Current and Future Distribution of Ricania shantungensis (Hemiptera: Ricaniidae) in Korea: Application of Spatial Analysis to Select Relevant Environmental Variables for MaxEnt and CLIMEX Modeling

Abstract
1. Introduction
2. Materials and Methods
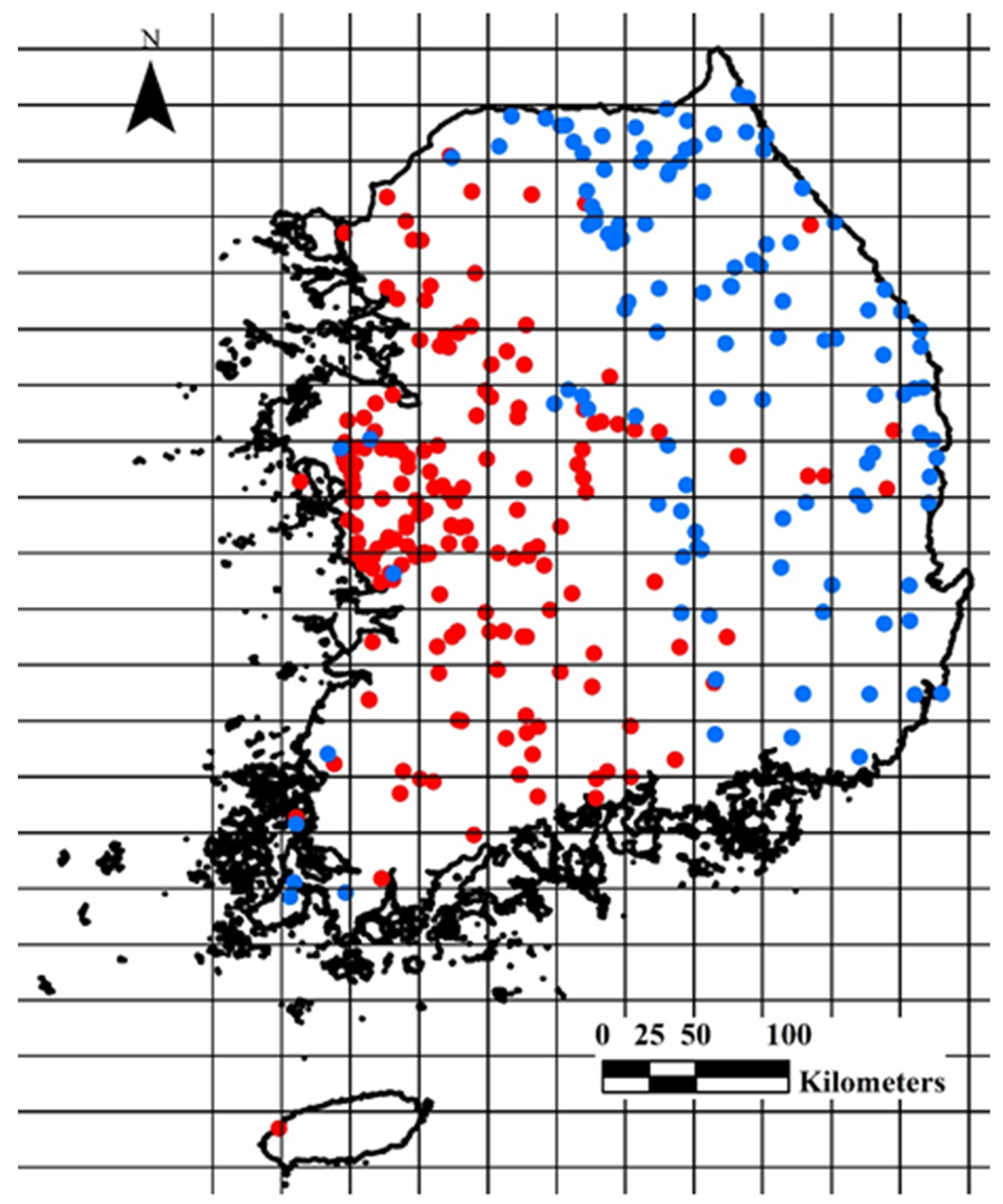
2.1. Data Collection for the Presence and Absence of R. shantungensis
2.2. Environmental, Traffic, Footprint, and Landcover Data Collection
2.3. Spatial Association between Environmental Variables and Distribution of R. shantungensis
