Non-Destructive Assessment of Wood Stiffness in Scots Pine (Pinus sylvestris L.) and its Use in Forest Tree Improvement


Abstract
1. Introduction
2. Materials and Methods
2.1. Test Material
2.2. Wood Density Assessment
2.3. SilviScan Data
2.4. Wood Stiffness Assessment through Acoustic Velocity
- Constant density of 1000 kg∙m−3 (DENCONST);
- Wood density estimated by SilviScan technology (DENSILV);
- Resistograph unadjusted density (DENRES);
- Resistograph adjusted (detrended and debarked) density (DENRES.TB);
- Density based on depth of Pilodyn’s pin penetration with bark (DENPIL);
- Density based on depth of Pilodyn’s pin penetration without bark (DENPIL.B).
2.5. Growth Data
2.6. Statistical Analysis
3. Results
3.1. Variation in Different Wood Stiffness (MOE) Estimates
3.2. Heritability
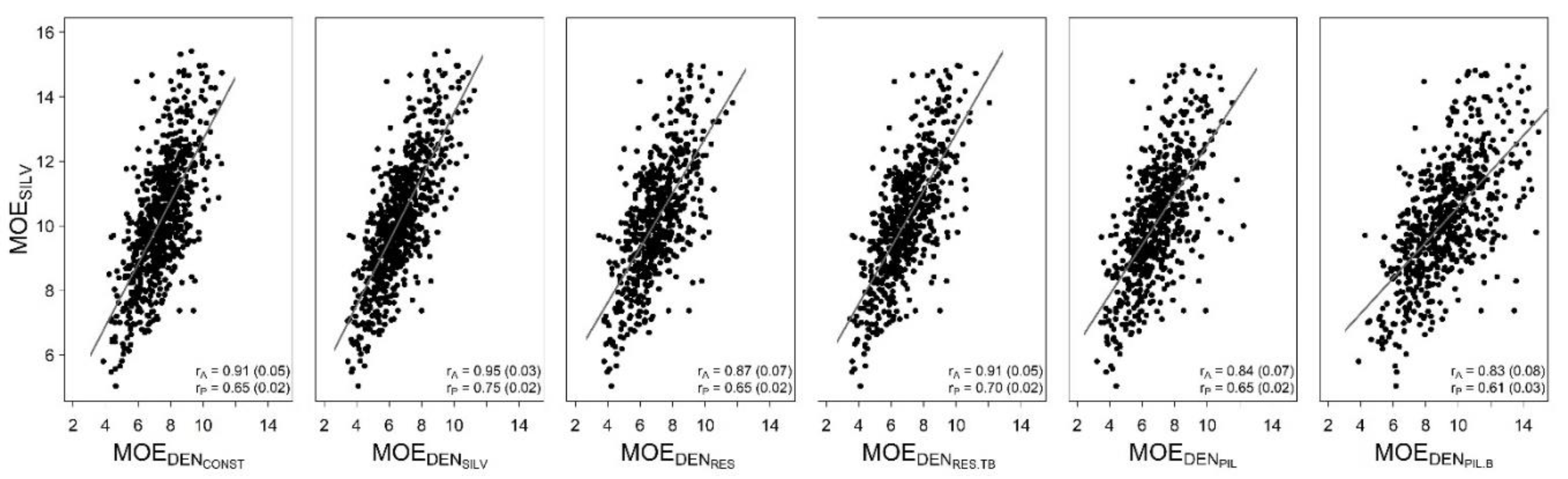
3.3. Comparison of Moduli of Elasticity
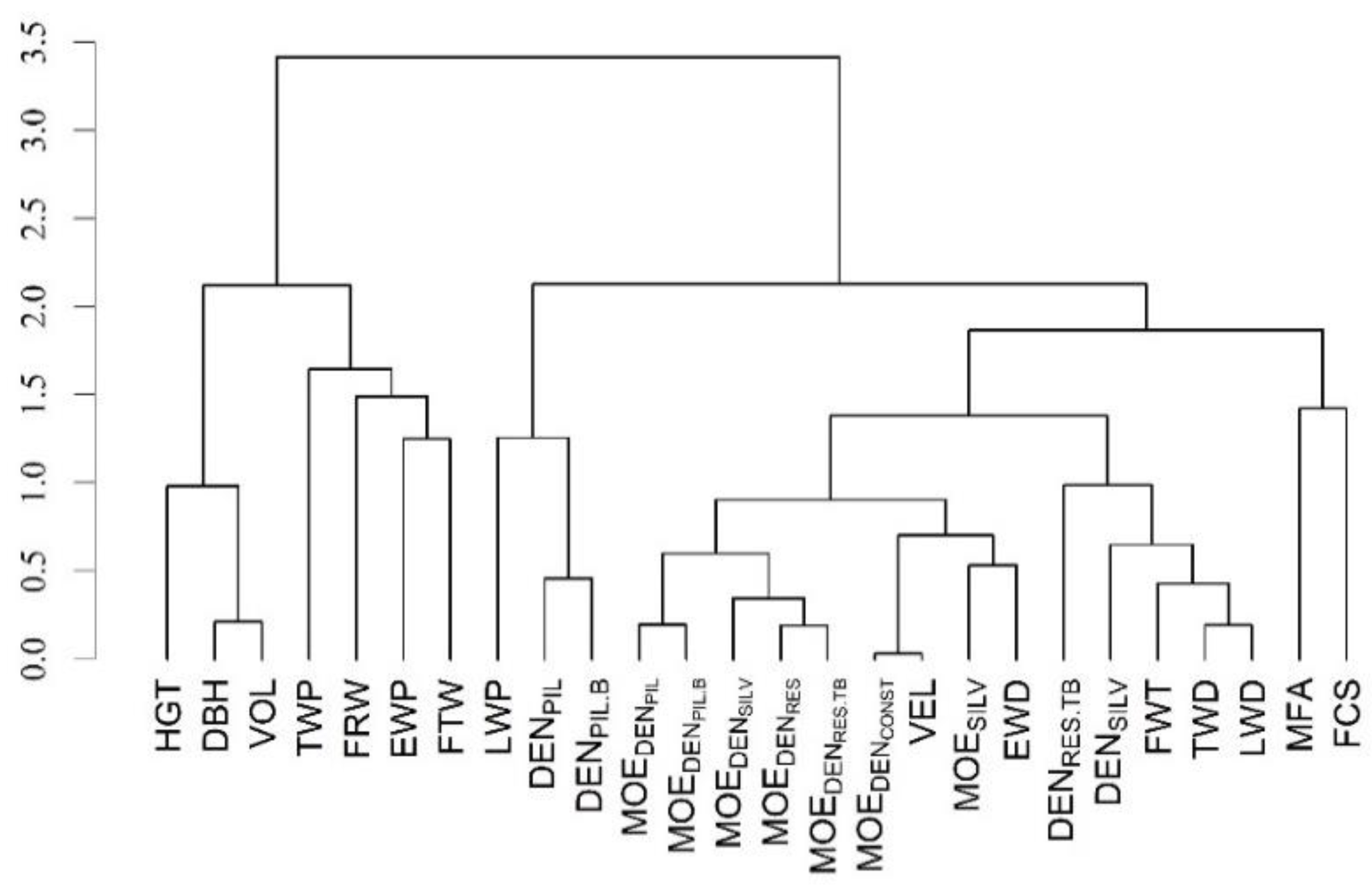
3.4. Phenotypic and Additive Genetic Correlations of Selected Moduli of Elasticity with Other Wood, Fiber, and Growth Traits
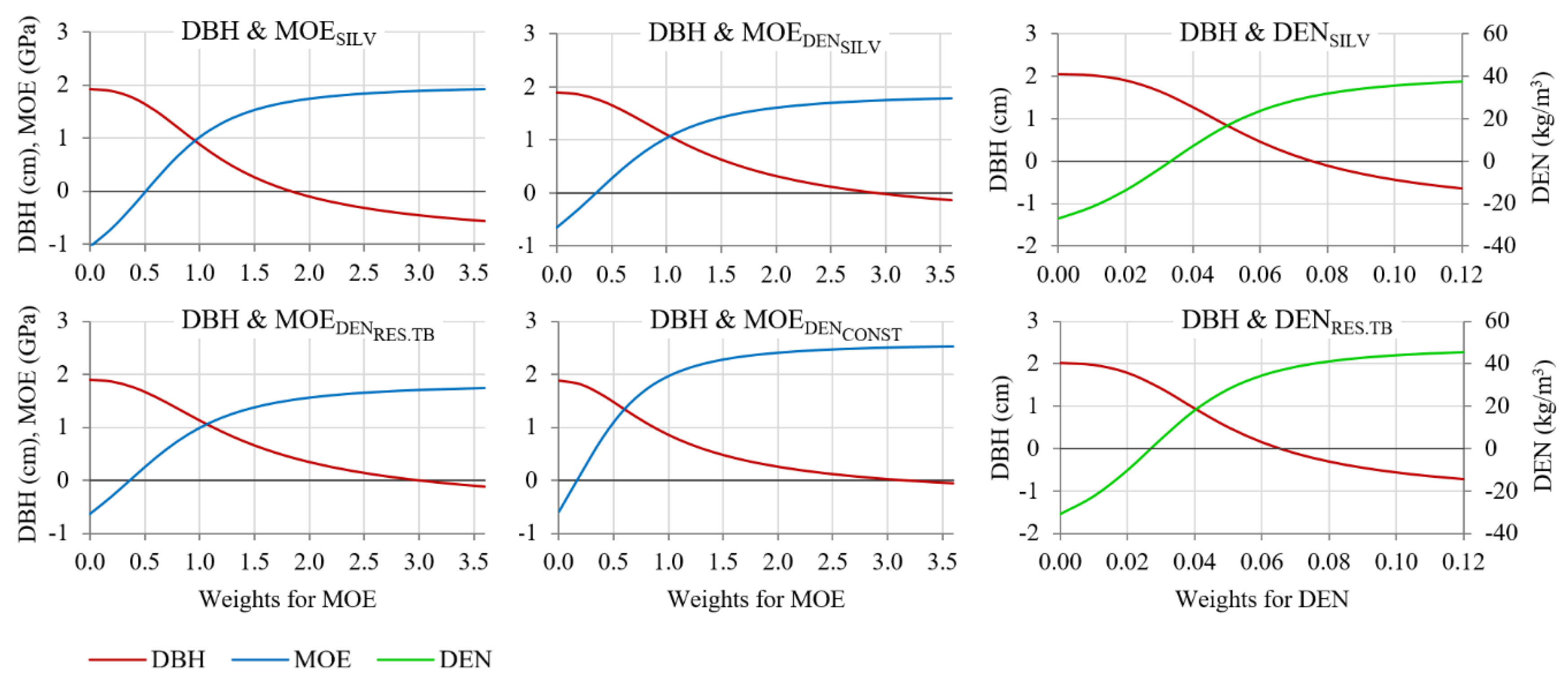
3.5. Index Selection
3.6. Response to Selection
4. Discussion
4.1. Estimation of Wood Stiffness
4.2. Heritability
4.3. Correlations
4.4. Factors Affecting Acoustic Velocity Measurements
4.5. Response to Selection
4.6. Practical Implications
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Falk, R.H. Wood as a sustainable building material. In Wood handbook - wood as an engineering material. General technical report FPL-GTR-190; U.S. Department of Agriculture, Forest Service, Forest Products Laboratory: Madison, WI, USA, 2010; Volume 190. [Google Scholar]
- Ramage, M.H.; Burridge, H.; Busse-Wicher, M.; Fereday, G.; Reynolds, T.; Shah, D.U.; Wu, G.; Yu, L.; Fleming, P.; Densley-Tingley, D. The wood from the trees: The use of timber in construction. Renew. Sust. Energ. Rev. 2017, 68, 333–359. [Google Scholar] [CrossRef]
