in stumps of Norway spruce. In Proceedings of the 8th IUFRO Conference on Root and Butt Rots, Wik, Sweden and Haikko, Finland, 9–16 August 1993; Johansson, M., Stenlid, J., Eds.; Swedish University of Agricultural Sciences: Uppsala, Sweden, 1994; pp. 243–248. [Google Scholar]
- Brauners, I.; Brūna, L.; Gaitnieks, T. Testing the ‘Rotstop’ biological preparation for controlling Heterobasidion root rot in Latvia. Res. Rural Dev. 2014, 2, 97–102. [Google Scholar]
- Bendz-Hellgren, M.; Stenlid, J. Effects of clear-cutting, thinning, and wood moisture content on the susceptibility of Norway spruce stumps to Heterobasidion annosum. Can. J. For. Res. 1998, 28, 759–765. [Google Scholar] [CrossRef]
- Swedjemark, G.; Stenlid, J. Population dynamics of the root rot fungus Heterobasidion annosum following thinning of Picea abies. Oikos 1993, 66, 247–254. [Google Scholar] [CrossRef]
- Redfern, D.B. The effect of wood moisture on infection of Sitka spruce stumps by basidiospores of Heterobasidion annosum. Eur. J. Forest Pathol. 1993, 23, 218–235. [Google Scholar] [CrossRef]
- Vollbrecht, G.; Gemmel, P.; Pettersson, N. The effect of precommercial thinning on the incidence of Heterobasidion annosum in planted Picea abies. Scand. J. Forest. Res. 1995, 10, 37–41. [Google Scholar] [CrossRef]
- Woods, C.M.; Woodward, S.; Redfern, D.B. Receptivity of Picea sitchensis stumps to infection by Heterobasidion annosum basidiospores. Forestry 2000, 73, 457–465. [Google Scholar] [CrossRef]
- Morrison, D.J. Factors affecting infection of precommercial thinning stumps by Heterobasidion annosum in coastal British Columbia. In Proceedings of the Symposium on Research and Management of Annosus Root Disease (Heterobasidion annosum) in Western North America, Monterey, CA, USA, 18–21 April 1989; Otrosina, W.J., Scharpf, R.F., Eds.; Forest Service, Berkeley, Canada: Toronto, ON, Canada, 1989; pp. 95–100. [Google Scholar]
- Gibbs, J.N.; Greig, B.J.W.; Pratt, J.E. Fomes root rot in Thetford Forest, East Anglia: Past, present and future. Forestry 2002, 75, 191–202. [Google Scholar] [CrossRef]
- Piri, T. The spreading of the S type of Heterobasidion annosum from Norway spruce stumps to the subsequent tree stand. Eur. J. Forest Pathol. 1996, 26, 193–204. [Google Scholar] [CrossRef]
- Schönhar, S. Fungal red-rot in Norway spruce trees damaged during log extraction. Allg. Forst-Und Jagt-Ztg. 1975, 146, 72–75. [Google Scholar]
- Berglund, M.; Rönnberg, J. Effectiveness of treatment of Norway spruce stumps with Phlebiopsis gigantea at different rates of coverage for the control of Heterobasidion. For. Pathol. 2004, 34, 233–243. [Google Scholar] [CrossRef]
- Morrison, D.J.; Johnson, A.L.S. Stump colonization and spread of Fomes annosus 5 years after thinning. Can. J. For. Res. 1978, 8, 177–180. [Google Scholar] [CrossRef]
- Redfern, D.B.; Pratt, J.E.; Gregory, S.C.; MacAskill, G. Natural infection of Sitka spruce thinning stumps in Britain by spores of Heterobasidion annosum and long-term survival of the fungus. Forestry 2001, 74, 53–72. [Google Scholar] [CrossRef]
- Oliva, J.; Bernat, M.; Stenlid, J. Heartwood stump colonisation by Heterobasidion parviporum and H. annosum s.s. in Norway spruce (Picea abies) stands. For. Ecol. Manag. 2013, 295, 1–10. [Google Scholar] [CrossRef]
- Meredith, D.S. Further observations on fungi inhabiting pine stumps. Ann. Bot. 1960, 24, 63–78. [Google Scholar] [CrossRef]
