N Isotope Fractionation in Tree Tissues During N Reabsorption and Remobilization in Fagus crenata Blume

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Sample Collection and Treatments
2.3. Soil Analysis
2.3.1. Total Nitrogen and δ15N Isotope Analysis
2.3.2. Amino Acid Analysis
2.3.3. Whole-Tree N Calculations
2.3.4. Statistical Analysis
3. Results
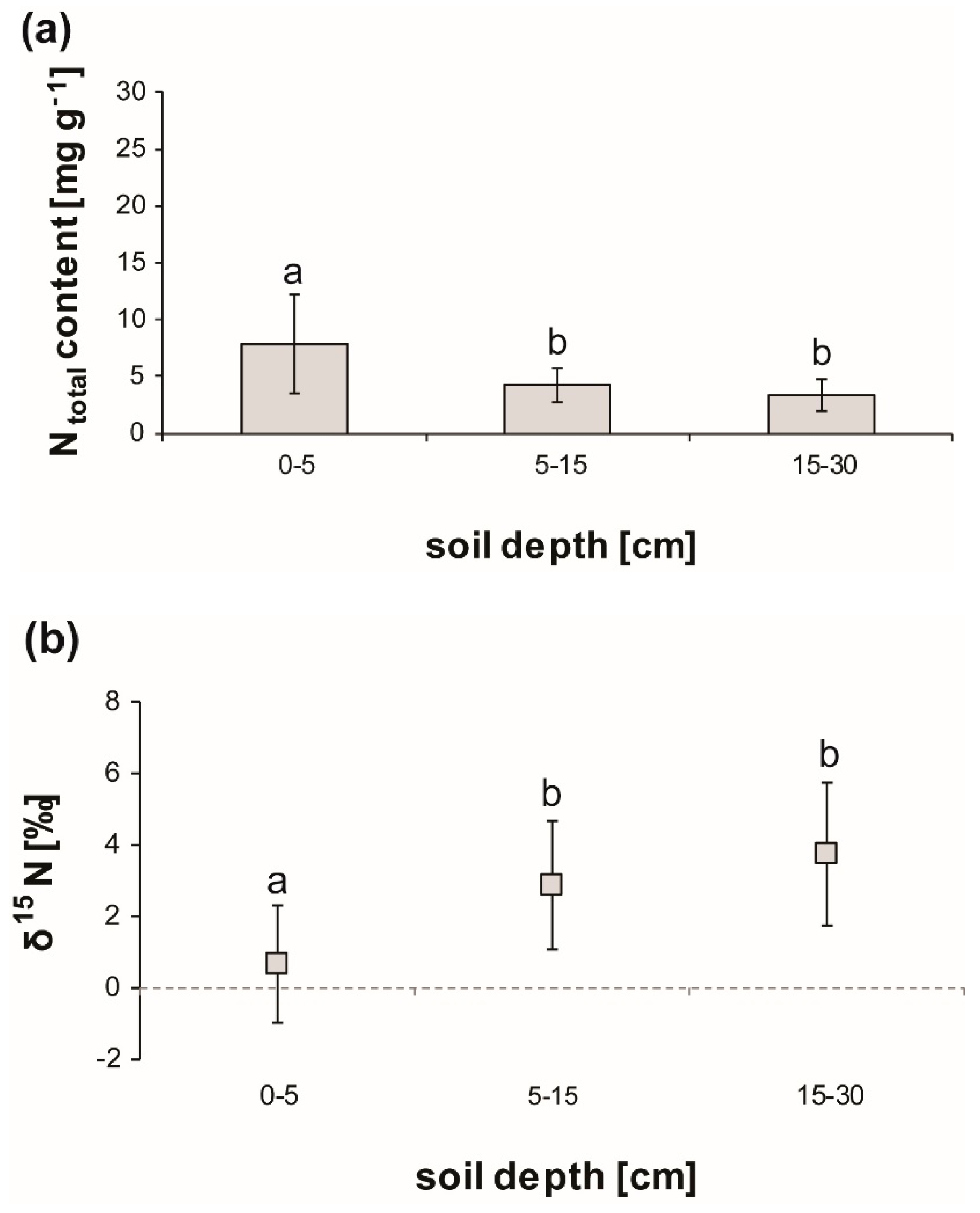
3.1. Soil Characteristics
3.2. Total Nitrogen Content in Soil and Tissues
3.3. δ15N Isotope Analysis
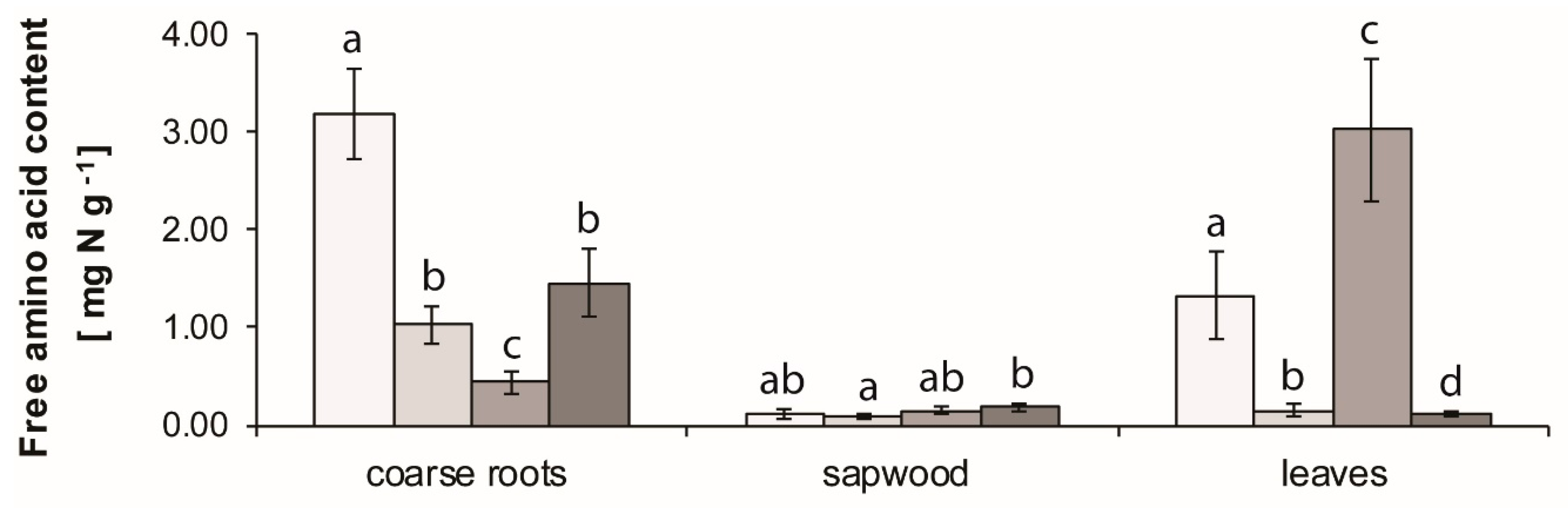
3.4. Amino Acid Analysis in Plant Tissues
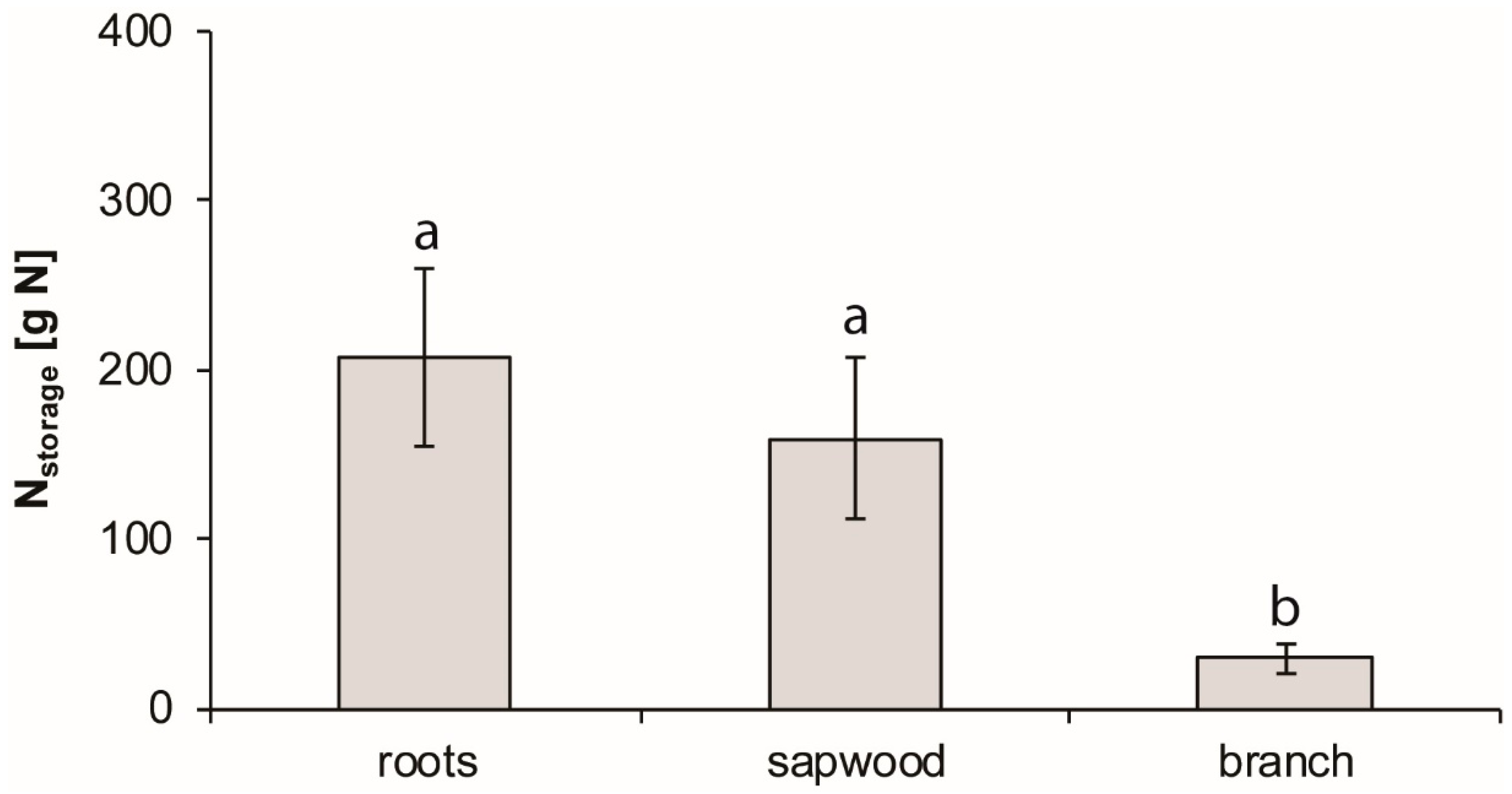
3.5. Whole-Tree Level
4. Discussion
4.1. Reabsorption, Storage, and Remobilization of N in Plant Tissues
4.2. Effect of Soil N Uptake on Plant δ15N
4.3. Effect of Intra-Plant N Movement on δ15N
4.4. Plant Tissues Free Amino Acids
4.5. Correlation between Free Amino Acid Content and Plant Tissues δ15N
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liang, N.; Nagayama, M.; Nakata, M.; Maruyama, K. Growth, photosynthesis and nitrogen content in Japanese beech (Fagus crenata Bl.) seedlings grown under five irradiances. Photosynthetica 1995, 31, 257–268. [Google Scholar]
- Watanabe, M.; Yamaguchi, M.; Matsumura, H.; Kohno, Y.; Izuta, T. Risk assessment of ozone impact on Fagus crenata in Japan: Consideration of atmospheric nitrogen deposition. Eur. J. For. Res. 2012, 131, 475–484. [Google Scholar] [CrossRef]
- Fukushima, T.; Takasuna, H.; Matsui, T.; Nishio, T.; Kyan, T.; Tsunetomi, Y. New phytosociological classification of beech forests in Japan. Jpn. J. Ecol. 1995, 45, 79–98, (In Japanese with English Synopsis). [Google Scholar]
- Shimano, K.; Masuzawa, T. Effects of snow accumulation on survival of beech (Fagus crenata) seed. Plant Ecol. 1998, 134, 235–241. [Google Scholar]
- Yasumura, Y.; Hikosaka, K.; Hirose, T. Resource allocation to vegetative and reproductive growth in relation to mast seeding in Fagus crenata. For. Ecol. Manag. 2006, 229, 228–233. [Google Scholar] [CrossRef]
- Han, Q.; Kabeya, D.; Iio, A.; Kakubari, Y. Masting in Fagus crenata and its influence on the nitrogen content and dry mass of winter buds. Tree Physiol. 2008, 28, 1269–1276. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, Y.; Maruyama, Y.; Chiba, Y.; Kobayashi, M.J.; Joseph, B.; Shimizu, K.K.; Mochida, K.; Hiura, T.; Kon, H.; Satake, A. Nitrogen as a key regulator of flowering in Fagus crenata: Understanding the physiological mechanism of masting by gene expression analysis. Ecol. Lett. 2014, 17, 1299–1309. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Kabeya, D.; Iio, A.; Inagaki, Y.; Kakubari, Y. Nitrogen storage dynamics are affected by masting events in Fagus crenata. Oecologia 2014, 174, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Aerts, R.; Chapin, F.S. The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns. Adv. Ecol. Res. 2000, 30, 1–67. [Google Scholar]
- Kolb, K.J.; Evans, R.D. Implication of leaf nitrogen recycling on the nitrogen isotope composition of deciduous plant tissues. New Phytol. 2002, 156, 57–64. [Google Scholar] [CrossRef]
- Yasumura, Y.; Hikosaka, K.; Matsui, K.; Hirose, T. Leaf-level nitrogen-use efficiency of canopy and understorey species in a beech forest. Funct. Ecol. 2002, 16, 826–834. [Google Scholar] [CrossRef]
- Yasumura, Y.; Onoda, Y.; Hikosaka, K.; Hirose, T. Nitrogen resorption from leaves under different growth irradiance in three deciduous woody species. Plant Ecol. 2005, 178, 29–37. [Google Scholar] [CrossRef]
- Lopez, M.L.; Mizota, C.; Nobori, Y.; Sasaki, T.; Yamanaka, T. Temporal changes in nitrogen acquisition of Japanese black pine (Pinus thunbergii) associated with black locust (Robinia Pseudoacacia). J. For. Res. 2014, 25, 585–589. [Google Scholar]
- Millard, P.; Grelet, G.-A. Nitrogen storage and remobilization by trees: Ecophysiological relevance in a changing world. Tree Physiol. 2010, 30, 1083–1095. [Google Scholar] [CrossRef] [PubMed]
- Dyckmans, J.; Flessa, H. Influence of tree internal N status on uptake and translocation of C and N in beech: A dual 13C and 15N labeling approach. Tree Physiol. 2001, 21, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Millard, P.; Hester, A.; Wendler, R.; Baillier, G. Interspecific defoliation responses of trees depend on sites of winter nitrogen storage. Funct. Ecol. 2001, 15, 535–543. [Google Scholar] [CrossRef]
- Kayama, M.; Makoto, K.; Nomura, M.; Satoh, F.; Koike, T. Nutrient dynamics and carbon partitioning in larch seedlings (Larix kaempferi) regenerated on a serpentine soil in northern Japan. Landsc. Ecol. Eng. 2009, 5, 125–135. [Google Scholar] [CrossRef]
- Leberecht, M.; Dannenmann, M.; Tejedor, J.; Simon, J.; Rennenberg, H.; Polle, A. Segregation of nitrogen use between ammonium and nitrate of ectomycorrhizas and beech trees. Plant Cell Environ. 2016, 39, 2691–2700. [Google Scholar] [CrossRef]
- Tateno, R.; Osada, N.; Terai, M.; Tokuchi, N.; Takeda, Y. Inorganic nitrogen source utilization by Fagus crenata on different soil types. Trees 2005, 19, 477–481. [Google Scholar] [CrossRef]
- Hobbie, E.A.; Högberg, P. Nitrogen isotopes link mycorrhizal fungi and plants to nitrogen dynamics. New Phytol. 2012, 196, 367–382. [Google Scholar]
- Gebauer, G.; Dietrich, P. Nitrogen Isotope Ratios in Different Compartments of a Mixed Stand of Spruce, Larch and Beech Trees and of Understorey Vegetation Including Fungi. Isot. Environ. Health Stud. 1993, 29, 35–44. [Google Scholar] [CrossRef]
- Fotelli, M.N.; Rennenberg, H.; Geßler, A. Effects of Drought on the Competitive Interference of an Early Succsessional Species (Rubus fruticosus) on Fagus sylvatica L. Seedlings: 15N Uptake and Partitioning, Responses of Amino Acids and other N Compounds. Plant Boil 2002, 4, 311–320. [Google Scholar] [CrossRef]
- Chikaraishi, Y.; Ogawa, N.O.; Kashiyama, Y.; Takano, Y.; Suga, H.; Tomitani, A.; Miyashita, M.; Kitazato, H.; Ohkouchi, N. Determination of aquatic food-web structure based on compound-specific nitrogen isotopic composition of amino acids. Limnol. Oceanogr. Methods 2009, 7, 740–750. [Google Scholar] [CrossRef]
- Chikaraishi, Y.; Ogawa, N.O.; Doi, H.; Ohkouchi, N. 15N/14N ratios of amino acids as a tool for studying terrestrial foodwebs: A case study of terrestrial insects (bees, wasps, and hornets). Ecol. Res. 2011, 26, 835–844. [Google Scholar] [CrossRef]
- Food and Agriculture Organization (FAO). Guidelines for Soil Description, 4th ed.; FAO: Rome, Italy, 2006. [Google Scholar]
- Tomlinson, G.; Siegwolf, R.T.W.; Buchmann, N.; Schleppi, P.; Waldner, P.; Weber, P. The mobility of nitrogen across tree-rings of Norway spruce (Picea abies L.) and the effect of extraction method on tree-ring δ 15N and δ 13C values. Rapid Commun. Mass Sprectrometry 2014, 28, 1258–1264. [Google Scholar] [CrossRef]
- Osada, N.; Yasumura, Y.; Ishida, A. Leaf nitrogen distribution in relation to crown architecture in the tall canopy species, Fagus crenata. Oecologia 2014, 175, 1093–1106. [Google Scholar] [CrossRef]
- Crooke, W.M.; Simpson, W.E. Determination of ammonium in Kjedahl digest of crops by an automated procedure. J. Sci. Food Agric. 1971, 22, 9–10. [Google Scholar] [CrossRef]
- Mulvaney, R.L. Nitrogen—Inorganic Forms. In Methods of Soil Analysis. Part 3, Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpoor, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; SSSA Book Series No. 5; SSSA: Madison, WI, USA, 1996; pp. 125–1184. [Google Scholar]
- Miranda, K.M.; Espey, M.G.; Wink, D.A. A rapid, simple spectrophotometric method for simultaneous detection of nitrate and nitrite. Nitric Oxide 2001, 5, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, A.; Ostuka, T.; Nakabayashi, R.; Jikumaru, Y.; Isuzugawa, K.; Murayama, H.; Saito, K.; Shiratake, K. Metabolic Profiling of Developing Pear Fruits Reveals Dynamic Variation in Primary and Secondary Metabolites, Including Plant Hormones. PLoS ONE 2015, 10, e0131408. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Yasuda, Y.; Matsuo, T.; Hoshino, D.; Chiba, Y.; Mori, S. Estimating forest biomass using allometric model in a cool-temperate Fagus crenata forest in the Appi highlands, Iwate, Japan. For. Res. Inst. Res. Rep. 2013, 12, 125–141. [Google Scholar]
- Tateishi, M.; Kumagai, T.; Suyama, Y.; Hiura, T. Differences in transpiration characteristics of Japanese beech trees, Fagus crenata, in Japan. Tree Physiol. 2010, 30, 748–760. [Google Scholar] [CrossRef][Green Version]
- Gebauer, T.; Horna, V.; Leuschner, C. Variability in radial sap flux density patterns and sapwood area among seven co-occurring temperate broad-leaved tree species. Tree Physiol. 2008, 28, 1821–1830. [Google Scholar] [CrossRef]
- Enta, A.; Hayashi, M.; Lopez, C.M.L.; Fujiyoshi, L.; Yamanaka, T.; Oikawa, A.; Seidel, F. Nitrogen resorption and fractionation during leaf senescence in typical tree species in Japan. J. For. Res. 2019, in press. [Google Scholar]
- Ueda, M.U.; Mizumachi, E.; Tokuchi, N. Foliage nitrogen turnover: Differences among nitrogen absorbed at different times by Quercus serrata saplings. Ann. Bot. 2011, 108, 169–175. [Google Scholar] [CrossRef]
- Warren, C.R.; Dreyer, E.; Adams, M.A. Photosynthesis-Rubisco relationships in foliage of Pinus sylvestris in response to nitrogen supply and the proposed role of Rubisco and amino acids as nitrogen stores. Trees 2003, 17, 359–366. [Google Scholar]
- Gessler, A.; Schneider, S.; von Sengbusch, D.; Weber, P. Field and laboratory experiments on net uptake of nitrate and ammonium by the roots of spruce (Picea abies) and beech (Fagus sylvatica) trees. New Phytol. 1998, 138, 275–285. [Google Scholar] [CrossRef]
- Cooke, J.E.K.; Weih, M. Nitrogen storage and seasonal nitrogen cycling in Populus: Bridging molecular physiology and ecophysiology. New Phytol. 2005, 167, 19–30. [Google Scholar] [CrossRef]
- Wyka, T.P.; Zytkowiak, R.; Oleksyn, J. Seasonal dynamics of nitrogen level and gas exchange in different cohorts of Scots pine needles: A conflict between nitrogen mobilization and photosynthesis? Eur. J. For. Res. 2016, 135, 483–493. [Google Scholar] [CrossRef]
- Koba, K.; Tokuchi, N.; Wada, E.; Nakajima, T.; Iwatsubo, G. Intermittent denitrification: The application of a 15N natural abundance method to a forested ecosystem. Gerchimica Cosmochim. Acta 1997, 61, 5043–5050. [Google Scholar] [CrossRef]
- Koba, K.; Tokuchi, N.; Yoshioka, T.; Hobbie, E.A.; Iwatsubo, G. Natural Abundance of Nitrogen-15 in a Forest Soil. Soil Sci. Soc. Am. J. 1998, 62, 778–781. [Google Scholar] [CrossRef]
- Shi, J.; Ohte, N.; Tokuchi, N.; Imamura, N.; Nagayama, M.; Oda, T.; Suzuki, M. Nitrate isotopic composition reveals nitrogen deposition and transformation dynamics along the canopy-soil continuum of a suburban forest in Japan. Rapid Commun. Mass Spectrom. 2014, 28, 2539–2549. [Google Scholar] [CrossRef]
- Zhou, W.-J.; Sha, L.-Q.; Schaefer, D.A.; Zhang, Y.-P.; Song, Q.-H.; Tan, Z.-H.; Deng, Y.; Deng, X.-B.; Guan, H.-L. Direct effects of litter decomposition on soil dissolved organic carbon and nitrogen in a tropical rainforest. Soil Biol. Biochem. 2014, 81, 255–258. [Google Scholar] [CrossRef]
- Rennenberg, H.; Dannenmann, M. Nitrogen nutrition of trees and temperate forests – the significance of nitrogen availability in pedosphere and atmosphere. Forests 2015, 6, 2820–2835. [Google Scholar] [CrossRef]
- Dannenmann, M.; Bimüller, C.; Gschwendtner, S.; Leberecht, M.; Tejedor, J.; Bilela, S.; Gasche, R.; Haewinkel, M.; Baltensweiler, A.; Kögel-Knabner, I.; et al. Climate change impairs nitrogen cycling in European beech forests. PLoS ONE 2016, 11, e0158823. [Google Scholar] [CrossRef]
- Högberg, P.; Johannisson, C.; Yarwood, S.; Callesen, I.; Näsholm, T.; Myrold, D.D.; Högberg, M.N. Recovery of ectomycorrhizal after ‘nitrogen saturation’ of a conifer forest. New Phytol. 2011, 189, 515–525. [Google Scholar] [CrossRef]
- Clemmensen, K.E.; Bahr, A.; Ovaskainen, O.; Dahlberg, A.; Ekblad, A.; Wallander, H.; Stenlid, J.; Finlay, R.D.; Wardle, D.A.; Lindahl, B.D. Roots and Associated Fungi Drive Long-Term Carbon Sequestration in Boreal Forests. Science 2013, 339, 1615–1618. [Google Scholar] [CrossRef]
- Finzi, A.C.; Berthrong, S.T. The uptake of amino acids by microbes and trees in three cold temperate forests. Ecology 2005, 86, 3345–3353. [Google Scholar] [CrossRef]
- Jones, D.L.; Shannon, D.; Murphy, D.V.; Farrar, J. Role of dissolved organic nitrogen (DON) in soil N cycling in grassland soils. Soil Biol. Biochem. 2004, 36, 749–756. [Google Scholar] [CrossRef]
- Roberts, P.; Jones, D.L. Microbial and plant uptake of free amino sugars in grassland soils. Soil Biol. Biochem. 2012, 49, 139–149. [Google Scholar] [CrossRef]
- Malaguti, D.; Millard, P.; Wendler, R.; Hepburn, A.; Tagliavini, M. Translocation of amino acids in the xylem of apple (Malus domestica Borkh.) trees in spring as a consequence of both N remobilization and root uptake. J. Exp. Bot. 2001, 52, 1665–1671. [Google Scholar]