- Wegner, T.; Skog, K.E.; Ince, P.J.; Michler, C.J. Uses and desirable properties of wood in the 21st century. J. For. 2010, 108, 165–173. [Google Scholar]
- Hong, Z.; Fries, A.; Wu, H. High negative genetic correlations between growth traits and wood properties suggest incorporating multiple traits selection including economic weights for the future Scots pine breeding programs. Ann. For. Sci. 2014, 71, 463–472. [Google Scholar] [CrossRef]
- Chen, Z.-Q.; Gil, M.R.G.; Karlsson, B.; Lundqvist, S.-O.; Olsson, L.; Wu, H.X. Inheritance of growth and solid wood quality traits in a large Norway spruce population tested at two locations in southern Sweden. Tree Genet. Genome. 2014, 10, 1291–1303. [Google Scholar] [CrossRef]
- Hayatgheibi, H.; Fries, A.; Kroon, J.; Wu, H.X. Genetic analysis of lodgepole pine (Pinus contorta) solid-wood quality traits. Can. J. For. Res. 2017, 47, 1303–1313. [Google Scholar] [CrossRef]
- Kennedy, S.G.; Cameron, A.D.; Lee, S.J. Genetic relationships between wood quality traits and diameter growth of juvenile core wood in Sitka spruce. Can. J. For. Res. 2013, 43, 1–6. [Google Scholar] [CrossRef]
- Wessels, C.B.; Malan, F.S.; Rypstra, T. A review of measurement methods used on standing trees for the prediction of some mechanical properties of timber. Eur. J. For. Res. 2011, 130, 881–893. [Google Scholar] [CrossRef]
- Legg, M.; Bradley, S. Measurement of stiffness of standing trees and felled logs using acoustics: A review. J. Acoust. Soc. Am. 2016, 139, 588–604. [Google Scholar] [CrossRef] [PubMed]
- Bucur, V. Acoustics of Wood, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2006; p. 394. [Google Scholar]
- Evans, R. Wood stiffness by X-ray diffractometry. In Characterization of the Cellulosic Cell Wall; Stokke, D., Groom, L., Eds.; Blackwell Publishing: Oxford, UK, 2006; pp. 138–146. [Google Scholar]
- Chen, Z.-Q.; Karlsson, B.; Lundqvist, S.-O.; Gil, M.R.G.; Olsson, L.; Wu, H.X. Estimating solid wood properties using Pilodyn and acoustic velocity on standing trees of Norway spruce. Ann. For. Sci. 2015, 72, 499–508. [Google Scholar] [CrossRef]
