High Precision Altimeter Demonstrates Simplification and Depression of Microtopography on Seismic Lines in Treed Peatlands



Abstract
1. Introduction
2. Materials and Methods
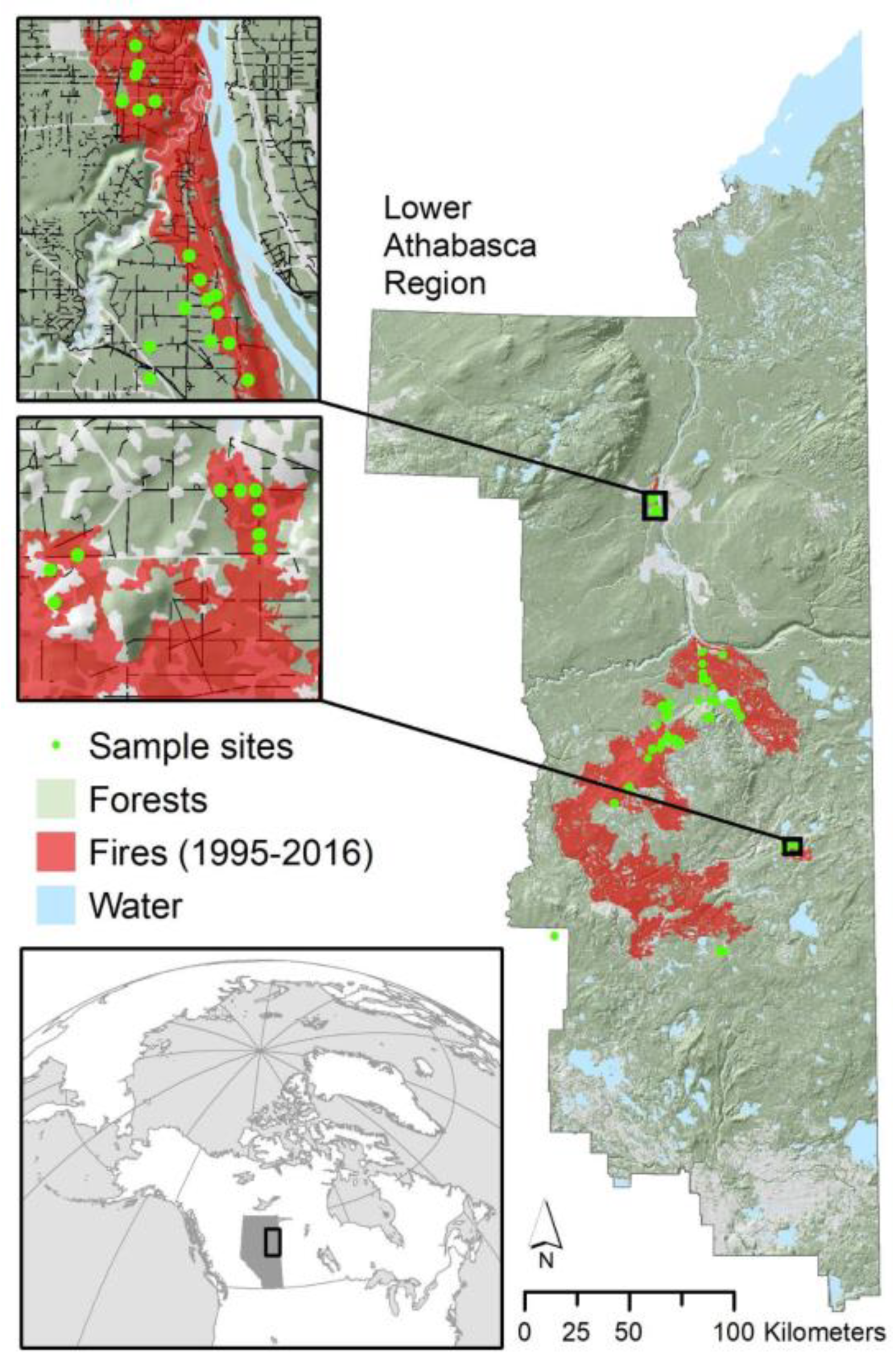
2.1. Study Area
2.2. Site Selection
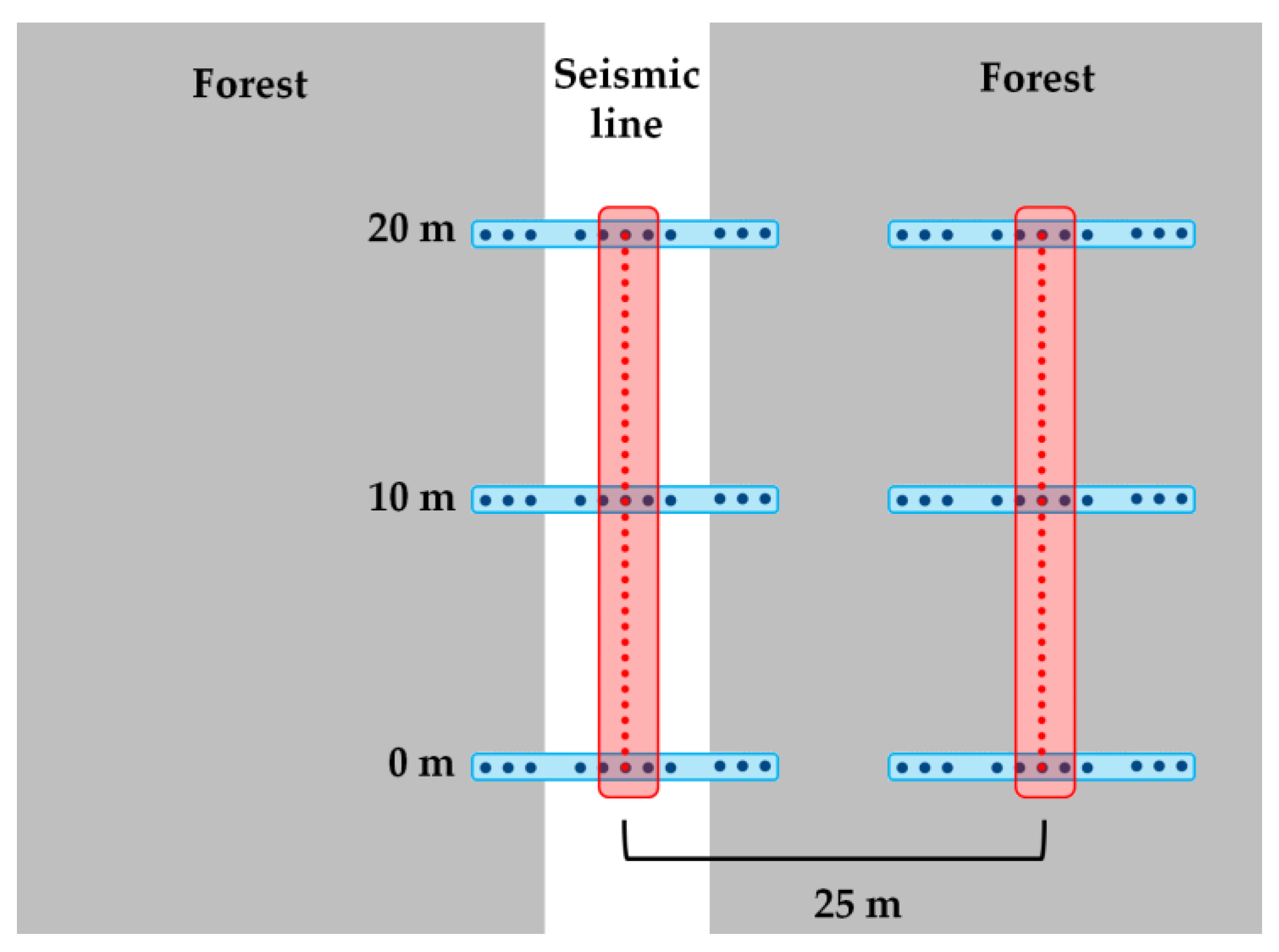
2.3. Field Measures of Microtopography
2.4. Data Analysis
3. Results
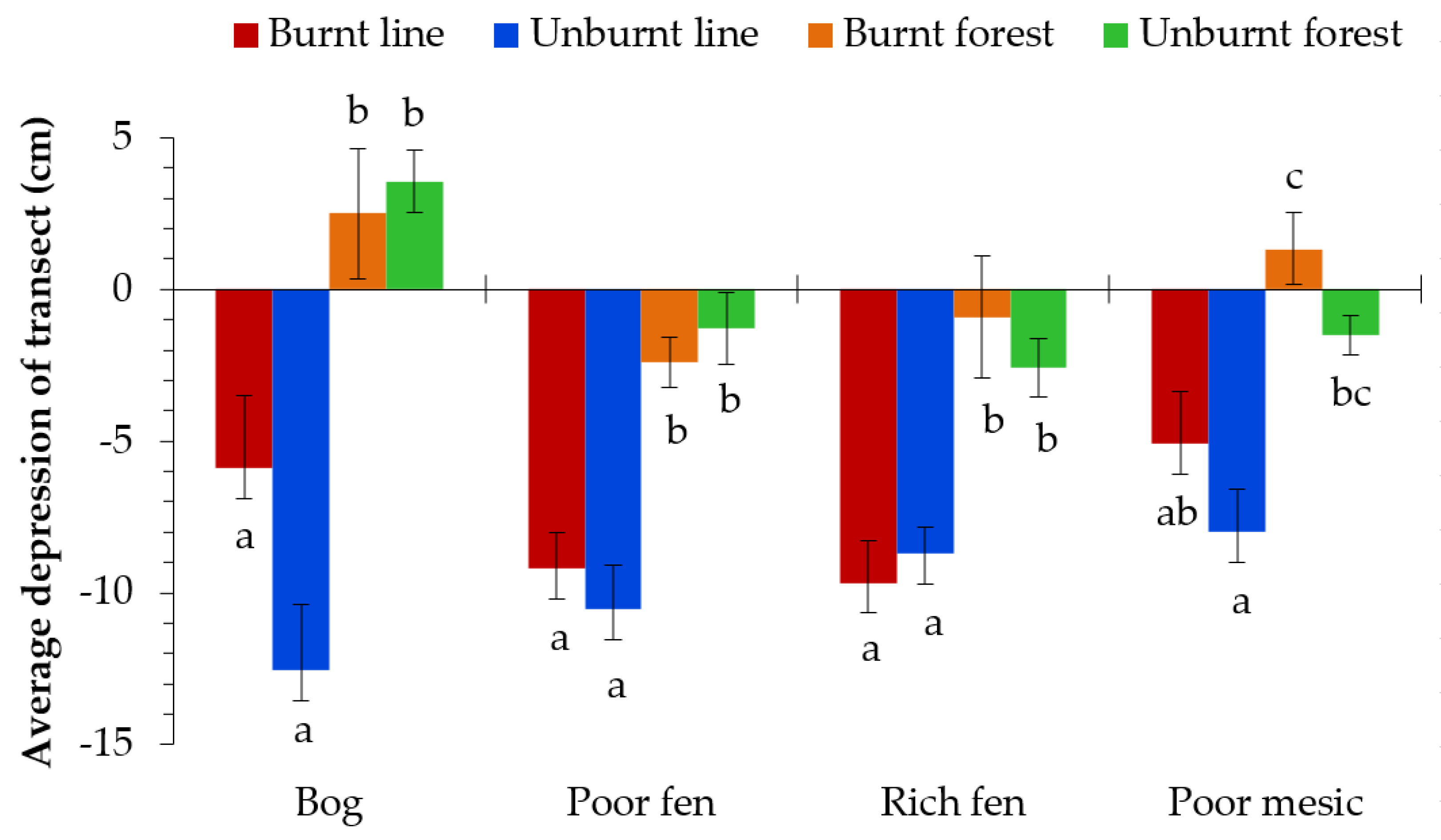
3.1. Seismic Line Depression
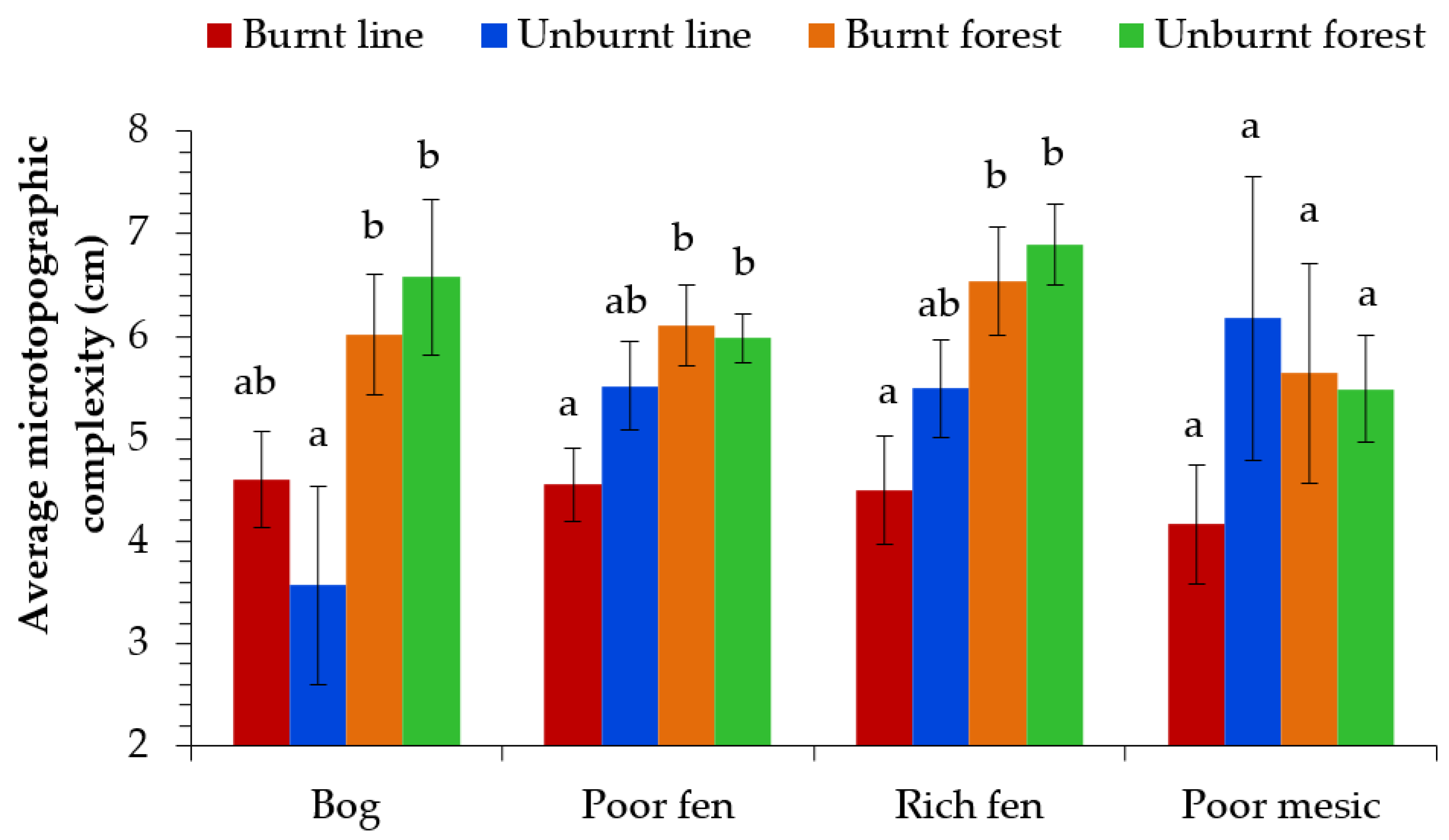
3.2. Seismic Line Simplification
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alexandratos, N. Counties with Rapid Population Growth and Resource Constraints: Issues of Food, Agriculture and Development. Popul. Dev. Rev. 2005, 31, 237–258. [Google Scholar] [CrossRef]
- Schneider, R.R. Alternative futures: Alberta’s boreal forest at the crossroads; Federation of Alberta Naturalists: Edmonton, AB, Canada, 2002. [Google Scholar]
