Variation of Leaf Carbon Isotope in Plants in Different Lithological Habitats in a Karst Area

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Plant Sample Collection
2.3. Soil Sample Collection
2.4. Measurement Methods
2.5. Integrated Fertility Indices (IFIs)
2.6. Data Processing
3. Results
3.1. Δ13C Values of the Dominant Species in the Three Lithological Soil Types
3.2. Δ13C Values of Different Dominant Species
3.3. Relationship between the Δ13C Value of Dominant Species and Comprehensive Lithological Soil Conditions
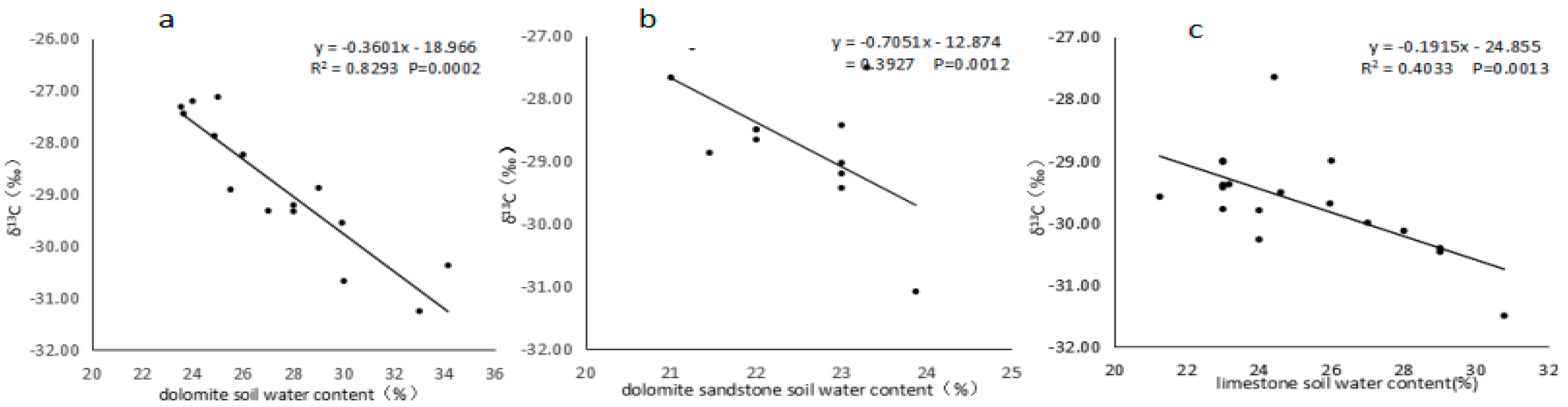
3.4. Relationship between the Δ13C Value of the Dominant Species and the Soil Water Content
4. Discussion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| WUE | Water-use efficiency |
| IRMS | Isotope ratio mass spectrometry |
| PDB | Pee Dee Belemnite standard |
| PCA | Principal component analysis |
| CA | Correlation analysis |
| IFI | Integrated fertility index |
References
- Zhu, S.Q. Study on Forest Ecology in Karst, 2nd ed.; People’s Publishing House of Guizhou Province: Guizhou, China, 2002; pp. 21–53. [Google Scholar]
- He, K.Q.; Du, R.L.; Jiang, W.F. Contrastive analysis of karst collapses and the distribution rules in northern and southern China. Environ. Earth Sci. 2010, 59, 1309–1318. [Google Scholar]
- Yu, L.F.; Zhu, S.Q.; Ye, J.Z. Preliminary study on the adaptability of tolerate-drought for different species group in karst forest. J. Nanjing For. Univ. 2002, 26, 18–22. [Google Scholar]
- Arslan, A.; Zapata, F.; Kumarasinghe, K.S. Carbon isotope discrimination as indicator of water-use efficiency of spring wheat as affected by salinity and gypsum addition. Commun. Soil Sci. Plant Anal. 1999, 30, 2681–2693. [Google Scholar] [CrossRef]
- Ehleringer, J.R.; Hall, A.E.; Farquhar, G.D. Carbon and water relations in desert plants: An isotopic perspective. Stable Isot. Carbon-Water Relat. 1993, 72, 155–172. [Google Scholar]
- Wildy, D.T.; Pate, J.S.; Sefcik, L.T. Water-use efficiency of a mallee eucalypt growing naturally and in short-rotation coppice cultivation. Plant Soil 2004, 262, 111–128. [Google Scholar] [CrossRef]