- El-Kassaby, Y.A.; Mansfield, S.; Isik, F.; Stoehr, M. In situ wood quality assessment in Douglas-fir. Tree Genet. Genome. 2011, 7, 553–561. [Google Scholar] [CrossRef]
- Newton, P.F. Acoustic-based non-destructive estimation of wood quality attributes within standing red pine trees. Forests 2017, 8, 380. [Google Scholar] [CrossRef]
- Raymond, C.A.; Joe, B.; Evans, R.; Dickson, R.L. Relationship between timber grade, static and dynamic modulus of elasticity, and SilviSvan properties for Pinus radiata in New South Wales. N. Z. J. For. Sci. 2007, 37, 186. [Google Scholar]
- Cherry, M.L.; Vikram, V.; Briggs, D.; Cress, D.W.; Howe, G.T. Genetic variation in direct and indirect measures of wood stiffness in coastal Douglas-fir. Can. J. For. Res. 2008, 38, 2476–2486. [Google Scholar] [CrossRef]
- Vikram, V.; Cherry, M.L.; Briggs, D.; Cress, D.W.; Evans, R.; Howe, G.T. Stiffness of Douglas-fir lumber: Effects of wood properties and genetics. Can. J. For. Res. 2011, 41, 1160–1173. [Google Scholar] [CrossRef]
- Wang, X.P.; Ross, R.J.; McClellan, M.; Barbour, R.J.; Erickson, J.R.; Forsman, J.W.; McGinnis, G.D. Nondestructive evaluation of standing trees with a stress wave method. Wood Fiber Sci. 2001, 33, 522–533. [Google Scholar]
- Auty, D.; Achim, A. The relationship between standing tree acoustic assessment and timber quality in Scots pine and the practical implications for assessing timber quality from naturally regenerated stands. Forestry 2008, 81, 475–487. [Google Scholar] [CrossRef]
- Wielinga, B.; Raymond, C.A.; James, R.; Matheson, A.C. Effect of green density values on Pinus radiata stiffness estimation using a stress-wave technique. N. Z. J. For. Sci. 2009, 39, 71–79. [Google Scholar]