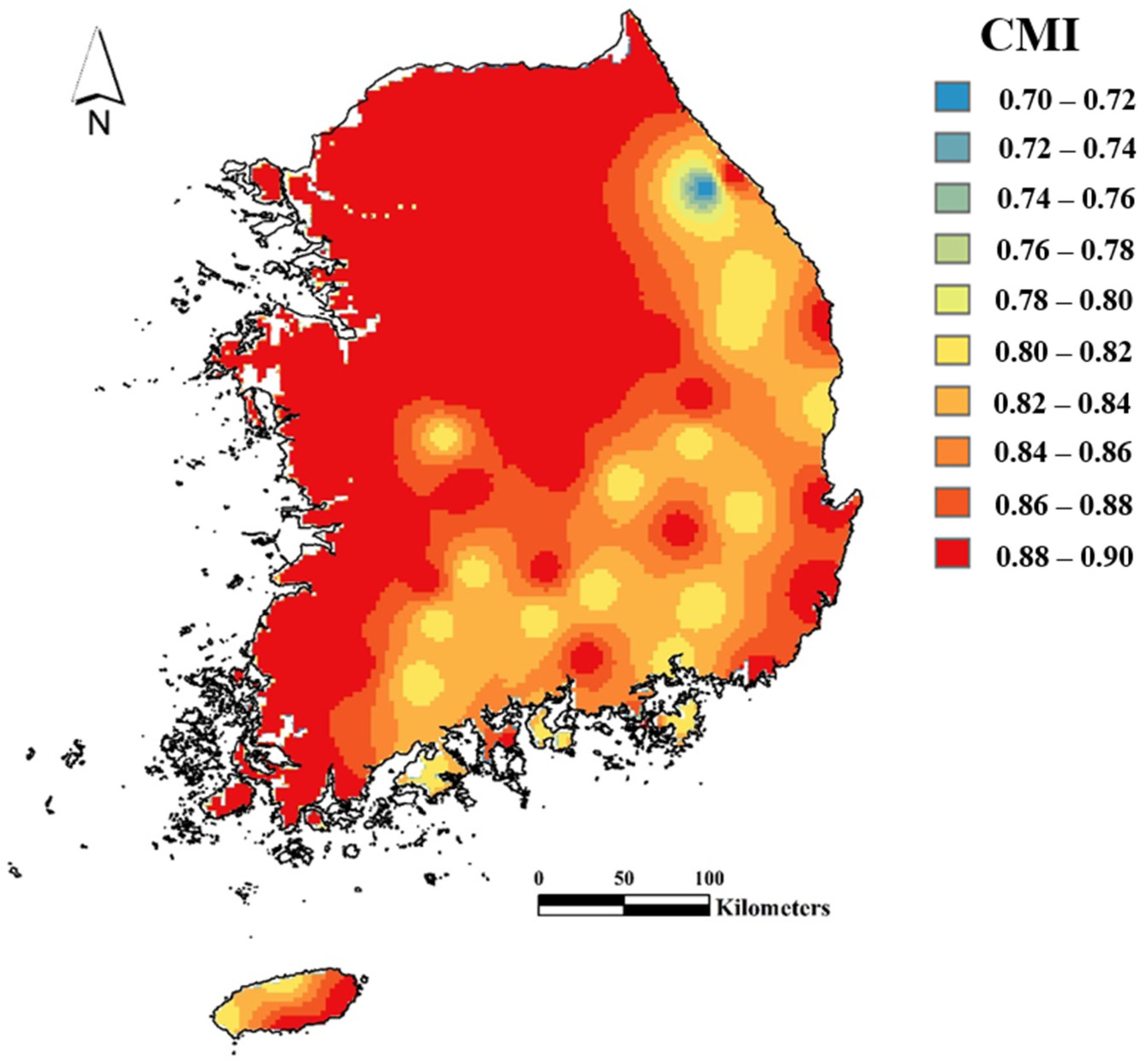
2.4. Prediction of Habitat Suitability of R. shantungensis with CLIMEX
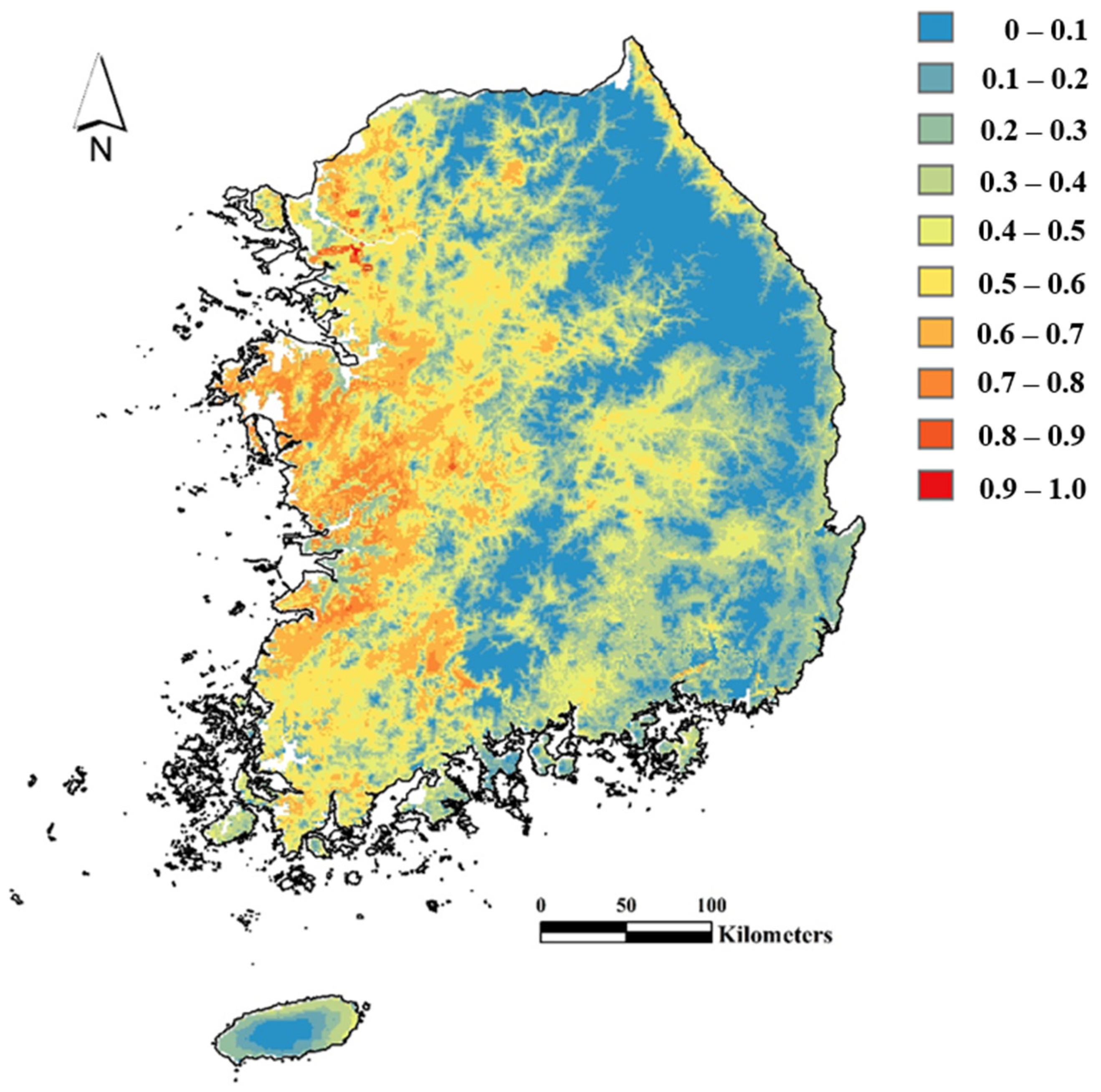
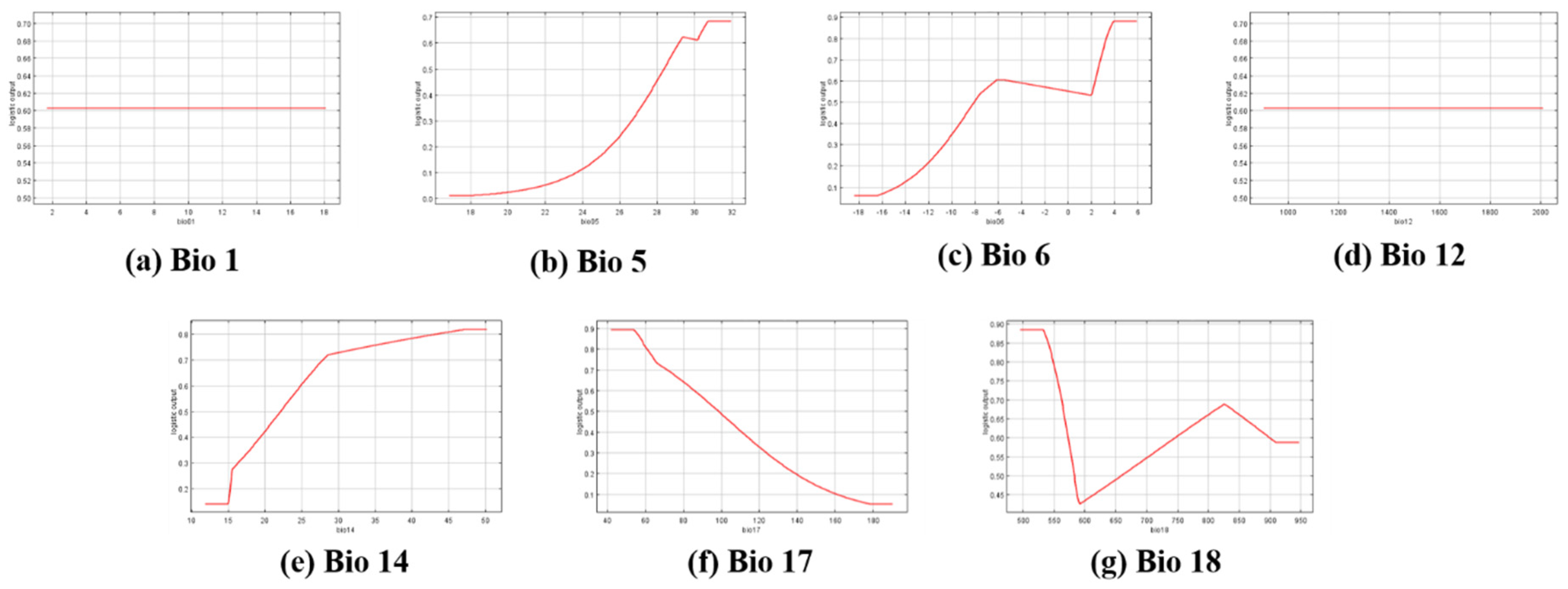
2.5. Prediction of Current Distribution for R. shantungensis with MaxEnt
2.6. Comparison between the Expected Occurrence by MaxEnt and the Actual Occurrence of R. shantungensis
2.7. Prediction of Future Distributions of R. shantungensis in Korea