- Lee, P.; Boutin, S. Persistence and developmental transitions of wide seismic lines in the western Boreal Plains of Canada. J. Environ. Manag. 2006, 78, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Veblen, T.T.; Kitzberger, T.; Donnegan, J. Climatic and human influences on fire regimes in ponderosa pine forests in the Colorado. Ecol. Appl. 2001, 10, 1178–1195. [Google Scholar] [CrossRef]
- Lefort, P.; Gauthier, S.; Bergeron, Y. The influence of fire weather and land use on fire activity of the Lake Abitibi area, Eastern Canada. Forest Sci. 2003, 49, 509–521. [Google Scholar]
- Arienti, M.C.; Cumming, S.G.; Krawchuk, M.A.; Boutin, S. Road network density correlated with increased lightning fire incidence in the Canadian western boreal forest. Int. J. Wildland Fire 2009, 19, 970–982. [Google Scholar] [CrossRef]
- van Rensen, C.K.; Nielsen, S.E.; White, B.; Vinge, R.; Lieffers, V.J. Natural regeneration of forest vegetation on legacy seismic lines in boreal habitats in Alberta’s oil sands region. Biol. Conserv. 2015, 184, 127–135. [Google Scholar] [CrossRef]
- Government of Alberta: Natural Regions Committee. Natural Regions and Subregions of Alberta; Government of Alberta: Edmonton, AB, Canada, 2006. Available online: https://www.albertaparks.ca/media/2942026/nrsrcomplete_may_06.pdf (accessed on 16 March 2019).
- Braithwaite, N.T.; Mallik, A.U. Edge effects of wildfire and riparian buffers along boreal forest streams. J. Appl. Ecol. 2012, 49, 192–201. [Google Scholar] [CrossRef]
- Laurance, W.F.; Nascimento, H.E.M.; Laurance, S.G.; Andrade, A.; Ewers, R.M.; Harms, K.E.; Luizao, R.C.C.; Ribeiro, J.E. Habitat Fragmentation, Variable Edge Effects, and the Landscape-Divergence Hypothesis. PLoS ONE 2007, 2, 2–8. [Google Scholar] [CrossRef]
- Filicetti, A.T.; Cody, M.; Nielsen, S.E. Caribou Conservation: Restoring Trees on Seismic Lines in Alberta, Canada. Forests 2019, 10, 185. [Google Scholar] [CrossRef]
- Vitt, D.H. An overview of factors that influence the development of Canadian peatlands. The Memoirs of the Entomological Society of Canada 1994, 126, 7–20. [Google Scholar] [CrossRef]
- Chen, J.; Budisulistiorini, S.H.; Itoh, M.; Lee, W.; Mijakawa, T.; Komasaki, Y.; Yang, L.D.Q.; Kuwata, M. Water uptake by fresh Indonesian peat burning particles is limited by water-soluble organic matter. Atmos. Chem. Phys. 2017, 17, 11591–11604. [Google Scholar] [CrossRef]
- Lovitt, J.; Rahman, M.M.; Saraswati, S.; McDermid, G.J.; Strack, M.; Xu, B. UAV Remote Sensing Can Reveal the Effects of Low-Impact Seismic Lines on Surface Morphology, Hydrology, and Methane (CH4) Release in a Boreal Treed Bog. JGR Biogeosci. 2018, 123, 741–1142. [Google Scholar] [CrossRef]
- Tigner, J.; Bayne, E.M.; Boutin, S. Black bear use of seismic lines in Northern Canada. J. Wildl. Manag. 2014, 78, 282–292. [Google Scholar] [CrossRef]
- Linke, J.; Franklin, S.E.; Huettman, F.; Stenhouse, G.B. Seismic Cutlines, Changing Landscape Metrics and Grizzly Bear Landscape use in Alberta. Landsc. Ecol. 2005, 20, 811–826. [Google Scholar] [CrossRef]
- Tigner, J.; Bayne, E.M.; Boutin, S. American Marten Respond to Seismic Lines in Northern Canada at Two Spatial Scales. PLoS ONE 2015, 10, e0118720. [Google Scholar] [CrossRef] [PubMed]
- Bayne, E.M.; Boutin, S.; Tracz, B.; Charest, K. Functional and numerical responses of ovenbirds (Seiurus aurocapilla) to changing seismic exploration practices in Alberta’s boreal forest. Ecoscience 2005, 12, 216–222. [Google Scholar] [CrossRef]
- Riva, F.; Acorn, J.H.; Nielsen, S.E. Localized disturbances from oil sands developments increase butterfly diversity and abundance in Alberta’s boreal forests. Biol. Conserv. 2018, 217, 173–180. [Google Scholar] [CrossRef]
- Environment Canada. Species at Risk Act Recovery Strategy Series; Environment Canada: Ottawa, ON, Canada, 2012.