- Kumar, S.; Jayawickrama, K.J.S.; Lee, J.; Lausberg, M. Direct and indirect measures of stiffness and strength show high heritability in a wind-pollinated radiata pine progeny test in New Zealand. Silvae Genet. 2002, 51, 256–261. [Google Scholar]
- Chauhan, S.S.; Walker, J.C.F. Variations in acoustic velocity and density with age, and their interrelationships in radiata pine. For. Ecol. Manag. 2006, 229, 388–394. [Google Scholar] [CrossRef]
- Hong, Z.; Fries, A.; Lundqvist, S.-O.; Gull, B.A.; Wu, H.X. Measuring stiffness using acoustic tool for Scots pine breeding selection. Scand. J. For. Res. 2015, 30, 363–372. [Google Scholar] [CrossRef]
- Fundova, I.; Funda, T.; Wu, H.X. Non-destructive wood density assessment of Scots pine (Pinus sylvestris L.) using Resistograph and Pilodyn. PLoS ONE 2018, 13. [Google Scholar] [CrossRef] [PubMed]
- Isik, F.; Li, B.L. Rapid assessment of wood density of live trees using the Resistograph for selection in tree improvement programs. Can. J. For. Res. 2003, 33, 2426–2435. [Google Scholar] [CrossRef]
- Walker, T.D.; Isik, F.; McKeand, S.E. Genetic variation in acoustic time of flight and drill resistance of juvenile wood in a large loblolly pine breeding population. For. Sci. 2019. [Google Scholar] [CrossRef]
- Hazel, L.N.; Lush, J.L. The efficiency of three methods of selection. J. Hered. 1942, 33, 393–399. [Google Scholar] [CrossRef]
- White, T.L.; Adams, W.T.; Neale, D.B. Forest Genetics; CAB International: Wallingford, UK, 2007. [Google Scholar]
- Namkoong, G. Optimum allocation of selection intensity in two stages of tuncation selection. Biometrics 1970, 26, 465–476. [Google Scholar] [CrossRef]
- Hazel, L.N. The genetic basis for constructing selection indexes. Genetics 1943, 28, 476–490. [Google Scholar] [PubMed]
- Falconer, D.S.; Mackay, T.F.C. Introduction to Quantitative Genetics; Longman: Harlow, UK, 1996; Volume 4, p. 464. [Google Scholar]
- Cotterill, P.P.; Dean, J.S. Successful Tree Breeding with Index Selection; Division of Forestry and Forest Products, CSIRO: Victoria, Australia, 1990.