3. Results
3.1. Habitat Suitability of R. shantungensis with CLIMEX
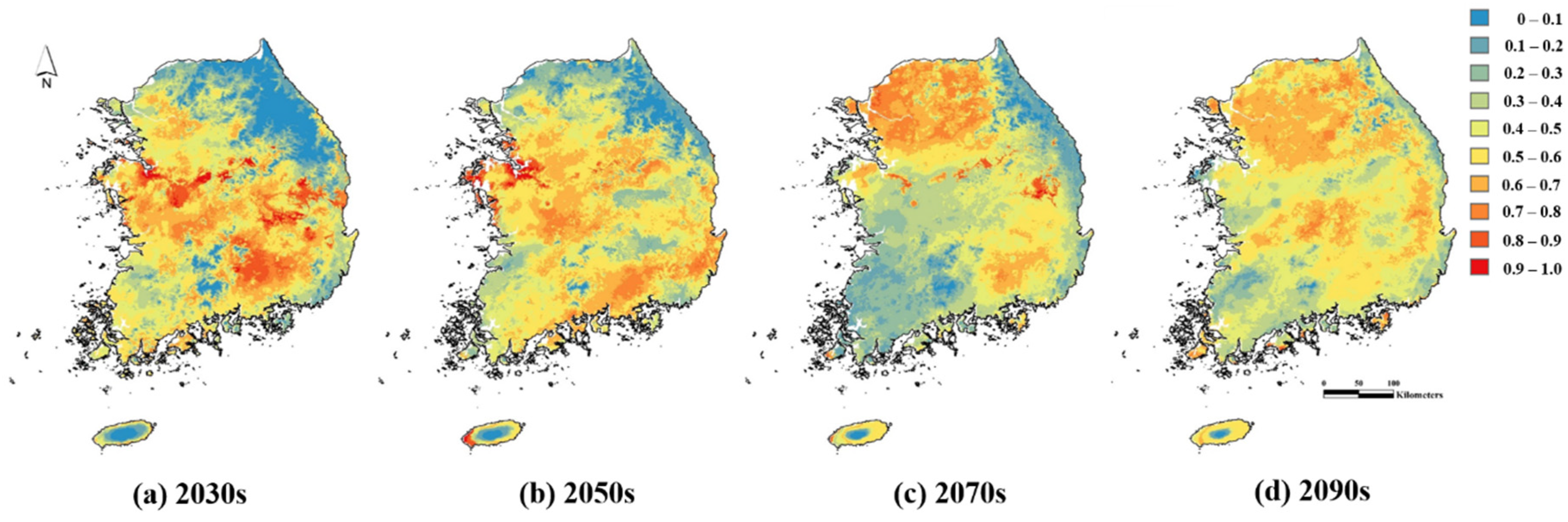
3.2. Prediction of Current and Future Distribution of R. shantungensis with MaxEnt
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jo, S.J. Study on the control and ecology of Ricania shantungensis. J. Tree Health 2014, 19, 35–44. [Google Scholar]
- Kang, D.W.; Ha, J.B.; Lee, S.B.; Han, I.Y.; Kwon, J.H.; Choi, Y.J. Occurrence characteristics and distribution of Ricania shantungensis in Gyungsangnam-Do. In Proceedings of the 2016 Fall Meeting of Korean Society of Applied Entomology, Buyeo Lotte Resorts, Buyeo, Korea, 20–21 October 2016. [Google Scholar]
- Hong, S.J.; Lee, K.J.; Kim, S.T.; No, H.I.; Jung, J.Y. Current distribution and status of Ricania shantungensis in Korea. In Proceedings of the 2017 Fall Meeting of Korean Society of Applied Entomology, Wellihillipark Resort, Hoengseoung, Korea, 26–27 October 2017. [Google Scholar]
- Kim, D.E.; Lee, H.; Kim, M.J.; Lee, D.-H. Predicting the potential habitat, host plants, and geographical distribution of Pochazia shantungensis (Hemiptera: Ricaniidae) in Korea. Kor. J. Appl. Entomol. 2015, 54, 179–189. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Pearson, R.G. Species’ Distribution Modeling for Conservation Educators and Practitioners. Synthesis. American Museum of Natural History. Available online: http://ncep.amnh.org (accessed on 17 September 2018).