- Dyer, S.J.; O’Neill, J.P.; Wasel, S.M.; Boutin, S. Avoidance of Industrial Development by Woodland Caribou. J. Wildl. Manag. 2001, 65, 531–542. [Google Scholar] [CrossRef]
- James, A.R.C.; Stuart-Smith, A.K. Distribution of caribou and wolves in relation to linear corridors. J. Wildl. Manag. 2000, 64, 154–159. [Google Scholar] [CrossRef]
- Latham, A.D.M.; Latham, M.C.; Boyce, M.S.; Boutin, S. Movement responses by wolves to industrial linear features and their effect on woodland caribou in northeastern Alberta. Ecol. Appl. 2011, 21, 2854–2865. [Google Scholar] [CrossRef]
- Latham, A.D.M.; Latham, M.C.; McCutchen, N.A.; Boutin, S. Invading White-Tailed Deer Change Wolf–Caribou Dynamics in Northeastern Alberta. J. Wildl. Manag. 2011, 75, 204–212. [Google Scholar] [CrossRef]
- Government of Canada. Woodland Caribou (Rangifer Tarandus Caribou), Boreal Population: Recovery Strategy 2012; Government of Canada: Ottawa, ON, Canada, 2012. Available online: https://www.canada.ca/en/environment-climate-change/services/species-risk-public-registry/recovery-strategies/woodland-caribou-boreal-population–2012.html (accessed on 16 March 2019).
- Kuhry, P. The Role of Fire in the Development of Sphagnum-Dominated Peatlands in Western Boreal Canada. J. Ecol. 1994, 82, 899–910. [Google Scholar] [CrossRef]
- Caners, T.; Lieffers, V.J. Divergent Pathways of Successional Recovery for In Situ Oil Sands Exploration Drilling Pads on Wooded Moderate-Rich Fens in Alberta, Canada. J. Res. Ecol. 2014, 22, 657–667. [Google Scholar] [CrossRef]
- Lieffers, V.J.; Caners, R.T.; Hangfei, G. Re-establishment of hummock topography promotes tree regeneration on highly disturbed moderate-rich fens. J. Environ. Manag. 2017, 197, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Benscoter, B.W.; Greencare, D.; Turetsky, M.R. Wildfire as a key determinant of peatland microtopography. Can. J. For. Res. 2015, 45, 1132–1136. [Google Scholar] [CrossRef]
- Benscoter, B.W.; Wieder, R.K.; Vitt, D.H. Linking microtopography with post-fire succession in bogs. J. Veg. Sci. 2005, 16, 453–460. [Google Scholar] [CrossRef]
- Weltzin, J.F.; Harth, C.; Bridgham, S.D.; Pastor, J.; Vonderharr, M. Production and microtopography of bog bryophytes; response to warming and water-table manipulations. Oecol 2001, 128, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Triisberg, T.; Karofeld, E.; Liira, J.; Orru, M.; Ramst, R.; Paal, J. Microtopography and the Properties of Residual Peat Are Convenient Indicators for Restoration Planning of Abandoned Extracted Peatlands. Restor. Ecol. 2014, 2, 31–39. [Google Scholar] [CrossRef]
- Alberta Biological Monitoring Institute (ABMI). Terrestrial Field Data Collection Protocol (Abridged Version). Available online: http://www.abmi.ca/home/publications/1-50/46.html (accessed on 15 May 2017).