- Ivković, M.; Wu, H.X.; McRae, T.A.; Powell, M.B. Developing breeding objectives for radiata pine structural wood production. I. Bioeconomic model and economic weights. Can. J. For. Res. 2006, 36, 2920–2931. [Google Scholar] [CrossRef]
- Fries, A. Genetic parameters, genetic gain and correlated responses in growth, fibre dimensions and wood density in a Scots pine breeding population. Ann. For. Sci. 2012, 69, 783–794. [Google Scholar] [CrossRef]
- Hannrup, B.; Ekberg, I.; Persson, A. Genetic correlations among wood, growth capacity and stem traits in Pinus sylvestris. Scand. J. For. Res. 2000, 15, 161–170. [Google Scholar] [CrossRef]
- RCoreTeam. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Brandel, G. Volume Functions for Individual Trees: Scots pine (Pinus sylvestris), Norway Spruce (Picea abies) and birch (Betula pendula & Betula pubescens); Report 26; Department of Forest Yield Research, Swedish University of Agricultural Sciences: Garpenberg, Sweden, 1990; p. 183. [Google Scholar]
- Gilmour, A.R.; Gogel, B.J.; Cullis, B.R.; Thompson, R. ASReml User Guide; Release 3.0; VSN International Ltd.: Hemel Hempstead, UK, 2009; Available online: www.vsni.co.uk (accessed on 11 February 2019).
- Smith, H.F. A discriminant function for plant selection. Ann. Eugen. 1936, 7, 240–250. [Google Scholar] [CrossRef]
- Desponts, M.; Perron, M.; DeBlois, J. Rapid assessment of wood traits for large-scale breeding selection in Picea mariana [Mill.] B.S.P. Ann. For. Sci. 2017, 74, 12. [Google Scholar] [CrossRef]
- Zhang, S.; Jiang, J.; Luan, Q. Index selection for growth and construction wood properties in Pinus elliottii open-pollinated families in southern China. South. For. J. For. Sci. 2018, 80, 209–216. [Google Scholar] [CrossRef]
- Lenz, P.; Auty, D.; Achim, A.; Beaulieu, J.; Mackay, J. Genetic improvement of white spruce mechanical wood traits—early screening by means of acoustic velocity. Forests 2013, 4, 575–594. [Google Scholar] [CrossRef]
- Baltunis, B.S.; Wu, H.X.; Powell, M.B. Inheritance of density, microfibril angle, and modulus of elasticity in juvenile wood of Pinus radiata at two locations in Australia. Can. J. For. Res. 2007, 37, 2164–2174. [Google Scholar] [CrossRef]
- Wang, X.P.; Carter, P.; Ross, R.J.; Brashaw, B.K. Acoustic assessment of wood quality of raw forest materials—A path to increased profitability. For. Prod. J. 2007, 57, 6–14. [Google Scholar]
- Carter, P. Breeding for wood quality; acoustic tools and technology. Presented at AFG & IUFRO SPWG Joint Conference, Hobart, Tasmania, April 2007. [Google Scholar]
- Lindström, H.; Harris, P.; Sorensson, C.T.; Evans, R. Stiffness and wood variation of 3-year old Pinus radiata clones. Wood Sci. Technol. 2004, 38, 579–597. [Google Scholar] [CrossRef]
- Grabianowski, M.; Manley, B.; Walker, J.C.F. Acoustic measurements on standing trees, logs and green lumber. Wood Sci. Technol. 2006, 40, 205–216. [Google Scholar] [CrossRef]
- Gao, S.; Wang, X.; Wang, L.; Allison, R.B. Effect of temperature on acoustic evaluation of standing trees and logs. Part 1. Laboratory investigation. Wood Fiber Sci. 2012, 44, 1–12. [Google Scholar]
- Chan, J.M.; Walker, J.C.; Raymond, C. Effects of moisture content and temperature on acoustic velocity and dynamic MOE of radiata pine sapwood boards. Wood Sci. Technol. 2011, 45, 609–626. [Google Scholar] [CrossRef]
- Wang, S.Y.; Chuang, S.T. Experimental data correction of the dynamic elastic moduli, velocity and density of solid wood as a function of moisture content above the fiber saturation point. Holzforschung 2000, 54, 309–314. [Google Scholar] [CrossRef]
- Newton, P. Quantifying the effects of wood moisture and temperature variation on time-of-flight acoustic velocity measures within standing red pine and jack pine trees. Forests 2018, 9, 527. [Google Scholar] [CrossRef]
- Škorpík, P.; Konrad, H.; Geburek, T.; Schuh, M.; Vasold, D.; Eberhardt, M.; Schueler, S. Solid wood properties assessed by non-destructive measurements of standing European larch (Larix decidua Mill.): Environmental effects on variation within and among trees and forest stands. Forests 2018, 9, 276. [Google Scholar] [CrossRef]
- Paradis, N.; Auty, D.; Carter, P.; Achim, A. Using a standing-tree acoustic tool to identify forest stands for the production of mechanically-graded lumber. Sensors 2013, 13, 3394. [Google Scholar] [CrossRef] [PubMed]
- Lindström, H.; Harris, P.; Nakada, R. Methods for measuring stiffness of young trees. Holz Als Roh-und Werkst. 2002, 60, 165–174. [Google Scholar] [CrossRef]
- Briggs, D.G.; Thienel, G.; Turnblom, E.C.; Lowell, E.; Dykstra, D.; Ross, R.J.; Wang, X.; Carter, P. Influence of thinning on acoustic velocity of Douglas-fir trees in western Washington and western Oregon. In Proceedings of the 15th International Symposium on Nondestructive Testing of Wood, Duluth, MN, USA, 10–12 September 2007; Forest Products Society: Madison, WI, USA, 2008; pp. 113–123. [Google Scholar]
- Ivković, M.; Wu, H.X.; Spencer, D.J.; McRae, T.A. Modelling the effects of stem sweep, branch size and wood stiffness of radiata pine on structural timber production. Austral. For. 2007, 70, 173–184. [Google Scholar] [CrossRef]
- Apiolaza, L.A.; Raymond, C.A.; Yeo, B.J. Genetic variation of physical and chemical wood properties of Eucalyptus globulus. Silvae Genet. 2005, 54, 160–166. [Google Scholar] [CrossRef]
- Chen, Z.-Q.; Karlsson, B.; Mörling, T.; Olsson, L.; Mellerowicz, E.J.; Wu, H.X.; Lundqvist, S.-O.; Gil, M.R.G. Genetic analysis of fiber dimensions and their correlation with stem diameter and solid-wood properties in Norway spruce. Tree Genet. Genome. 2016, 12, 123. [Google Scholar] [CrossRef]
- Hayatgheibi, H.; Fries, A.; Kroon, J.; Wu, H. Genetic analysis of fiber-dimension traits and combined selection for simultaneous improvement of growth and stiffness in lodgepole pine (Pinus contorta). Can. J. For. Res. 2019, 49, 500–509. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Trait | Units | Description | Min | Max | Mean | SD | CV | DMRT | |