- Akhter, J.; Mahmood, K.; Tasneem, M. Comparative water-use efficiency of Sporobolus arabicus and Leptochloa fusca and its relation with carbon-isotope discrimination under semi-arid conditions. Plant Soil 2003, 249, 263–269. [Google Scholar] [CrossRef]
- Jędrysek, M.O.; Krąpiec, M.; Skrzypek, G.; Kałużny, A. Air-pollution effect and paleotemperature scale versus δ13C records in tree rings and in a peat core (Southern Poland). Water Air Soil Pollut. 2003, 145, 359–375. [Google Scholar] [CrossRef]
- Máguas, C.; Griffiths, S.H. Applications of Stable Isotopes in Plant Ecology; Progress in Botany; Springer: Berlin, Germany, 2003; Volume 64, pp. 472–505. [Google Scholar]
- Hatté, C.; Schwartz, D. Reconstruction of paleoclimates by isotopic analysis: What can the fossil isotopic record tell us about the plant life of past environments? Phytochem. Rev. 2003, 2, 163. [Google Scholar] [CrossRef]
- Renard, D.; Birk, J.J.; Glaser, B. Origin of mound-field landscapes: A multi-proxy approach combining contemporary vegetation, carbon stable isotopes and phytoliths. Plant Soil 2012, 351, 337–353. [Google Scholar] [CrossRef]
- Wei, J.; Liu, W.G.; Cheng, J.M.; Li, W.J. δ13C values of plants as indicators of soil water content in modern ecosystems of the Chinese Loess Plateau. Ecol. Eng. 2015, 77, 51–59. [Google Scholar] [CrossRef]
- Xu, Y.F.; Wang, K.L.; Qi, X.K. Difference in spatial-temporal change of karst vegetation in dolomite and limestone based on TM image. Acta Ecol. Sin. 2016, 36, 180–189. [Google Scholar]
- Wang, L.C.; Hu, W.X.; Wang, X.L. Variation of Sr content and ~(87) Sr/~(86) Sr isotope fractionation during dolomitization and their implications. Oil Gas Geol. 2016, 37, 464–471. [Google Scholar]
- Zhou, D.Q.; Wang, S.J.; Liu, X.M. Study on geo-chemical process in limest stone soil profile. Earth Environ. 2005, 33, 31–38. [Google Scholar]
- Gao, F.H.; Zhang, Y.S.; Pu, X.G. Type of dolomization and formation mechanism: A case study of Palaeogene Es1 in Qikou Sag. J. Jilin Univ. 2017, 47, 355–369. [Google Scholar]