- StataCorp. Stata Statistical Software: Release 15; StataCorp LLC: College Station, TX, USA, 2017. [Google Scholar]
- Uldis, S.; Bladon, K.D.; Kelly, E.N.; Esch, E.; Spence, K.R.; Stone, M.; Emelko, M.B.; Boon, S.; Wagner, M.J.; Williams, C.H.S.; Tichkowsky, I. Fire-year legacy of wildfire and salvage logging impacts on nutrient runoff and aquatic plant, invertebrate, and fish productivity. Ecohydrology 2014, 7, 1508–1523. [Google Scholar]
- Pouliot, R.; Rochefort, L.R.; Karofeld, E. Initiation of microtopography in re-vegetated cutover peatlands: evolution of plant species composition. Appl. Veg. Sci. 2019, 15, 369–382. [Google Scholar] [CrossRef]
- Brubaker, K.M.; Myers, W.L.; Drohan, P.L.; Miller, D.A. The Use of LiDAR Terrain Data in Characterizing Surface Roughness and Microtopography. Appl. Environ. Soil Sci. 2013, 4, 1–13. [Google Scholar] [CrossRef]
- Franklin, S.E. Pixel-and object-based multispectral classification of forest tree species from small unmanned aerial vehicles. J. Unmanned Veh. Sys. 2018, 6, 195–211. [Google Scholar] [CrossRef]
- Feduck, C.; McDermid, G.J.; Castilla, G. Detection of Coniferous Seedlings in UAV Imagery. Forests 2018, 9, 432. [Google Scholar] [CrossRef]
- McDonald, T.P.; Fulton, J.P.; Darr, M.J.; Gallagher, T.V. Evaluation of a system to spatially monitor hand planting of pine seedlings. Comput. Electron. Agr. 2008, 64, 173–182. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total | Bog | Poor Fen | Rich Fen | Poor Mesic | Total |
|---|---|---|---|---|---|
| 15 | 46 | 18 | 23 | 102 | |
| Unburnt | 6 | 19 | 8 | 9 | 42 |
| Burnt | 9 | 27 | 10 | 14 | 60 |
| Fire Year | |||||
| 1995 | 3 | 7 | 2 | 0 | 12 |
| 1999 | 3 | 1 | 1 | 5 | 10 |
| 2009 | 3 | 5 | 0 | 6 | 14 |
| 2011 | 0 | 8 | 2 | 2 | 12 |
| 2016 | 0 | 6 | 5 | 1 | 12 |
| Statistic | Depression Depth (cm) | Microtopographic Complexity (cm) | ||
|---|---|---|---|---|
| Seismic Line | Adjacent Forest | Seismic Line | Adjacent Forest | |
| Mean | −8.42 | −0.54 | 4.85 | 6.08 |
| Standard error | 0.66 | 0.57 | 0.22 | 0.21 |
| t | 9.093 | 4.824 | ||
| df | 202 | 202 | ||
| p-value | <0.001 | <0.001 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stevenson, C.J.; Filicetti, A.T.; Nielsen, S.E. High Precision Altimeter Demonstrates Simplification and Depression of Microtopography on Seismic Lines in Treed Peatlands. Forests 2019, 10, 295. https://doi.org/10.3390/f10040295
Stevenson CJ, Filicetti AT, Nielsen SE. High Precision Altimeter Demonstrates Simplification and Depression of Microtopography on Seismic Lines in Treed Peatlands. Forests. 2019; 10(4):295. https://doi.org/10.3390/f10040295
Chicago/Turabian StyleStevenson, Cassondra J., Angelo T. Filicetti, and Scott E. Nielsen. 2019. "High Precision Altimeter Demonstrates Simplification and Depression of Microtopography on Seismic Lines in Treed Peatlands" Forests 10, no. 4: 295. https://doi.org/10.3390/f10040295
APA StyleStevenson, C. J., Filicetti, A. T., & Nielsen, S. E. (2019). High Precision Altimeter Demonstrates Simplification and Depression of Microtopography on Seismic Lines in Treed Peatlands. Forests, 10(4), 295. https://doi.org/10.3390/f10040295