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudik, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Zeng, Y.; Low, B.W.; Yeo, D.C.J. Novel methods to select environmental variables in MaxEnt: A case study using invasive crayfish. Ecol. Model. 2016, 341, 5–13. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Ann. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Peterson, A.T. Predicting species distributions from small numbers of occurrence records: a test case using cryptic geckos in Madagascar. J. Biogeo. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Jung, J.-M.; Lee, W.-H.; Jung, S. Insect distribution in response to climate change based on a model: Review of function and use of CLIMEX. Entomol. Res. 2016, 46, 223–235. [Google Scholar] [CrossRef]
- Seo, C.-W.; Park, Y.-R.; Choi, Y.-S. Comparison of species distribution models according to location data. J. Korean Soc. Geospat. Inform. Syst. 2008, 16, 59–64. [Google Scholar]
- Sutherst, R.W.; Maywald, G.F.; Kriticos, D.J. CLIMEX Version 3 User’s Guide; Hearne Scientific Software Pty Ltd.: Melbourne, Australia, 2007. [Google Scholar]
- Kwon, D.H.; Kim, S.-J.; Kang, T.-J.; Lee, J.H.; Kim, D.H. Analysis of the molecular phylogenetics and genetic structure of an invasive alien species, Ricania shantungensis, in Korea. J. Asia Pac. Entomol. 2017, 20, 901–906. [Google Scholar] [CrossRef]
- Chou, I.; Lu, J.S.; Huang, J.; Wang, S. Economic Insect Fauna of China, Fasc. 36; Homoptera Fulgoroidae. Science Press: Beijing, China, 1985. [Google Scholar]
- Hijmans, R.J.; Guarion, L.; Mathur, P. DIVA-GIS. Free GIS for Biodiversity Research, Version 7.5. Available online: http://www.diva-gis.org/ (accessed on 13 September 2018).
- Baek, S. Spatial and Temporal Distribution of Hemlock Woolly Adelgid (Hemiptera: Adelgidae) ovisacs And Its Associations with the Environment. Master Thesis, West Virginia University, Morgantown, WV, USA, 2015. [Google Scholar]
- Perry, J.N. Spatial analysis by distance indices. J. Anim. Ecol. 1995, 64, 303–314. [Google Scholar] [CrossRef]
- Perry, J.N.; Dixon, P. A new method for measuring spatial association in ecological count data. Ecoscience 2002, 9, 133–141. [Google Scholar] [CrossRef]
- Park, Y.-L.; Perring, T.M.; Farrar, C.A.; Gispert, C. Spatial and temporal distributions of two sympatric Homalodisca spp. (Hemiptera: Cicadellidae): Implications for areawide pest management. Agric. Ecosyst. Environ. 2006, 113, 168–174. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Kramer-Schadt, S.; Niedballa, J.; Pilgrim, J.D.; Schröder, B.; Lindenborn, J.; Reinfelder, V.; Stillfried, M.; Heckmann, I.; Scharf, A.K.; Augeri, D.M.; et al. The importance of correcting for sampling bias in MaxEnt species distribution models. Divers. Distrib. 2013, 36, 1366–1379. [Google Scholar] [CrossRef]
- Boria, R.A.; Olson, L.E.; Goodman, S.M.; Anderson, R.P. Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecol. Model. 2014, 275, 73–77. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchman, C.; Carl, G.; Carré, G.; García Marquéz, J.R.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. Maxent Software for Modeling Species Niches and Distributions (Version 3.4.1). Available online: http://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 13 September 2018).
- SAS Institute. SAS/STAT User’s Guide, Version 9.3; SAS Institute: Cary, NC, USA, 2011. [Google Scholar]
- Bradie, J.; Leung, B. A quantitative synthesis of the importance of variables used in MaxEnt species distribution models. K. Biogeogr. 2017, 44, 1344–1361. [Google Scholar] [CrossRef]
- Petitpierre, B.; Broennimann, O.; Kueffer, C.; Daehler, C.; Guisan, A. Selecting predictors to maximize transferability of species distribution models: Lessons from cross-continental plant invasions. Global Ecol. Biogeogr. 2017, 26, 275–287. [Google Scholar] [CrossRef]
- Braunisch, V.; Coppers, J.; Arlettaz, R.; Suchant, R.; Schmid, H.; Bollmann, K. Selecting from correlated climate variables: A major source of uncertainty for predicting species distributions under climate change. Ecography 2013, 36, 971–983. [Google Scholar] [CrossRef]
- Brotons, L.; Thuiller, W.; Araujo, M.B.; Hirzel, A.H. Presence-absence versus presence-only modeling methods for predicting bird habitat suitability. Ecography 2004, 27, 437–448. [Google Scholar] [CrossRef]
- Hirzel, A.; Guisan, A. Which is the optimal sampling strategy for habitat suitability modeling. Ecol. Model. 2002, 157, 331–341. [Google Scholar] [CrossRef]
- Chapman, R.F. The Insects Structure and Function, 3rd ed.; Harvard University Press: Cambridge, MA, USA, 1982. [Google Scholar]
- Choi, Y.S.; Seo, H.Y.; Jo, S.H.; Hwang, I.S.; Yee, Y.S.; Park, D.G. Host preference of Ricania spp. (Hemiptera: Ricaniidae) at different developmental stages. Kor. J. Appl. Entomol. 2017, 56, 319–329. [Google Scholar]
- Kim, D.H.; Yang, C.R.; Kim, H.H.; Seo, M.H.; Yun, J.B. Effect of moisture content of pruned blueberry and peach twigs on hatchability of Ricania shantungensis (Hemiptera: Ricaniidae) eggs. Kor. J. Appl. Entomol. 2017, 56, 357–363. [Google Scholar]
- Duque-Lazo, J.; van Gils, H.; Groen, T.A.; Navarro-Cerrilo, R.M. Transferability of species distribution models: The case of phytophthora cinnamomi in Southwest Spain and Southwest Australia. Ecol. Model. 2016, 320, 62–70. [Google Scholar] [CrossRef]
- Lockwood, J.L.; Hoopes, M.A.; Marchetti, M.P. Invasion Ecology; Blackwell Publishing: Malden, MA, USA, 2007. [Google Scholar]
- Liebhold, A.M.; Tobin, P.C. Population ecology of insect invasions and their management. Annu. Rev. Entomol. 2008, 53, 387–408. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Unit | Range |
|---|---|---|
| Bio1 = Annual mean temperature | °C | 2.6–16.9 |
| Bio2 = Mean diurnal range (Mean of monthly (max temp.–min temp.)) | °C | 5.0–12.9 |
| Bio3 = Isothermality (Bio2/Bio7 × 100) | - | 19.6–32.9 |
| Bio4 = Temperature seasonality (standard deviation × 100) | - | 707.6–1072.1 |
| Bio5 = Max temperature of warmest month | °C | 17.7–30.7 |
| Bio6 = Min temperature of coldest month | °C | −18.8–4.1 |
| Bio7 = Temperature annual range (Bio5–Bio6) | °C | 25.2–40.4 |
| Bio8 = Mean temperature of wettest quarter | °C | 13.0–25.4 |
| Bio9 = Mean temperature of driest quarter | °C | −11.2–10.5 |
| Bio10 = Mean temperature of warmest quarter | °C | 13.0–25.9 |
| Bio11 = Mean temperature of coldest quarter | °C | −11.2–8.3 |
| Bio12 = Annual precipitation | °C | 995–1918 |
| Bio13 = Precipitation of wettest month | mm | 208–403 |
| Bio14 = Precipitation of driest month | mm | 15–47 |
| Bio15 = Precipitation seasonality (coefficient of variation) | - | 59.5–108.0 |
| Bio16 = Precipitation of wettest quarter | mm | 557–1023 |
| Bio17 = Precipitation of driest quarter | mm | 54–178 |
| Bio18 = Precipitation of warmest quarter | mm | 530–910 |
| Bio19 = Precipitation of coldest quarter | mm | 54–182 |
| Elevation | m | −3–1819 |
| Traffic volume | ea | 834–176,191 |
| Footprint | - | 12–100 |
| Landcover | - | - |
| Environmental Variables | X1 | P | Environmental Variables | X | P |
|---|---|---|---|---|---|
| Bio 1 | 0.2466 | 0.0006 | Bio 12 | 0.3011 | 0.0011 |
| Bio 2 | −0.0984 | 0.8373 | Bio 13 | 0.2142 | 0.0207 |
| Bio 3 | −0.1692 | 0.9442 | Bio 14 | 0.4088 | <0.0001 |
| Bio 4 | 0.0413 | 0.3443 | Bio 15 | −0.016 | 0.5625 |
| Bio 5 | 0.3881 | <0.0001 | Bio 16 | 0.2688 | 0.0035 |
| Bio 6 | 0.2491 | 0.0046 | Bio 17 | 0.2937 | 0.0027 |
| Bio 7 | 0.0498 | 0.3169 | Bio 18 | 0.3835 | <0.0001 |
| Bio 8 | 0.3461 | <0.0001 | Bio 19 | 0.2715 | 0.0052 |
| Bio 9 | 0.1235 | 0.1078 | Elevation | −0.2611 | 0.9956 |
| Bio 10 | 0.3812 | <0.0001 | Traffic | 0.0304 | 0.3830 |
| Bio 11 | 0.2293 | 0.0151 | Footprint | 0.0833 | 0.2153 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baek, S.; Kim, M.-J.; Lee, J.-H. Current and Future Distribution of Ricania shantungensis (Hemiptera: Ricaniidae) in Korea: Application of Spatial Analysis to Select Relevant Environmental Variables for MaxEnt and CLIMEX Modeling. Forests 2019, 10, 490. https://doi.org/10.3390/f10060490
Baek S, Kim M-J, Lee J-H. Current and Future Distribution of Ricania shantungensis (Hemiptera: Ricaniidae) in Korea: Application of Spatial Analysis to Select Relevant Environmental Variables for MaxEnt and CLIMEX Modeling. Forests. 2019; 10(6):490. https://doi.org/10.3390/f10060490
Chicago/Turabian StyleBaek, Sunghoon, Min-Jung Kim, and Joon-Ho Lee. 2019. "Current and Future Distribution of Ricania shantungensis (Hemiptera: Ricaniidae) in Korea: Application of Spatial Analysis to Select Relevant Environmental Variables for MaxEnt and CLIMEX Modeling" Forests 10, no. 6: 490. https://doi.org/10.3390/f10060490
APA StyleBaek, S., Kim, M.-J., & Lee, J.-H. (2019). Current and Future Distribution of Ricania shantungensis (Hemiptera: Ricaniidae) in Korea: Application of Spatial Analysis to Select Relevant Environmental Variables for MaxEnt and CLIMEX Modeling. Forests, 10(6), 490. https://doi.org/10.3390/f10060490