- Li, Y.B.; Wang, S.J.; Li, R.L. Karst ecosystem soil. J. Ecol. Environ. 2004, 13, 434–438. [Google Scholar]
- He, Z.H.; Chen, X.X.; Liang, H. Study on drought mechanism of typical karst basin with lithologic composite structure-A case study of Guizhou province. Chin. J. Geol. 2015, 50, 340–353. [Google Scholar]
- Yang, C.; Liu, C.Q.; Song, Z.L. Carbon isotope composition of plant leaf in Guizhou karst mountain area. Earth Environ. 2007, 35, 33–38. [Google Scholar]
- Xiong, K.L.; Li, P.; Zhong, Z.F. Remote Sensing of Rocky Desertification in Karst-GIS Typical Study-Take Guizhou as an Example; Geological Publishing House: Beijing, China, 2002; pp. 1–183. [Google Scholar]
- Zhu, S.Q. Preliminary study on forest niche characteristics in Karst. In Study on Forest Ecology in Karst, 1st ed.; Science and Technology Press: Guiyang, China, 1993; pp. 52–62. [Google Scholar]
- Li, R.L.; Wang, S.J.; Zhou, D.Q.; Zhang, D.F. Correlation analysis between lithology and rocky desertification in Guizhou karst area. Acta Geogr. Sin. 2003, 58, 315–320. [Google Scholar]
- Fu, Y.H.; Huang, Z.S.; Yu, L.F. Changes of foliar δ13C value of Quercus fabric in different root underground habitat types in Karst area. Chin. J. Appl. Ecol. 2012, 23, 2961–2967. [Google Scholar]
- Lin, G.H. Stable Isotope Ecology, 1st ed.; Higher Education Press: Beijing, China, 2013; pp. 7–8. [Google Scholar]
- Bao, S.D. Soil and Agricultural Chemistry Analysis; China Agriculture Press: Beijing, China, 1999; pp. 229–291. [Google Scholar]
- Pang, X.Y. Comparative Study of Soil Characteristics during Different Successional Stages for Subalpine Coniferous Forest in Western Sichuan. Master’s Thesis, Sichuan Agricultural University, Yaan, China, 2002; pp. 31–37. [Google Scholar]
- Wei, Y.; Yu, L.F.; Zhang, J.C. Evaluation on soil ecologic fertility quantity during vegetation succession of degraded karst—A case study in Huajiang valley, Guizhou. Carsologica Sin. 2009, 28, 61–67. [Google Scholar]
- Rong, L.; Wang, S.J.; Du, X.L. Responses of foliar δ13C values of woody plants to different karst rocky desertification degrees in Huajiang gorge, Guizhou. Sci. Silvae Sin. 2007, 43, 38–44. [Google Scholar]
- Qu, C.M.; Han, X.G.; Su, P. Characteristics of leaf δ13C value of fragmented rainforest plant in Xishuangbanna of Yunnan and the water use efficiency. Chin. Bull. Bot. 2001, 43, 186–192. [Google Scholar]
- Chen, T.; Ma, J.; Feng, H.Y. Analysis of environment for stable carbon isotope value of typical desert C3 plants in Fukang area. Arid Land Geogr. 2002, 25, 342–345. [Google Scholar]
- Buchmann, N.; Kao, W.Y.; Ehleringer, J.R. Influence of stand structure on carbon-13 of vegetation, solids, and canopy air within deciduous and evergreen forests in Utah, United States. Oecologia 1997, 110, 109–119. [Google Scholar] [CrossRef]
- Yan, C.R.; Han, X.G.; Chen, L.Z. Foliar δ13C within temperate deciduous Forest: Its spatial change and interspecies variation. Acta Bot. Sin. 1998, 40, 853–859. [Google Scholar]
- Gessler, A.; Ferrio, J.P.; Hommel, R. Stable isotopes in tree rings: Towards a mechanistic understanding of isotope fractionation and mixing processes from the leaves to the wood. Tree Physiol. 2014, 34, 796–818. [Google Scholar] [CrossRef]
- Pate, J.S. Carbon isotope discrimination and plant water-use efficiency. In Stable Isotope Techniques in the Study of Biological Processes and Functioning of Ecosystems. Current Plant Science and Biotechnology in Agriculture; Unkovich, M., Pate, J., McNeill, A., Gibbs, D.J., Eds.; Springer: Dordrecht, The Netherlands, 2001; Volume 40, pp. 19–36. [Google Scholar]
- Liu, X.Z.; Gao, C.C.; Su, Q.; Zhang, Y.; Song, Y. Altitudinal trends in δ13C value, stomatal density and nitrogen content of Pinus tabuliformis needles on the southern slope of the middle Qinling Mountains, China. J. Mt. Sci. 2016, 13, 1066–1077. [Google Scholar] [CrossRef]
- Schuster, W.S.F.; Sandquist, D.R.; Philips, S.L. Comparisons of carbon isotope discrimination in populations of arid land plant species differing in lifespan. Oecologia 1992, 91, 332–337. [Google Scholar] [CrossRef]
- Rong, L.; Wang, S.J.; Du, X.L. Research of relation between δ13C value of common plants and environmental factors in Karst. Environ. Sci. 2008, 29, 2885–2893. [Google Scholar]
- Song, L.; Zhu, J.; Li, M. Intrinsic water use efficiency in wet and dry years at young and old plantations of Pinus sylvestris var. mongolica in semiarid China. J. For. Res. 2015, 20, 263–271. [Google Scholar] [CrossRef]
- Santini, F.; Ferrio, J.P.; Hereş, A.M. Scarce population genetic differentiation but substantial spatiotemporal phenotypic variation of water-use efficiency in Pinus sylvestris at its western distribution range. Eur. J. For. Res. 2018, 137, 863–878. [Google Scholar] [CrossRef]
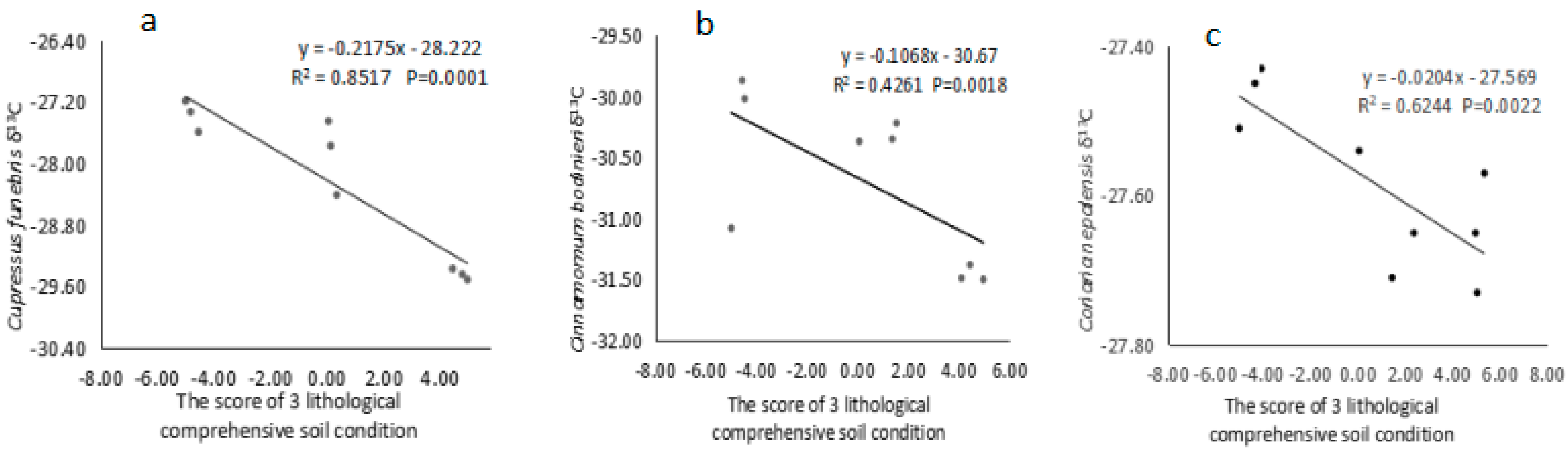

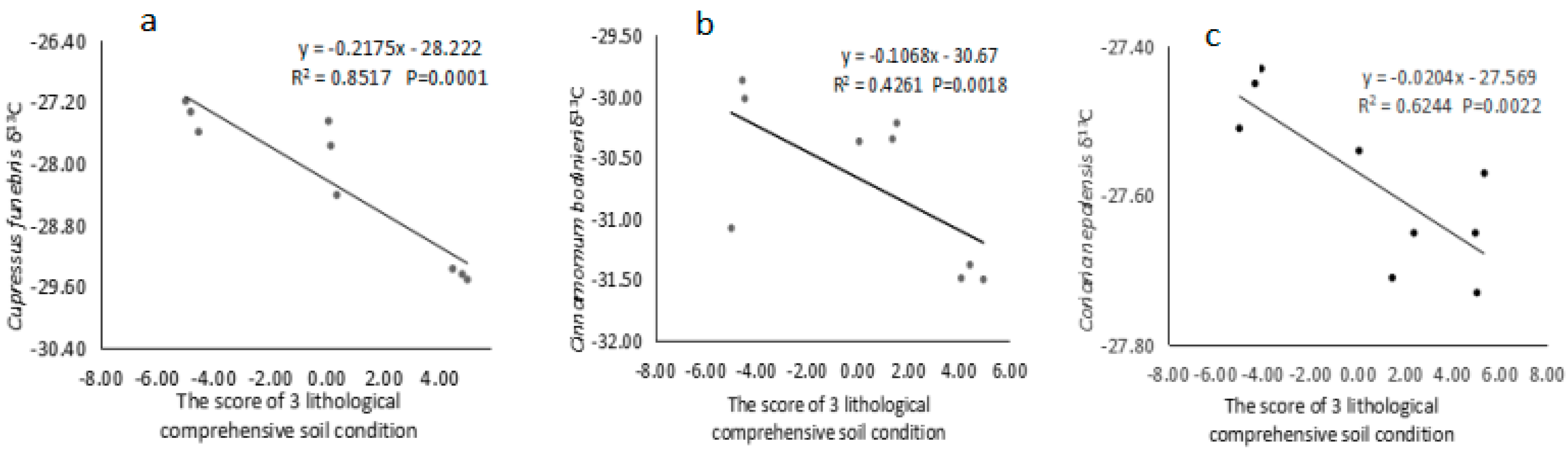{kind=link}
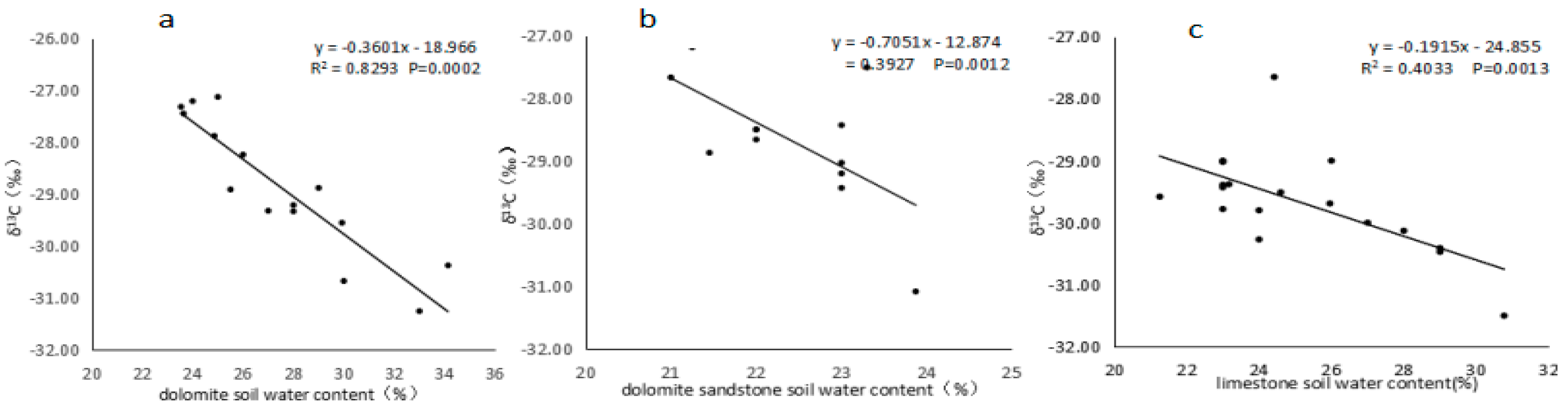{kind=link}
| Lithology | Dominant Species | Minimum Value (‰) | Maximum Value (‰) | Mean (‰) ± Standard Deviation | Significant Difference Across sites |
|---|---|---|---|---|---|
| Dolomite | Cinnamomum bodinieri | −30.70 | −29.97 | −30.37 ± 0.36 a | -- |
| Cupressus funebris | −27.60 | −27.26 | −27.45 ± 0.15 d | 1 | |
| Coriaria nepalensis | −28.10 | −27.70 | −27.88 ± 0.17 d | -- | |
| Betula luminifera | −29.94 | −28.96 | −29.55 ± 0.47 b | -- | |
| Alnus cremastogyne | −29.40 | −28.39 | −28.91 ± 0.41 c | 1 | |
| Dolomite sandstone | Cinnamomum bodinieri | −32.49 | −30.10 | −31.08 ± 1.23 a | -- |
| Cupressus funebris | −27.88 | −26.20 | −27.19 ± 0.76 c | 1 | |
| Coriaria nepalensis | −28.14 | −26.90 | −27.51 ± 0.60 c | -- | |
| Platycarya longipes | −29.62 | −28.30 | −28.87 ± 0.57 b | -- | |
| Limestone | Cinnamomum bodinieri | −32.00 | −30.44 | −31.50 ± 0.72 a | -- |
| Cupressus funebris | −29.80 | −29.32 | −29.51 ± 0.21 b | 2 | |
| Coriaria nepalensis | −28.60 | −26.99 | −27.65 ± 0.77 c | -- | |
| Betula luminifera | −29.80 | −29.25 | −29.58 ± 0.27 b | -- | |
| Alnus cremastogyne | −30.00 | −29.26 | −29.69 ± 0.31 b | 2 | |