|---|---|---|---|---|---|---|---|---|---|
| MOESILV | GPa | Benchmark modulus of elasticity from SilviScan | 5.0 | 16.0 | 10.2 | 1.9 | 18.5 | 0.39 (0.09) | b |
| VEL | km·s−1 | Acoustic velocity measured by Hitman | 2.8 | 4.7 | 3.8 | 0.3 | 9.0 | 0.37 (0.09) | - |
| MOECONST | GPa | Dynamic modulus of elasticity with DEN = 1000 | 7.7 | 22.3 | 14.9 | 2.6 | 17.7 | 0.37 (0.09) | a |
| MOEDENSILV | GPa | Dynamic modulus of elasticity with DEN = DENSILV | 3.4 | 11.2 | 6.7 | 1.5 | 21.7 | 0.46 (0.10) | d |
| MOEDENRES | GPa | Dynamic modulus of elasticity with DEN = DENRES/4 | 3.4 | 11.7 | 7.0 | 1.4 | 20.7 | 0.44 (0.11) | d |
| MOEDENRES.TB | GPa | Dynamic modulus of elasticity with DEN = DENRES.TB/4 | 3.4 | 12.0 | 6.9 | 1.5 | 21.7 | 0.45 (0.11) | d |
| MOEDENPIL | GPa | Dynamic modulus of elasticity with DEN = 1/DENPIL | 3.2 | 12.2 | 6.9 | 1.6 | 22.7 | 0.40 (0.10) | d |
| MOEDENPIL.B | GPa | Dynamic modulus of elasticity with DEN = 1/DENPIL.B | 3.9 | 16.7 | 9.2 | 2.1 | 22.8 | 0.41 (0.10) | c |
| Correlations with MOESILV | ||
|---|---|---|
| Genetic | Phenotypic | |
| VEL | 0.91 (0.05) | 0.65 (0.02) |
| MOEDENCONST | 0.91 (0.05) | 0.65 (0.02) |
| MOEDENSILV | 0.95 (0.03) | 0.75 (0.02) |
| MOEDENRES | 0.87 (0.07) | 0.65 (0.02) |
| MOEDENRES.TB | 0.91 (0.05) | 0.70 (0.02) |
| MOEDENPIL | 0.84 (0.07) | 0.65 (0.02) |
| MOEDENPIL.B | 0.83 (0.08) | 0.61 (0.03) |
| Genetic Correlations | Phenotypic Correlations | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| MOESILV | VEL | MOEDENCONST | MOEDENSILV | MOEDENRES.TB | MOESILV | VEL | MOEDENCONST | MOEDENSILV | MOEDENRES.TB | |
| Wood traits | ||||||||||
| DENSILV | 0.84 (0.07) | 0.73 (0.11) | 0.73 (0.11) | 0.87 (0.06) | 0.92 (0.05) | 0.60 (0.03) | 0.33 (0.04) | 0.33 (0.04) | 0.62 (0.03) | 0.53 (0.03) |
| DENRES.TB | 0.63 (0.13) | 0.58 (0.14) | 0.57 (0.15) | 0.76 (0.09) | 0.83 (0.07) | 0.45 (0.03) | 0.27 (0.04) | 0.27 (0.04) | 0.49 (0.03) | 0.64 (0.03) |
| DENPIL | 0.44 (0.18) | 0.50 (0.17) | 0.51 (0.17) | 0.57 (0.15) | 0.66 (0.14) | 0.33 (0.04) | 0.24 (0.04) | 0.24 (0.04) | 0.33 (0.04) | 0.35 (0.04) |
| DENPIL.B | 0.43 (0.18) | 0.48 (0.18) | 0.49 (0.17) | 0.62 (0.14) | 0.62 (0.14) | 0.26 (0.04) | 0.22 (0.04) | 0.22 (0.04) | 0.34 (0.04) | 0.36 (0.04) |
| EWD | 0.95 (0.05) | 0.81 (0.09) | 0.81 (0.09) | 0.88 (0.06) | 0.96 (0.05) | 0.51 (0.03) | 0.30 (0.04) | 0.31 (0.04) | 0.56 (0.03) | 0.47 (0.03) |
| TWD | 0.79 (0.08) | 0.69 (0.11) | 0.69 (0.11) | 0.83 (0.07) | 0.85 (0.07) | 0.66 (0.02) | 0.39 (0.03) | 0.39 (0.03) | 0.63 (0.02) | 0.58 (0.03) |
| LWD | 0.78 (0.08) | 0.70 (0.11) | 0.70 (0.11) | 0.83 (0.07) | 0.84 (0.07) | 0.73 (0.02) | 0.46 (0.03) | 0.46 (0.03) | 0.65 (0.02) | 0.61 (0.03) |
| EWP | 0.00 (0.23) | 0.00 (0.24) | 0.00 (0.24) | −0.06 (0.23) | −0.12 (0.24) | 0.20 (0.04) | 0.15 (0.04) | 0.14 (0.04) | 0.04 (0.04) | 0.08 (0.04) |
| TWP | −0.27 (0.23) | −0.21 (0.25) | −0.21 (0.25) | −0.23 (0.24) | −0.16 (0.25) | −0.36 (0.03) | −0.22 (0.04) | −0.21 (0.04) | -0.18 (0.04) | −0.19 (0.04) |
