Physical Properties of Seeds of Eleven Fir Species

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Physical Properties
2.3. Statistical Analysis
3. Results and Discussion
3.1. Experimental Material
3.2. Correlations Between Seed Parameters
3.3. Seed Separation
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Arista, M.; Herrera, J.; Talavera, S. Abies pinsapo Boiss.: A protected species in a protected area. Bocconea 1997, 7, 427–436. [Google Scholar]
- Edwards, D.G.W. Abies P. Mill. In Woody Plant Seed Manual—Part II; Agriculture Handbook 727; United States Department of Agriculture, Forest Service: Washington, DC, USA, 2008; pp. 149–198. [Google Scholar]
- Linares, J.C. Biogeography and evolution of Abies (Pinaceae) in the Mediterranean Basin: The roles of long-term climatic change and glacial refugia. J. Biogeogr. 2011, 38, 619–630. [Google Scholar] [CrossRef]
- Aguirre-Planter, É.; Jaramillo-Correa, J.P.; Gómez-Acevedo, S.; Khasa, D.P.; Bousquet, J.; Eguiarte, L.E. Phylogeny, diversification rates and species boundaries of Mesoamerican firs (Abies, Pinaceae) in a genus-wide context. Mol. Phylogenet. Evol. 2012, 62, 263–274. [Google Scholar] [CrossRef]
- Ghimire, B.; Lee, C.; Yang, J.; Heo, K. Comparative leaf anatomy of some species of Abies and Picea (Pinaceae). Acta Bot. Brasilica 2015, 29, 346–353. [Google Scholar] [CrossRef]
- Caudullo, G.; Tinner, W. Abies—Circum-Mediterranean firs in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publication Office of the European Union: Luxembourg, Luksembourg, 2016; p. e015be7+. [Google Scholar]
- Agrawal, T. Abies: A threatened genus. Br. J. Res. 2017, 4, 26. [Google Scholar] [CrossRef]
- Seneta, W.; Dolatowski, J. Dendrologia (Dendrology); Wydawnictwo Naukowe PWN: Warszawa, Poland, 2012; pp. 31–38. ISBN 9788301153694. (In Polish) [Google Scholar]
- Huth, F.; Wehnert, A.; Tiebel, K.; Wagner, S. Direct seeding of silver fir (Abies alba Mill.) to convert Norway spruce (Picea Abies L.) forests in Europe: A review. Forest Ecol. Manag. 2017, 403, 61–78. [Google Scholar] [CrossRef]
- Gosling, P. Raising Trees and Shrubs from Seed (Practice Guide (FCPS)); Forestry Commission: Edinburgh, UK, 2007; pp. 1–34. ISBN 9780855387365.
- Varsamis, G.K.; Takos, I.A.; Merou, T.P.; Galatsidas, S.A.; Panayiotis, D.D. Germination characteristics of Abies × borisii-regis. Seed Technol. 2014, 36, 51–59. [Google Scholar]
- Bhat, H.A.; Mughal, A.H.; Din Dar, M.U.; Mugloo, J.A. Cone, seed and germination characteristics in silver fir (Abies pindrow Spach) along the altitudinal gradient in western Himalayas. Int. J. Chem. Stud. 2018, 6, 2052–2055. [Google Scholar]
- Skrzyszewska, K.; Chłanda, J. A study on the variation of morphological characteristics of silver fir (Abies alba Mill.) seeds and their internal structure determined by X-ray radiography in the Beskid Sądecki and Beskid Niski mountain ranges of the Carpathians (Southern Poland). J. For. Sci. 2009, 55, 403–414. [Google Scholar] [CrossRef]
- