Small Roots of Parashorea chinensis Wang Hsie Decompose Slower than Twigs

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
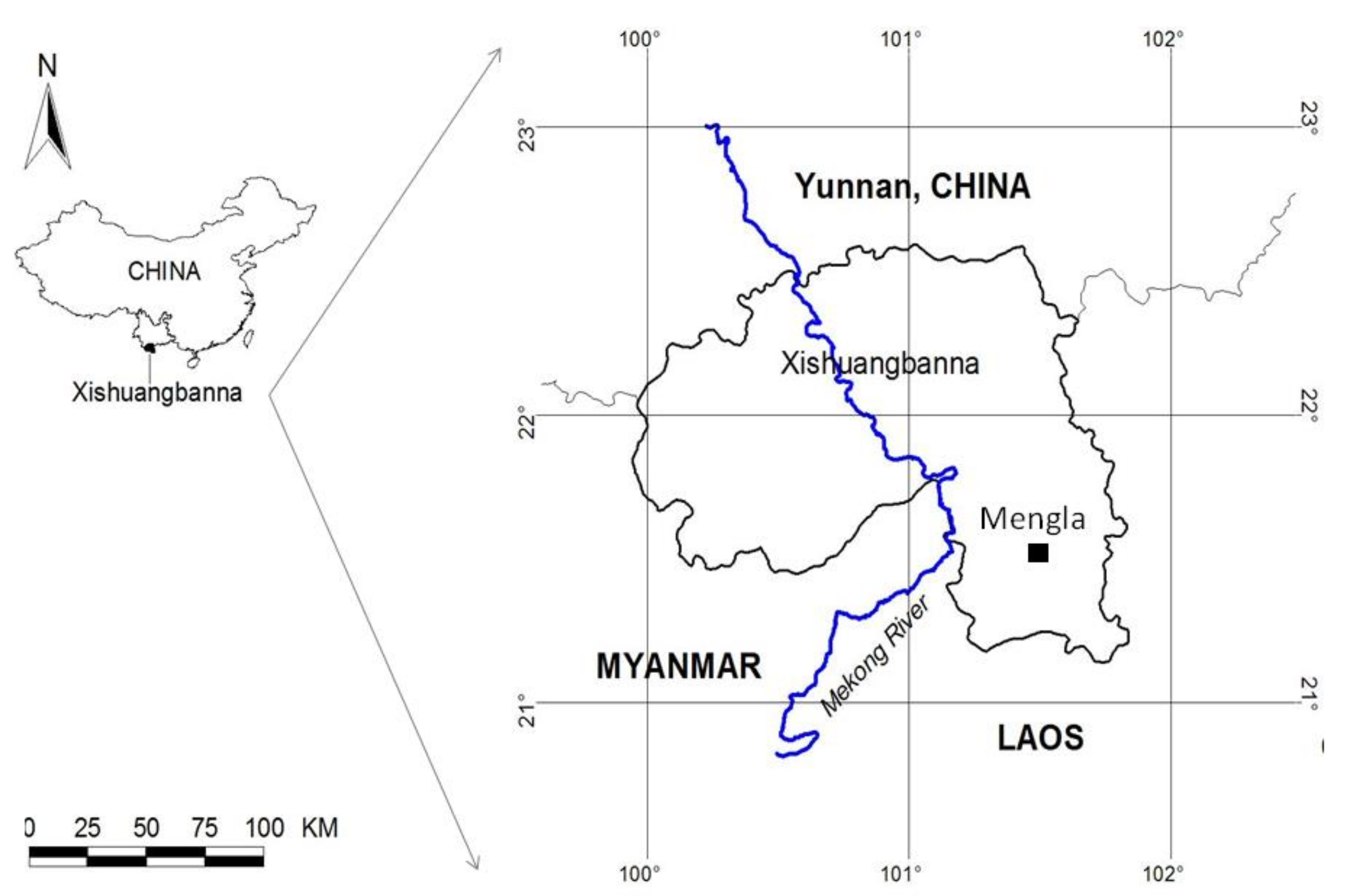
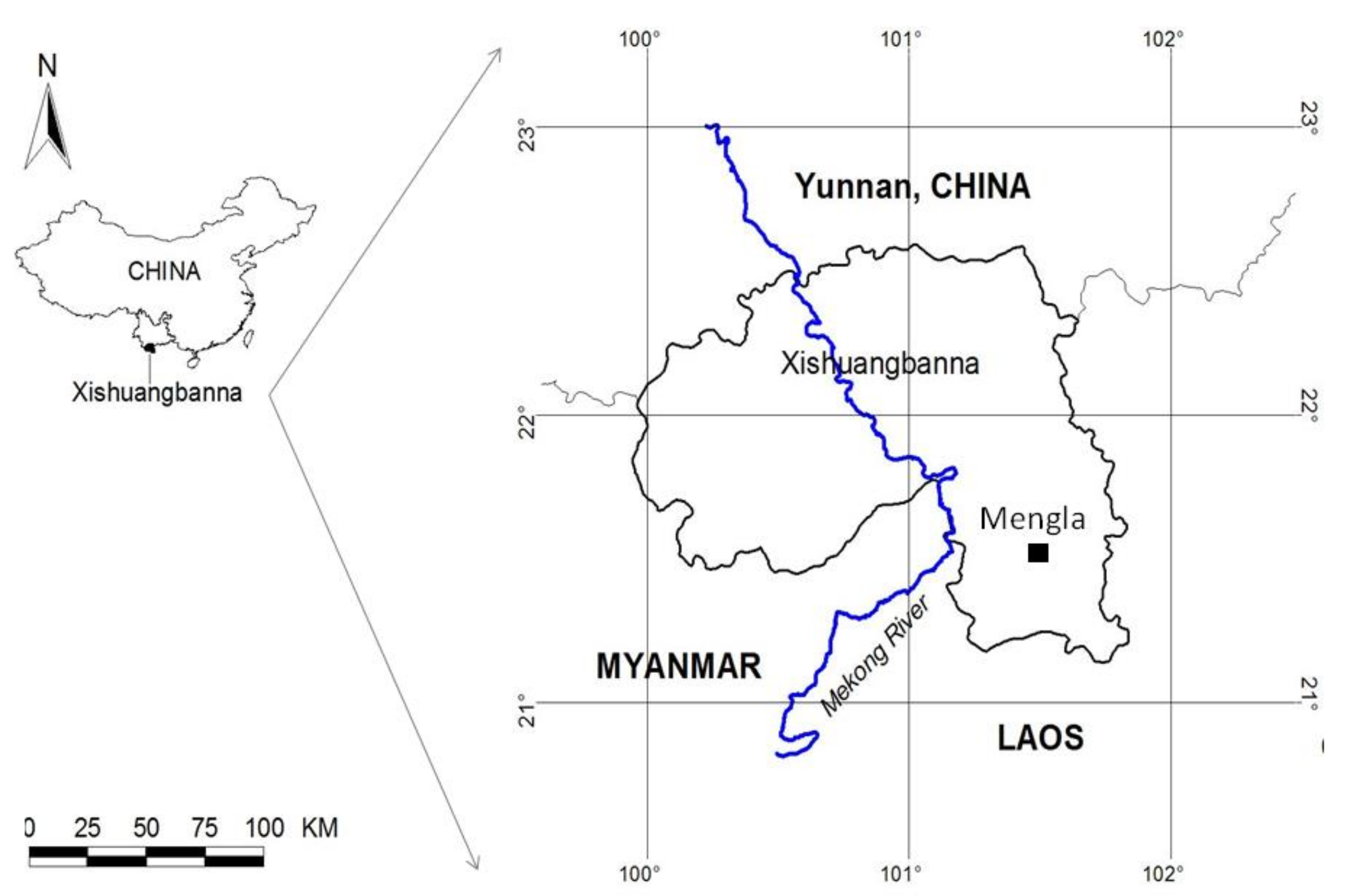
2.1. Study Site
2.2. Litter Materials and Litter Bag Preparation
2.3. Chemical Analysis
2.4. Data Analysis
3. Results
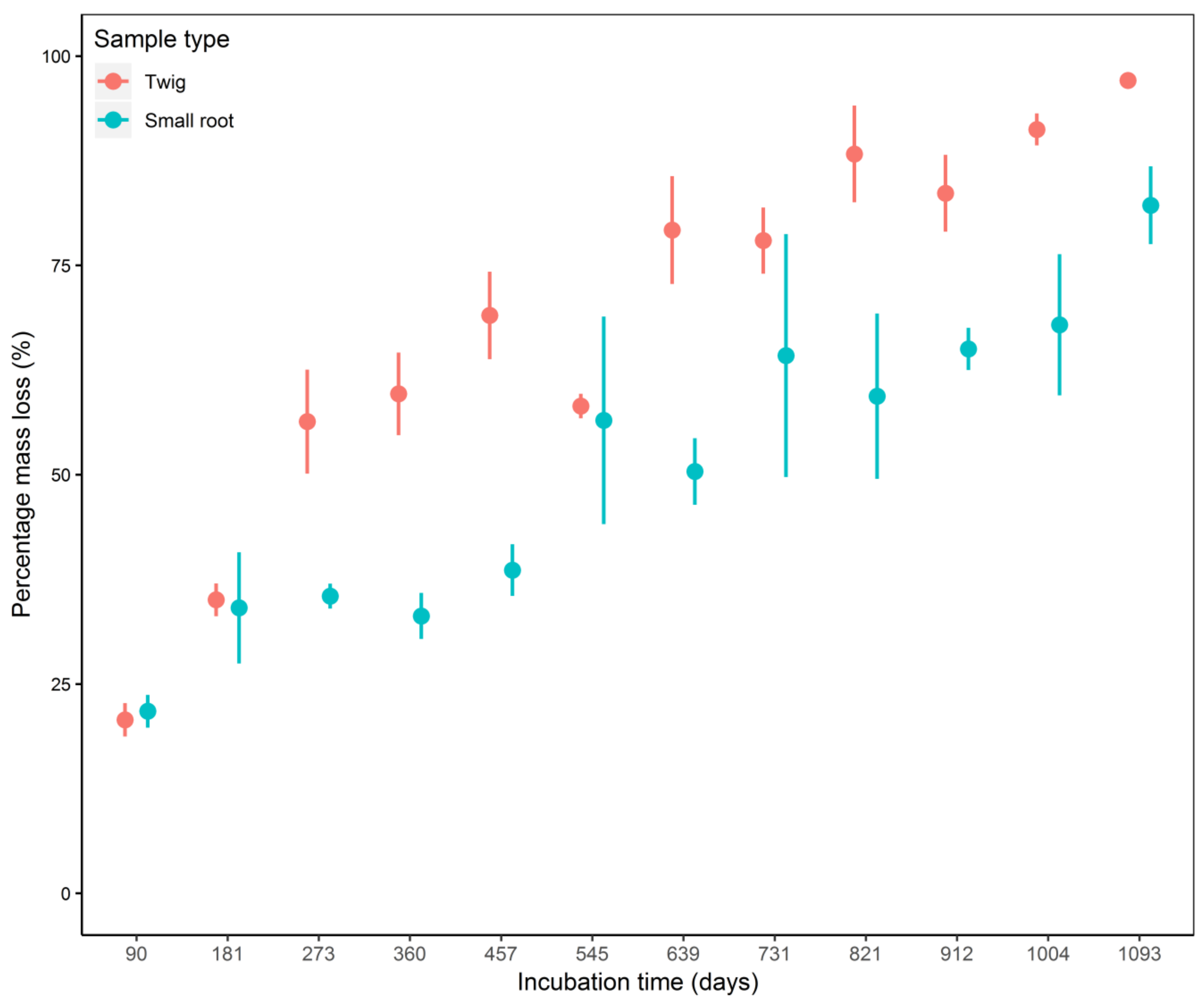
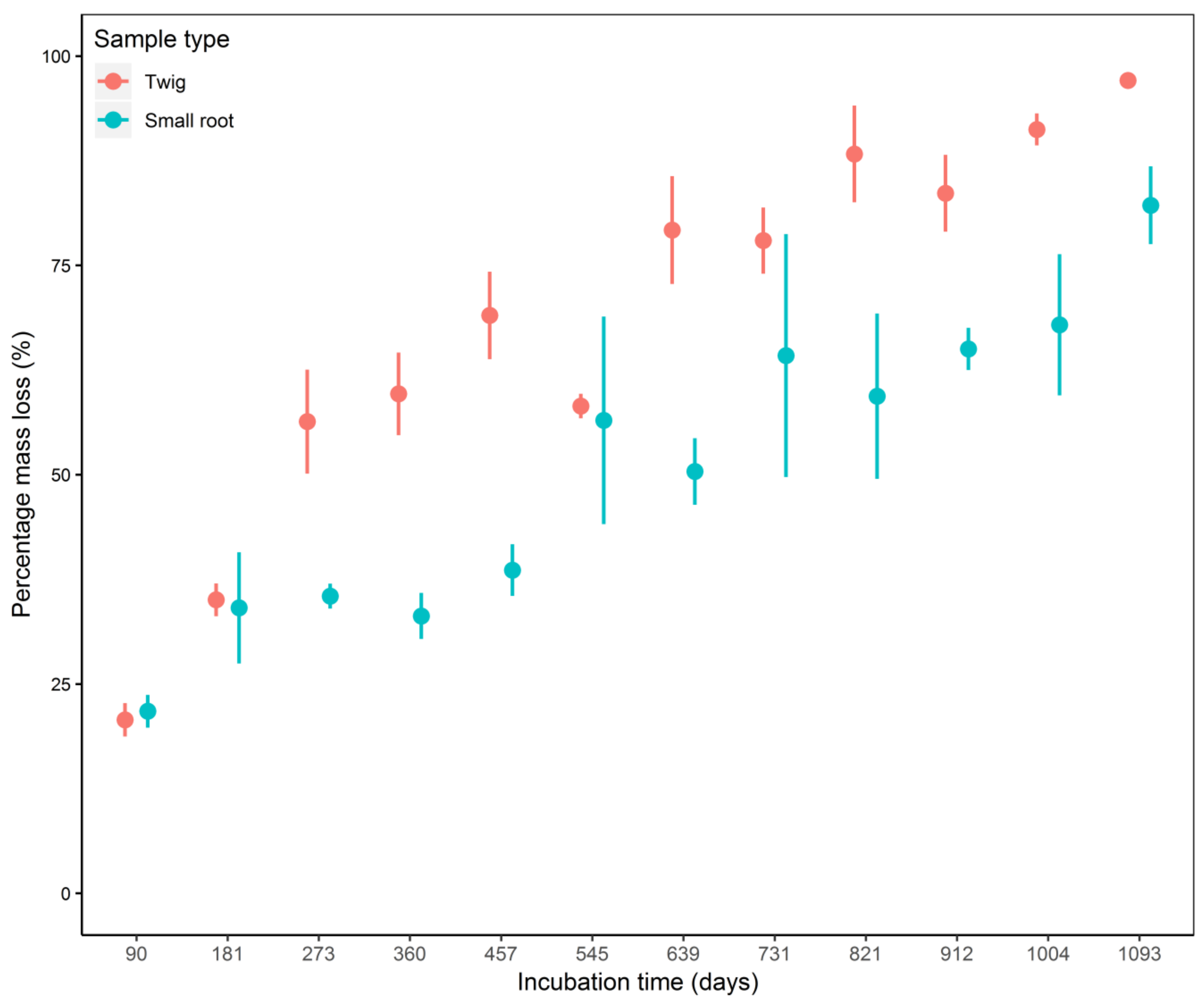
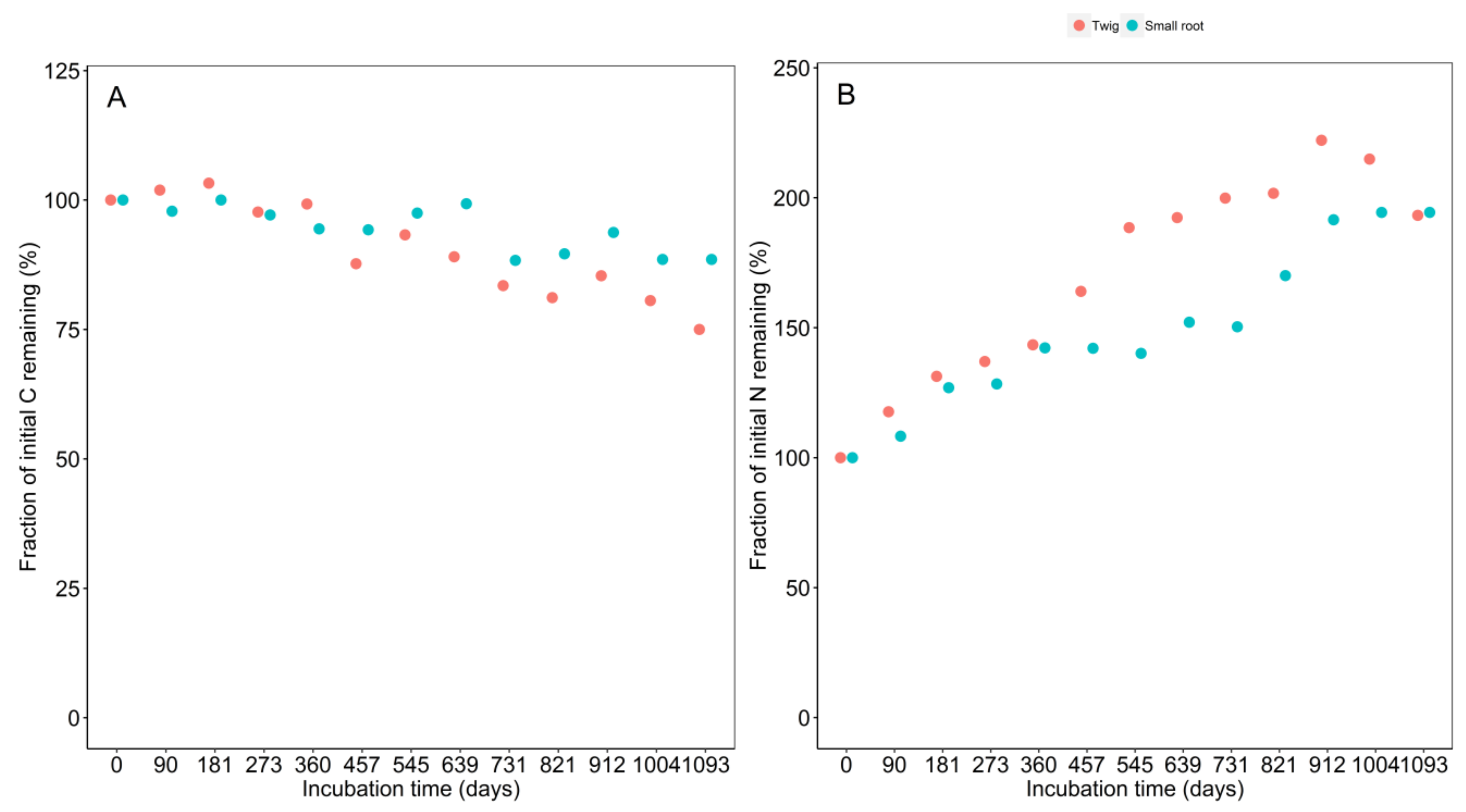
3.1. Decomposition Dynamics and Decay Rate
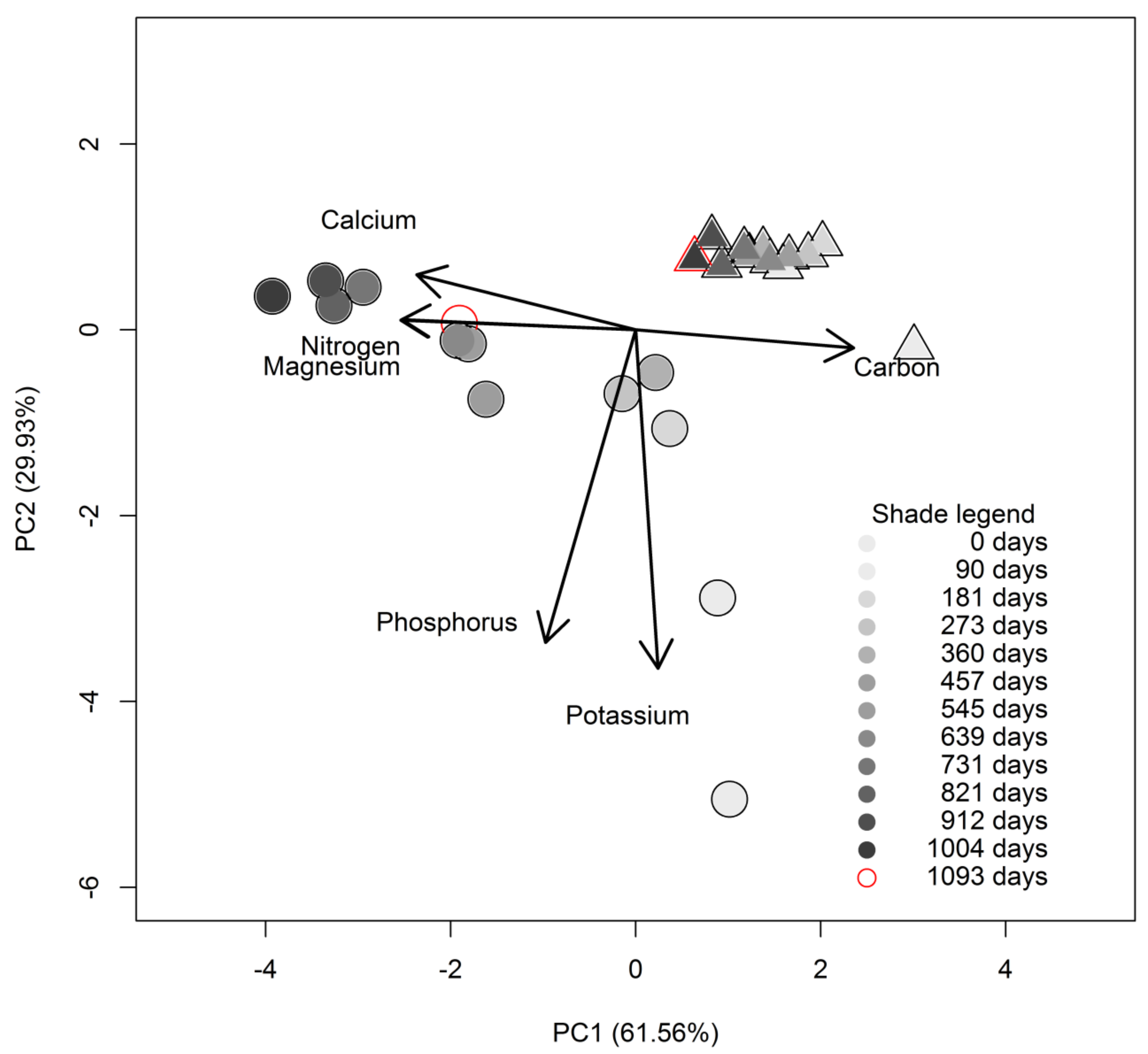
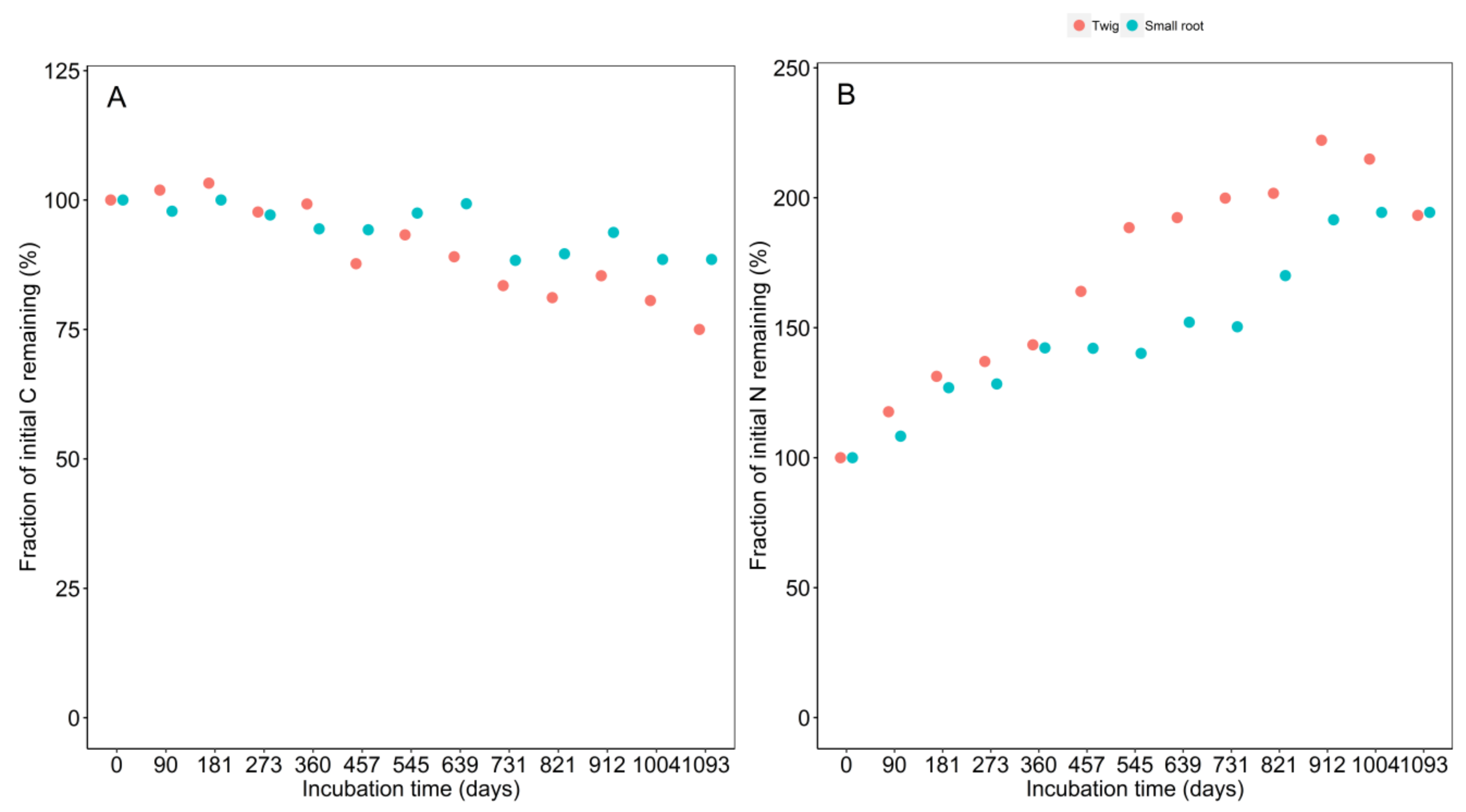
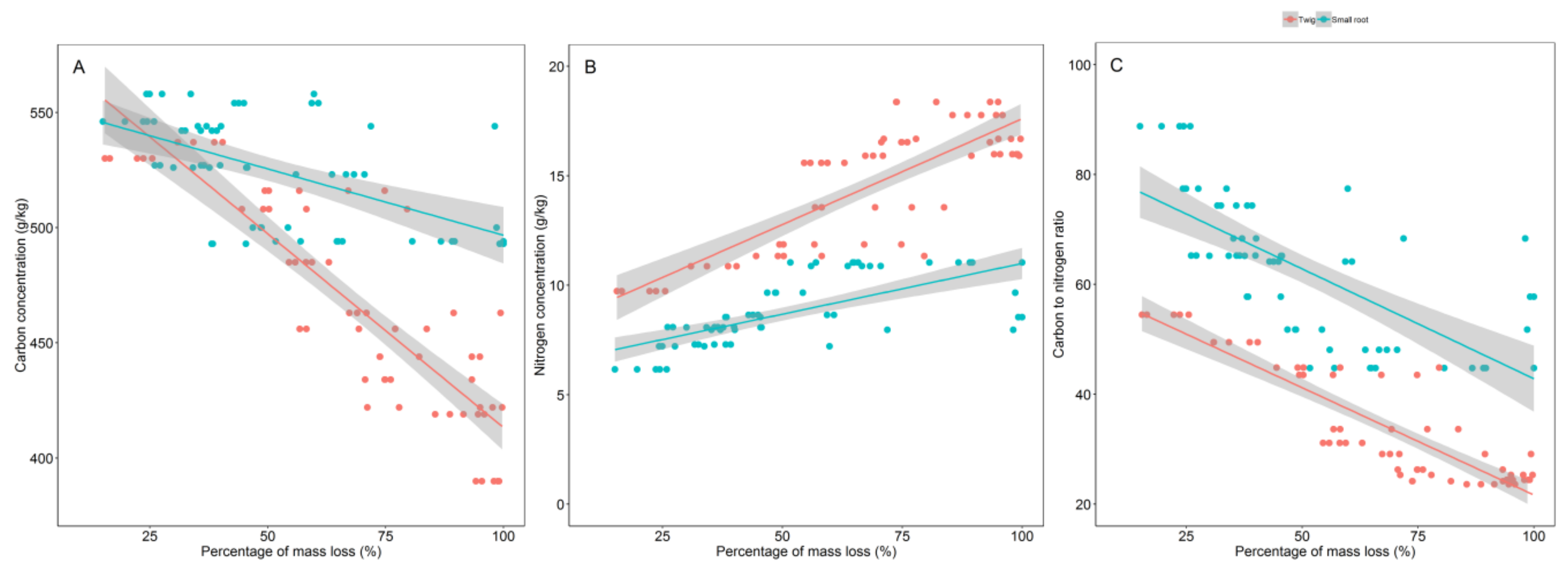
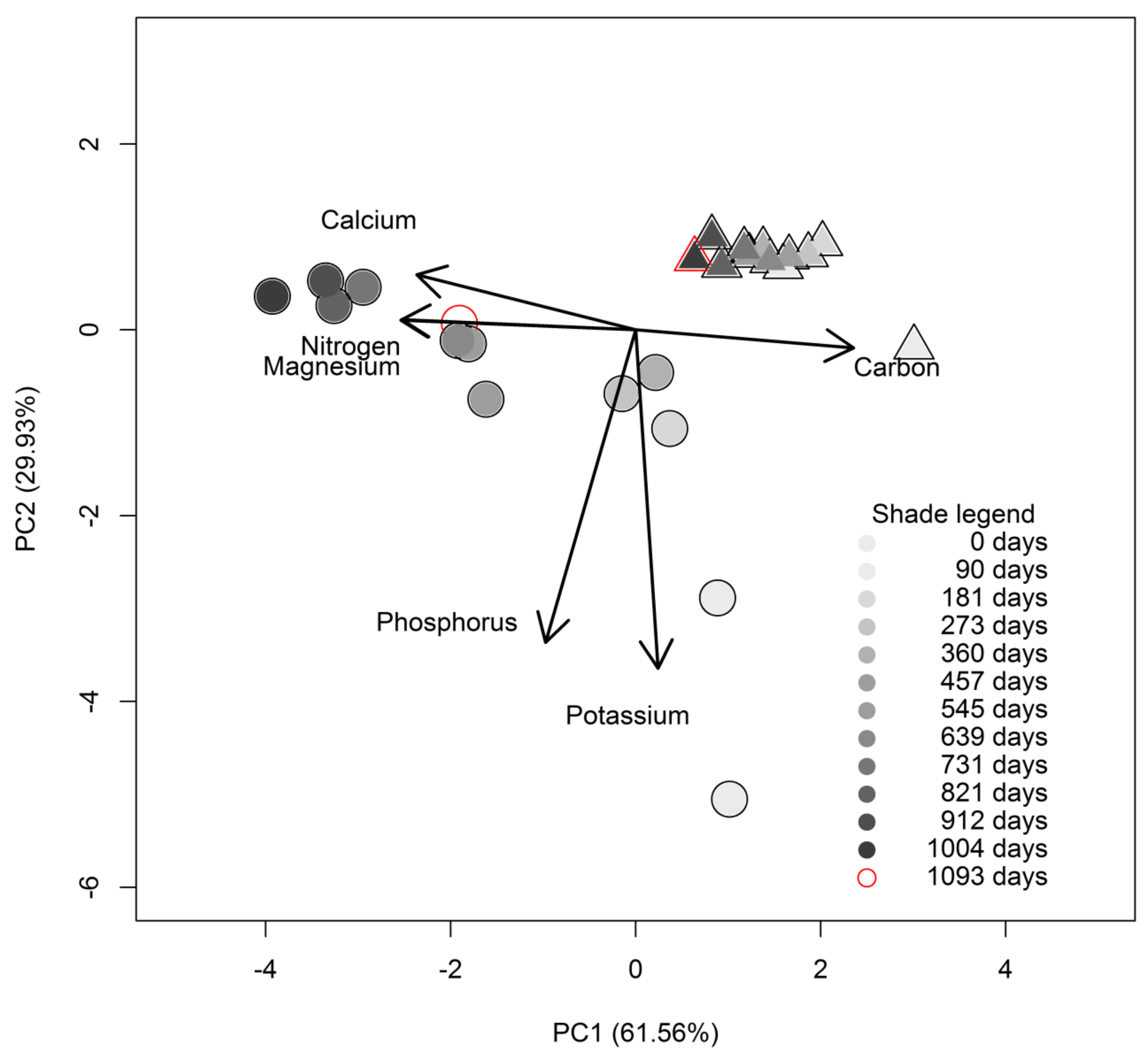
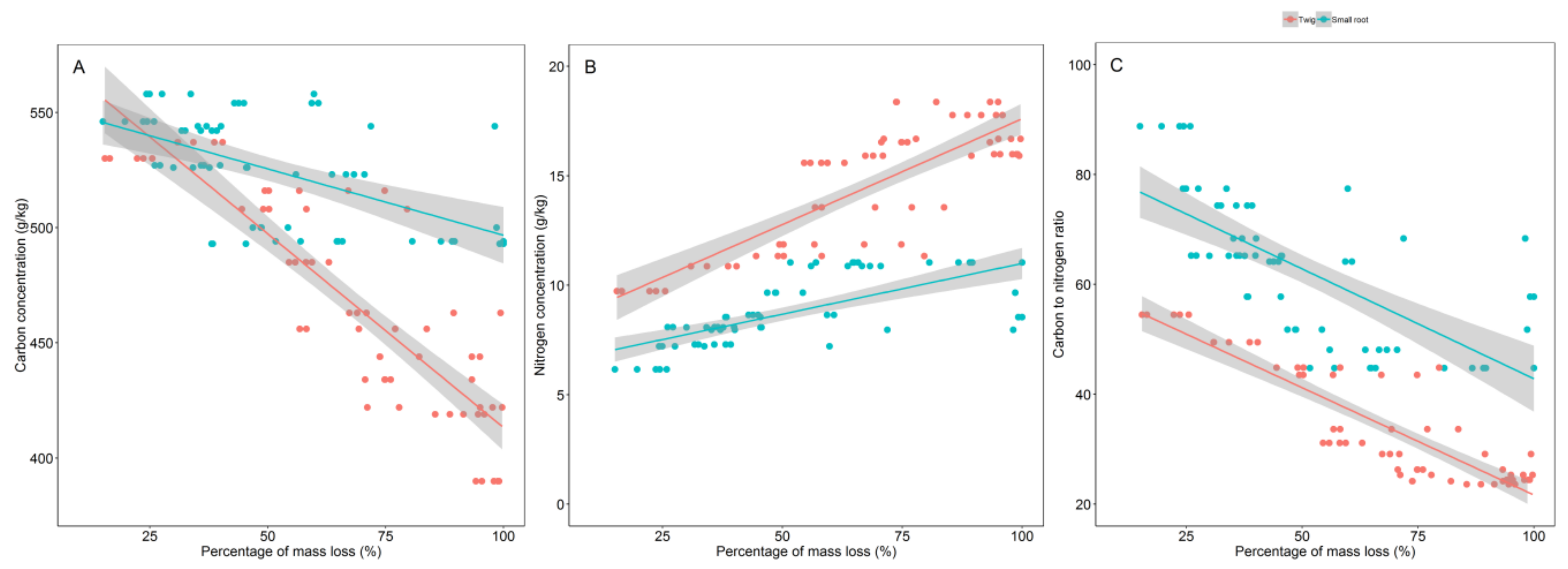
3.2. Initial Chemistry and Nutrient Dynamics during Decomposition
3.3. Relationship between Decomposition and C and N Concentrations Dynamics
4. Discussion
4.1. Decomposition Dynamics and Decay Rate
4.2. Initial Chemistry and Nutrients Dynamics during Decomposition
4.3. Relationship between Decomposition and C and N Dynamics
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lalnunzira, C.; Tripathi, S.K. Leaf and root production, decomposition and carbon and nitrogen fluxes during stand development in tropical moist forests, north-east India. Soil Res. 2018, 56, 306–317. [Google Scholar] [CrossRef]
- Ma, Z.; Guo, D.; Xu, X.; Lu, M.; Bardgett, R.D.; Eissenstat, D.M.; McCormack, M.L.; Hedin, L.O. Evolutionary history resolves global organization of root functional traits. Nature 2018, 555, 94–97. [Google Scholar] [CrossRef]
- Scheu, S.; Schauermann, J. Decomposition of roots and twigs: Effects of wood type (beech and ash), diameter, site of exposure and macrofauna exclusion. Plant Soil 1994, 163, 13–24. [Google Scholar] [CrossRef]
- Beidler, K.V.; Pritchard, S.G. Maintaining connectivity: Understanding the role of root order and mycelial networks in fine root decomposition of woody plants. Plant Soil 2017, 420, 19–36. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Cornelissen, J.H.C.; Amatangelo, K.; Dorrepaal, E.; Eviner, V.T.; Godoy, O.; Hobbie, S.E.; Hoorens, B.; Kurokawa, H.; Pérez-Harguindeguy, N.; et al. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol. Lett. 2008, 11, 1065–1071. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Sass-Klaassen, U.; Poorter, L.; van Geffen, K.; van Logtestijn, R.S.P.; van Hal, J.; Goudzwaard, L.; Sterck, F.J.; Klaassen, R.K.W.M.; Freschet, G.T.; et al. Controls on coarse wood decay in temperate tree species: Birth of the LOGLIFE experiment. Ambio 2012, 41 (Suppl. 3), 231–245. [Google Scholar] [CrossRef] [PubMed]
- Silver, W.L.; Miya, R.K. Global patterns in root decomposition: Comparisons of climate and litter quality effects. Oecologia 2001, 129, 407–419. [Google Scholar] [CrossRef]
- Luo, D.; Cheng, R.; Shi, Z.; Wang, W. Decomposition of leaves and fine roots in three subtropical plantations in China affected by litter substrate quality and soil microbial community. Forests 2017, 8, 412. [Google Scholar] [CrossRef]
- Sun, T.; Hobbie, S.E.; Berg, B.; Zhang, H.; Wang, Q.; Wang, Z.; Hättenschwiler, S. Contrasting dynamics and trait controls in first-order root compared with leaf litter decomposition. Proc. Natl. Acad. Sci. USA 2018, 115, 10392–10397. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Mao, Z.; Sun, T. Condensed tannin effects on decomposition of very fine roots among temperate tree species. Soil Biol. Biochem. 2016, 103, 489–492. [Google Scholar] [CrossRef]
- Zuo, J.; Hefting, M.M.; Berg, M.P.; van Logtestijn, R.S.P.; van Hal, J.; Goudzwaard, L.; Liu, J.C.; Sass-Klaassen, U.; Sterck, F.J.; Poorter, L.; et al. Is there a tree economics spectrum of decomposability? Soil Biol. Biochem. 2018, 119, 135–142. [Google Scholar] [CrossRef]
- Berg, B.; Mcclaugherty, C. Plant Litter: Decomposition, Humus Formation, Carbon Sequestration, 3rd ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA; Dordrecht, The Netherlands; London, UK, 2014; ISBN 9783642388200. [Google Scholar]
- Van Geffen, K.G.; Poorter, L.; Sass-Klaassen, U.; van Logtestijn, R.S.P.; Cornelissen, J.H.C. The trait contribution to wood decomposition rates of 15 Neotropical tree species. Ecology 2010, 91, 3686–3697. [Google Scholar] [CrossRef]
- Harmon, M.E.M.; Franklin, J.F.J.; Swanson, F.J.; Sollins, P.; Gregory, S.V.; Lattin, J.D.; Anderson, N.H.; Cline, S.P.; Aumen, N.G.; Sedell, J.R.; et al. Ecology of coarse woody debris in temperate ecosystems. Adv. Ecol. Res. 1986, 15, 133–302. [Google Scholar]
- Freschet, G.T.; Aerts, R.; Cornelissen, J.H.C. A plant economics spectrum of litter decomposability. Funct. Ecol. 2012, 26, 56–65. [Google Scholar] [CrossRef]
- Ichie, T.; Igarashi, S.; Yoshida, S.; Kenzo, T.; Masaki, T.; Tayasu, I. Are stored carbohydrates necessary for seed production in temperate deciduous trees? J. Ecol. 2013, 101, 525–531. [Google Scholar] [CrossRef]
- Eissenstat, D.M.; Wells, C.E.; Yanai, R.D.; Whitbeck, J.L. Building roots in a changing environment: Implications for root longevity. New Phytol. 2000, 147, 33–42. [Google Scholar] [CrossRef]
- Dossa, G.G.O.; Schaefer, D.; Zhang, J.-L.; Tao, J.-P.; Cao, K.-F.; Corlett, R.T.; Cunningham, A.B.; Xu, J.-C.; Cornelissen, J.H.C.; Harrison, R.D. The cover uncovered: Bark control over wood decomposition. J. Ecol. 2018, 106, 2147–2160. [Google Scholar] [CrossRef]
- Fan, P.; Guo, D. Slow decomposition of lower order roots: A key mechanism of root carbon and nutrient retention in the soil. Oecologia 2010, 163, 509–515. [Google Scholar] [CrossRef]
- Bloomfield, J.; Vogt, K.A.; Vogt, D.J. Decay rate and substrate quality of fine roots and foliage of two tropical tree species in the Luquillo Experimental Forest, Puerto Rico. Plant Soil 1993, 233–245. [Google Scholar] [CrossRef]
- Fujimaki, R.; Takeda, H.; Wiwatiwitaya, D. Fine root decomposition in tropical dry evergreen and dry deciduous forests in Thailand. J. For. Res. 2008, 13, 338–346. [Google Scholar] [CrossRef]
- Xiong, Y.; Fan, P.; Fu, S.; Zeng, H.; Guo, D. Slow decomposition and limited nitrogen release by lower order roots in eight Chinese temperate and subtropical trees. Plant Soil 2013, 363, 19–31. [Google Scholar] [CrossRef]
- Adam Langley, J.; Chapman, S.; Hungate, B.A. Ectomycorrhizal colonization slows root decomposition: The post-mortem fungal legacy. Ecol. Lett. 2006, 9, 955–959. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H. Research of community ecology on Shorea chinensis forest in Xishuangbanna. Acta Bot. Yunnanica 1992, 14, 237–258, (Chinese with English abstract). [Google Scholar]
- Cao, M.; Zhang, J.-H. Tree species diversity of tropical forest vegetation in Xishuangbanna, SW China. Biodivers. Conserv. 1997, 6, 995–1006. [Google Scholar] [CrossRef]
- Swift, M.J.; Heal, O.W.; Anderson, J.M. Decomposition in Terrestrial Ecosystems; University of California Press: Berkeley, CA, USA, 1979. [Google Scholar]
- Bradford, M.A.; Tordoff, G.M.; Eggers, T.; Jones, T.H.; John, E.; Newington, J.E. Microbiota, Fauna, and Mesh Size Interactions in Litter Decomposition in litter fauna, and mesh size interactions decomposition. Oikos 2002, 99, 317–323. [Google Scholar] [CrossRef]
- R Foundation for Statistical Computing R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; ISBN 3900051070.
- Olson, J.S. Energy Storage and the Balance of Producers and Decomposers in Ecological Systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Weedon, J.T. Decomposition trajectories of diverse litter types: A model selection analysis. Methods Ecol. Evol. 2014, 5, 173–182. [Google Scholar] [CrossRef]
- Oberle, B.; Covey, K.R.; Dunham, K.M.; Hernandez, E.J.; Walton, M.L.; Young, D.F.; Zanne, A.E. Dissecting the Effects of Diameter on Wood Decay Emphasizes the Importance of Cross-Stem Conductivity in Fraxinus americana. Ecosystems 2018, 21, 85–97. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; Debroy, S.; Sarkar, D.; Team, R.D.C. nlme: Linear and Nonlinear Mixed Effects Models; R Package Version 3.1-101; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Warton, D.I.; Hui, F.K.C. The arcsine is asinine: The analysis of proportions in ecology. Ecology 2011, 92, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Dornbush, M.E.; Isenhart, T.M.; Raich, J.W. Quantitying fine-root decomposition: An alternative to burried litterbags. Ecology 2002, 83, 2985–2990. [Google Scholar] [CrossRef]
- Berg, B.; Mcclaugherty, C. Decomposition of Fine Root and Woody Litter. In Plant Litter; Springer: Berlin, Heidelberg, 2008. [Google Scholar]
- Goebel, M.; Hobbie, S.E.; Bulaj, B.; Zadworny, M.; Archibald, D.D.; Oleksyn, J.; Reich, P.B.; Eissenstat, D.M. Decomposition of the finest root branching orders: Linking belowground dynamics to fine-root function and structure. Ecol. Monogr. 2011, 81, 89–102. [Google Scholar] [CrossRef]
- Mladkova, P.; Mladek, J.; Hejduk, S.; Hejcman, M.; Pakeman, R.J. Calcium plus magnesium indicates digestibility: The significance of the second major axis of plant chemical variation for ecological processes. Ecol. Lett. 2018, 21, 885–895. [Google Scholar] [CrossRef]
- García-Palacios, P.; Mckie, B.G.; Handa, I.T.; Frainer, A.; Hättenschwiler, S. The importance of litter traits and decomposers for litter decomposition: A comparison of aquatic and terrestrial ecosystems within and across biomes. Funct. Ecol. 2015, 30, 819–829. [Google Scholar] [CrossRef]
- Garrett, L.G.; Kimberley, M.O.; Oliver, G.R.; Pearce, S.H.; Beets, P.N. Decomposition of coarse woody roots and branches in managed Pinus radiata plantations in New Zealand—A time series approach. For. Ecol. Manag. 2012, 269, 116–123. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Estimate | Standard Error | t-Value | p-Value |
|---|---|---|---|---|
| (Intercept) | −0.769 | 0.142 | −5.406 | 0.000 |
| Incubation time | 0.002 | 0.000 | 11.369 | 0.000 |
| Small root | −0.229 | 0.201 | −1.140 | 0.256 |
| Incubation time: Small root | −0.001 | 0.000 | −2.067 | 0.041 |
| Incubation Time (Days) | Twig | Root | Twig | Root | Twig | Root | Twig | Root | Twig | Root | Twig | Root | Twig | Root |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C (g per kg) | N (g per kg) | P (g per kg) | K (g per kg) | Ca (g per kg) | Mg (g per kg) | C to N Ratio | ||||||||
| 0 | 520 | 558 | 8.27 | 5.68 | 1.96 | 0.33 | 8.72 | 3.4 | 11.77 | 8.37 | 1.51 | 1.2 | 62.88 | 98.24 |
| 90 | 530 | 546 | 9.73 | 6.15 | 1.1 | 0.3 | 7.28 | 1.91 | 14.6 | 16 | 1.71 | 1.78 | 54.47 | 88.78 |
| 181 | 537 | 558 | 10.86 | 7.21 | 0.87 | 0.3 | 3.91 | 1.1 | 16.97 | 13.26 | 2 | 1.53 | 49.45 | 77.39 |
| 273 | 508 | 542 | 11.33 | 7.29 | 0.9 | 0.33 | 2.97 | 1.32 | 17.92 | 13.68 | 2.04 | 1.45 | 44.84 | 74.35 |
| 360 | 516 | 527 | 11.86 | 8.08 | 0.95 | 0.34 | 2.05 | 1.26 | 16.3 | 15.11 | 1.74 | 1.56 | 43.51 | 65.22 |
| 457 | 456 | 526 | 13.56 | 8.07 | 0.93 | 0.34 | 3.28 | 1.52 | 19.65 | 16.57 | 2.63 | 1.57 | 33.63 | 65.18 |
| 545 | 485 | 544 | 15.59 | 7.96 | 0.86 | 0.3 | 2.18 | 1.57 | 21.39 | 14.42 | 2.69 | 1.56 | 31.11 | 68.34 |
| 639 | 463 | 554 | 15.91 | 8.64 | 0.88 | 0.37 | 2.03 | 1.44 | 21.31 | 15.82 | 2.5 | 1.61 | 29.10 | 64.12 |
| 731 | 434 | 493 | 16.53 | 8.54 | 0.77 | 0.282 | 1.47 | 1.47 | 25.96 | 12.86 | 2.81 | 1.63 | 26.26 | 57.73 |
| 821 | 422 | 500 | 16.68 | 9.66 | 0.71 | 0.36 | 2.21 | 1.59 | 24.53 | 13.22 | 3.25 | 1.74 | 25.30 | 51.76 |
| 912 | 444 | 523 | 18.37 | 10.88 | 0.75 | 0.36 | 1.48 | 1.05 | 27.71 | 17.5 | 2.93 | 1.44 | 24.17 | 48.07 |
| 1004 | 419 | 494 | 17.77 | 11.04 | 0.82 | 0.37 | 1.72 | 1.49 | 28.08 | 14.74 | 3.35 | 1.62 | 23.58 | 44.75 |
| 1093 | 390 | 501 | 15.98 | 9.37 | 0.65 | 0.32 | 2.35 | 1.63 | 14.98 | 11.5 | 2.48 | 1.83 | 24.41 | 53.47 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dossa, G.G.O.; Jin, Y.-Q.; Lü, X.-T.; Tang, J.-W.; Harrison, R.D. Small Roots of Parashorea chinensis Wang Hsie Decompose Slower than Twigs. Forests 2019, 10, 301. https://doi.org/10.3390/f10040301
Dossa GGO, Jin Y-Q, Lü X-T, Tang J-W, Harrison RD. Small Roots of Parashorea chinensis Wang Hsie Decompose Slower than Twigs. Forests. 2019; 10(4):301. https://doi.org/10.3390/f10040301
Chicago/Turabian StyleDossa, Gbadamassi G. O., Yan-Qiang Jin, Xiao-Tao Lü, Jian-Wei Tang, and Rhett D. Harrison. 2019. "Small Roots of Parashorea chinensis Wang Hsie Decompose Slower than Twigs" Forests 10, no. 4: 301. https://doi.org/10.3390/f10040301
APA StyleDossa, G. G. O., Jin, Y.-Q., Lü, X.-T., Tang, J.-W., & Harrison, R. D. (2019). Small Roots of Parashorea chinensis Wang Hsie Decompose Slower than Twigs. Forests, 10(4), 301. https://doi.org/10.3390/f10040301