| Platycarya longipes | −29.90 | −28.20 | −29.38 ± 0.80 b | -- |
| Soil Bulk Density (x1, g/cm3) | Soil Water Content (x2, %) | Organic Matter (x3, g/kg) | Total Nitrogen (x4, g/kg) | Total Phosphorus (x5, g/kg) | Total Potassium (x6, g/kg) | Alkali-Hydrolysable Nitrogen (x7, g/kg) | Available Phosphorus (x8, mg/kg) | Olsen-K (x9, mg/kg) | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Dolomite | 0.998 | 29.610 | 55.671 | 2.889 | 0.467 | 17.797 | 183.255 | 2.087 | 81.923 | |
| −1.068 | 0.956 | −0.100 | −0.178 | −0.510 | 0.809 | −0.023 | 0.237 | −0.372 | ||
| Dolomite sandstone | 1.078 | 23.280 | 47.991 | 2.453 | 0.424 | 7.120 | 143.903 | 1.639 | 76.700 | |
| 0.914 | −1.039 | −0.946 | −0.899 | −0.642 | −1.118 | −0.988 | −1.097 | −0.761 | ||
| Limestone | 1.047 | 26.840 | 66.088 | 3.650 | 1.018 | 15.028 | 225.450 | 2.296 | 102.105 | |
| 0.154 | 0.083 | 1.046 | 1.077 | 1.152 | 0.309 | 1.011 | 0.860 | 1.133 |
| P1 | P2 | Overall Evaluation | ||||
|---|---|---|---|---|---|---|
| Lithology Type | y1 | Rank | y2 | Rank | IFIs | Rank |
| Dolomite | 0.997 | 2 | −2.669 | 3 | 0.047 | 2 |
| Dolomite sandstone | −7.111 | 3 | 1.033 | 2 | −5.001 | 3 |
| Limestone | 6.114 | 1 | 1.636 | 1 | 4.954 | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, J.; Yu, L.; Huang, Z. Variation of Leaf Carbon Isotope in Plants in Different Lithological Habitats in a Karst Area. Forests 2019, 10, 356. https://doi.org/10.3390/f10040356
Zou J, Yu L, Huang Z. Variation of Leaf Carbon Isotope in Plants in Different Lithological Habitats in a Karst Area. Forests. 2019; 10(4):356. https://doi.org/10.3390/f10040356
Chicago/Turabian StyleZou, Jun, Lifei Yu, and Zongsheng Huang. 2019. "Variation of Leaf Carbon Isotope in Plants in Different Lithological Habitats in a Karst Area" Forests 10, no. 4: 356. https://doi.org/10.3390/f10040356
APA StyleZou, J., Yu, L., & Huang, Z. (2019). Variation of Leaf Carbon Isotope in Plants in Different Lithological Habitats in a Karst Area. Forests, 10(4), 356. https://doi.org/10.3390/f10040356