- Miyashita, Y.; Dolferus, R.; Ismond, K.P.; Goog, A.G. Alanine aminotransferase catalyses the breakdown of alanine after hypoxia in Arabidopsis thaliana. Plant J. 2007, 49, 1108–1121. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef]
- Näsholm, T. Removal of nitrogen during needle senescence in Scots pine (Pinus sylvestris L.). Oecologia 1994, 99, 290–296. [Google Scholar] [CrossRef]
- Näsholm, T.; Ericsson, A. Seasonal changes in amino acids, protein and total nitrogen in needles of fertilized Scots pine trees. Tree Physiol. 1990, 6, 267–281. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Coordinates | m.a.s.l. | Tree | DBH | Height | Age |
|---|---|---|---|---|---|---|
| B 1 | N38° 33.390’ E139° 52.630’ | 670 | A | 30.7 | 25.4 | 71 |
| B | 30.2 | 24.4 | 71 | |||
| C | 17.0 | 27.5 | 73 | |||
| B 2 | N38° 33.260’ E139° 52.570’ | 730 | A | 25.8 | 35.2 | 75 |
| B | 31.4 | 29.1 | 78 | |||
| C | 29.9 | 43.2 | 73 | |||
| B 3 | N38° 33.310’ E139° 52.600’ | 740 | A | 17.0 | 28.1 | 80 |
| B | 25.8 | 23.0 | 78 | |||
| C | 26.0 | 31.8 | 76 |
| Depth [cm] | pH [-] | Total N [g N kg−1] | TNb [g N kg−1] | Ammonium [g N kg−1] | Nitrate [g N kg−1] | DON [g N kg−1] | C : N |
|---|---|---|---|---|---|---|---|
| 0–5 | 3.6 ± 0.1 | 7.8 ± 2.1 | 0.29 ± 0.08 | 0.25 ± 0.05 | 0.004 ± 0.006 | 0.04 ± 0.03 | 20 ± 2 |
| 5–15 | 3.7 ± 0.3 | 3.6 ± 0.9 | 0.17 ± 0.03 | 0.10 ± 0.07 | 0.003 ± 0.005 | 0.07 ± 0.07 | 19 ± 1 |
| 15–30 | 3.8 ± 0.2 | 1.0 ± 0.2 | 0.14 ± 0.05 | 0.10 ± 0.08 | 0.003 ± 0.004 | 0.03 ± 0.03 | 15 ± 1 |
| Tissue | Period | Alanine [g N kg−1] | Arginine [g N kg−1] | Asparagine [g N kg−1] |
|---|---|---|---|---|
| Perennial roots | shoot growth | 0.34 ± 0.06 | 0.15 ± 0.10 | 2.2 ± 0.03 |
| green leaf | 0.06 ± 0.02 | 0.06 ± 0.03 | 0.76 ± 0.09 | |
| pre-abscission | 0.05 ± < 0.01 | 0.03 ± 0.01 | 0.24 ± 0.10 | |
| post abscission | 0.18 ± 0.05 | 0.13 ± 0.08 | 0.88 ± 0.021 | |
| Sapwood | shoot growth | 0.03 ± 0.10 | 0.00 ± 0.00 | 0.02 ± 0.01 |
| green leaf | 0.02 ± 0.01 | 0.00 ± 0.00 | 0.03 ± 0.01 | |
| pre-abscission | 0.06 ± 0.01 | 0.00 ± 0.00 | 0.04 ± 0.02 | |