- Berglund, M.; Rönnberg, J.; Holmer, L.; Stenlid, J. Comparison of five strains of Phlebiopsis gigantea and two Trichoderma formulations for treatment against natural Heterobasidion spore infections on Norway spruce stumps. Scand. J. Forest. Res. 2005, 20, 12–17. [Google Scholar] [CrossRef]
- Kenigsvalde, K.; Brauners, I.; Korhonen, K.; Zaļuma, A.; Mihailova, A.; Gaitnieks, T. Evaluation of the biological control agent Rotstop in controlling the infection of spruce and pine stumps by Heterobasidion in Latvia. Scand. J. Forest. Res. 2016, 31, 254–261. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Sample Plot No. | Age of Trees, Years | Age of Stumps at Sampling, Weeks | Number of Analyzed Stumps | Mean Stump Diameter (Range in Brackets), cm | Proportion of Infected Stumps (Heterobasidion/P. gigantea), % |
|---|---|---|---|---|---|
| 1 | 35 | 48 | 112 | 4.7 (2.3–9.7) | 57.1/12.5 |
| 2 | 36 | 39 | 162 | 4.1 (1.6–9.9) | 61.7/3.1 |
| 3 | 67 | 37 | 41 | 4.3 (1.8–9.7) | 56.1/0.0 |
| 4 | 67 | 33 | 89 | 5.5 (2.2–9.7) | 86.5/13.5 |
| 5 | 65 | 34 | 101 | 6.2 (1.7–10.4) | 76.2/1.0 |
| 6 | 64 | 34 | 128 | 6.3 (1.9–10.4) | 77.3/0.7 |
| 7 | 68 | 34 | 58 | 5.5 (1.6–9.0) | 65.5/0.0 |
| 8 | 48 | 33 | 65 | 5.3 (1.7–9.9) | 56.9/0.0 |
| Mean stand age, years. ± SD | 56.3 ± 14.3 |
| Mean stump age at sampling, weeks ± SD | 36.5 ± 5.1 |
| Total number of stumps | 756 |
| Mean stump diameter ± SD, cm. | 5.2 ± 2.1 |
| Number of infected stumps (Heterobasidion spp./P. gigantea) | 515/33 |
| Infection frequency Heterobasidion spp. /P.gigantea,% | 68/4 |
| Stump area occupied by Heterobasidion, cm2. Mean ± SD | 5.8 ± 5.5 |
| Mean stump area occupied by P. gigantea, cm2 ± SD | 3.3 ± 5.0 |
| Parameter | Estimate | SEM | z | P |
|---|---|---|---|---|
| Intercept | −0.77 | 1.40 | −0.55 | −0.58 |
| Stump diameter, cm | 0.65 | 0.06 | 10.55 | <0.01 |
| Age of stumps at sampling, weeks | −0.03 | 0.03 | −1.31 | 0.19 |
| Age of trees, years | −0.01 | 0.01 | −0.54 | 0.59 |
| Occurrence of P. gigantea | 0.86 | 0.56 | 1.54 | 0.12 |
| Parameter | Estimate | SEM | t | P |
|---|---|---|---|---|
| Intercept | 0.39 | 0.17 | 2.30 | 0.02 |
| Age of stumps at sampling, weeks | −0.01 | 0.01 | −2.54 | 0.01 |
| Stump diameter, cm | 0.10 | 0.01 | 16.02 | <0.01 |
| Age of trees, years | −0.01 | 0.01 | −0.24 | 0.81 |
| Parameter | Estimate | SEM | t | P |
|---|---|---|---|---|
| Intercept | −4.6 | 2.37 | −1.94 | 0.06 |
| Age of stumps at sampling, weeks | 0.06 | 0.04 | 1.68 | 0.10 |
| Stump diameter, cm | 0.01 | 0.06 | 0.03 | 0.98 |
| Age of trees, years | 0.04 | 0.02 | 2.73 | 0.01 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaitnieks, T.; Zaļuma, A.; Kenigsvalde, K.; Kļaviņa, D.; Brauners, I.; Piri, T. Susceptibility of Small-Diameter Norway Spruce Understory Stumps to Heterobasidion Spore Infection. Forests 2019, 10, 521. https://doi.org/10.3390/f10060521
Gaitnieks T, Zaļuma A, Kenigsvalde K, Kļaviņa D, Brauners I, Piri T. Susceptibility of Small-Diameter Norway Spruce Understory Stumps to Heterobasidion Spore Infection. Forests. 2019; 10(6):521. https://doi.org/10.3390/f10060521
Chicago/Turabian StyleGaitnieks, Tālis, Astra Zaļuma, Kristīne Kenigsvalde, Dārta Kļaviņa, Indulis Brauners, and Tuula Piri. 2019. "Susceptibility of Small-Diameter Norway Spruce Understory Stumps to Heterobasidion Spore Infection" Forests 10, no. 6: 521. https://doi.org/10.3390/f10060521
APA StyleGaitnieks, T., Zaļuma, A., Kenigsvalde, K., Kļaviņa, D., Brauners, I., & Piri, T. (2019). Susceptibility of Small-Diameter Norway Spruce Understory Stumps to Heterobasidion Spore Infection. Forests, 10(6), 521. https://doi.org/10.3390/f10060521