| LWP | 0.39 (0.18) | 0.31 (0.20) | 0.31 (0.20) | 0.47 (0.17) | 0.50 (0.17) | 0.32 (0.03) | 0.08 (0.04) | 0.09 (0.04) | 0.26 (0.04) | 0.21 (0.04) |
| Fiber traits | ||||||||||
| MFA | −0.90 (0.04) | −0.86 (0.07) | −0.86 (0.07) | −0.79 (0.09) | −0.66 (0.13) | −0.89 (0.01) | −0.62 (0.02) | −0.61 (0.02) | −0.59 (0.03) | −0.56 (0.03) |
| FWT | 0.80 (0.08) | 0.69 (0.12) | 0.69 (0.12) | 0.80 (0.08) | 0.80 (0.09) | 0.52 (0.03) | 0.26 (0.04) | 0.27 (0.04) | 0.54 (0.03) | 0.46 (0.04) |
| FCS | 0.61 (0.13) | 0.50 (0.16) | 0.49 (0.16) | 0.56 (0.14) | 0.49 (0.16) | 0.32 (0.04) | 0.13 (0.04) | 0.14 (0.04) | 0.34 (0.04) | 0.28 (0.04) |
| FRW | −0.02 (0.20) | −0.19 (0.20) | −0.19 (0.20) | −0.23 (0.19) | −0.40 (0.18) | −0.09 (0.04) | −0.06 (0.04) | −0.07 (0.04) | −0.09 (0.04) | −0.09 (0.04) |
| FTW | −0.02 (0.20) | 0.02 (0.20) | 0.01 (0.20) | −0.03 (0.19) | −0.10 (0.20) | −0.27 (0.04) | −0.19 (0.04) | −0.19 (0.04) | −0.27 (0.04) | −0.25 (0.04) |
| Growth traits | ||||||||||
| DBH | −0.45 (0.18) | −0.26 (0.21) | −0.25 (0.22) | −0.32 (0.20) | −0.32 (0.21) | −0.40 (0.03) | −0.29 (0.04) | −0.29 (0.04) | −0.33 (0.03) | −0.33 (0.04) |
| HGT | −0.38 (0.19) | −0.18 (0.21) | −0.17 (0.21) | −0.22 (0.20) | −0.26 (0.20) | −0.12 (0.04) | −0.10 (0.04) | −0.10 (0.04) | −0.08 (0.04) | −0.09 (0.04) |
| VOL | −0.44 (0.19) | −0.24 (0.21) | −0.23 (0.22) | −0.30 (0.20) | −0.30 (0.21) | −0.37 (0.03) | −0.27 (0.04) | −0.27 (0.04) | −0.30 (0.04) | −0.29 (0.04) |
| Selection Traits | Weights | Target Traits | ||||
|---|---|---|---|---|---|---|
| DBH | VOL | MOESILV | DENSILV | MFA | ||
| DBH | - | 9.40 | 19.75 | −6.47 | −2.90 | 5.66 |
| MOESILV | - | −5.69 | −11.85 | 19.27 | 7.02 | −19.62 |
| DBH & MOESILV | [1 1.0] | 4.04 | 8.61 | 9.57 | 2.95 | −10.75 |
| DBH & MOEDENSILV | [1 1.0] | 5.51 | 11.68 | 8.45 | 3.09 | −8.65 |
| DBH & MOEDENRES.TB | [1 1.1] | 4.95 | 10.03 | 9.83 | 4.31 | −8.60 |
| DBH & MOEDENCONST | [1 0.6] | 6.55 | 13.84 | 5.77 | 1.49 | −7.34 |
| DBH & DENSILV | [1 0.05] | 5.39 | 11.63 | 3.65 | 2.24 | −1.94 |
| DBH & DENRES.TB | [1 0.04] | 3.16 | 6.87 | 5.93 | 4.50 | −0.86 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fundova, I.; Funda, T.; Wu, H.X. Non-Destructive Assessment of Wood Stiffness in Scots Pine (Pinus sylvestris L.) and its Use in Forest Tree Improvement. Forests 2019, 10, 491. https://doi.org/10.3390/f10060491
Fundova I, Funda T, Wu HX. Non-Destructive Assessment of Wood Stiffness in Scots Pine (Pinus sylvestris L.) and its Use in Forest Tree Improvement. Forests. 2019; 10(6):491. https://doi.org/10.3390/f10060491
Chicago/Turabian StyleFundova, Irena, Tomas Funda, and Harry X. Wu. 2019. "Non-Destructive Assessment of Wood Stiffness in Scots Pine (Pinus sylvestris L.) and its Use in Forest Tree Improvement" Forests 10, no. 6: 491. https://doi.org/10.3390/f10060491
APA StyleFundova, I., Funda, T., & Wu, H. X. (2019). Non-Destructive Assessment of Wood Stiffness in Scots Pine (Pinus sylvestris L.) and its Use in Forest Tree Improvement. Forests, 10(6), 491. https://doi.org/10.3390/f10060491