| post abscission | 0.06 ± 0.01 | 0.01 ± <0.01 | 0.07 ± 0.02 | |
| leaf | shoot growth | 0.63 ± 0.28 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| green leaf | 0.05 ± 0.02 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| pre-abscission | 1.02 ± <0.01 | 0.01 ± <0.01 | 0.14 ± 0.01 | |
| Litter | post abscission | 0.02 ± <0.01 | 0.01 ± <0.01 | 0.09 ± 0.05 |
| Tissue | Amino Acid | Estimate | R2 | Adjusted R2 | p |
|---|---|---|---|---|---|
| roots | Asparagine | 1.52 | 0.67 | 0.60 | 0.002** |
| Other 1 | −2.90 | 0.003** | |||
| sapwood | Alanine | −14.53 | 0.53 | 0.36 | 0.026* |
| Asparagine | 37.18 | 0.349 | |||
| Other 1 | 0.38 | 0.976 | |||
| leaves | Alanine | 1.56 | 0.76 | 0.73 | 0.013* |
| Other 1 | 0.87 | 0.001** |
| Tissue | Biomass [kg] | N [g N] | ||
|---|---|---|---|---|
| Green Leaf Stage | Pre-abscission Stage | Post-abscission Stage | ||
| roots | 117 ± 9 a | 679 ± 53 | 862 ± 119* | 893 ± 106* |
| sapwood | 362 ± 20 b | 245 ± 70 | 208 ± 84 | 339 ± 126* |
| branches | 68 ± 5 a | 54 ± 8 | 52 ± 5 | 83 ± 10* |
| leaves | 8.7 ± 0.1 c | 193 ± 20 | 144 ± 26* | 64 ± 6* |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seidel, F.; Lopez C., M.L.; Celi, L.; Bonifacio, E.; Oikawa, A.; Yamanaka, T. N Isotope Fractionation in Tree Tissues During N Reabsorption and Remobilization in Fagus crenata Blume. Forests 2019, 10, 330. https://doi.org/10.3390/f10040330
Seidel F, Lopez C. ML, Celi L, Bonifacio E, Oikawa A, Yamanaka T. N Isotope Fractionation in Tree Tissues During N Reabsorption and Remobilization in Fagus crenata Blume. Forests. 2019; 10(4):330. https://doi.org/10.3390/f10040330
Chicago/Turabian StyleSeidel, Felix, M. Larry Lopez C., Luisella Celi, Eleonora Bonifacio, Akira Oikawa, and Toshiro Yamanaka. 2019. "N Isotope Fractionation in Tree Tissues During N Reabsorption and Remobilization in Fagus crenata Blume" Forests 10, no. 4: 330. https://doi.org/10.3390/f10040330
APA StyleSeidel, F., Lopez C., M. L., Celi, L., Bonifacio, E., Oikawa, A., & Yamanaka, T. (2019). N Isotope Fractionation in Tree Tissues During N Reabsorption and Remobilization in Fagus crenata Blume. Forests, 10(4), 330. https://doi.org/10.3390/f10040330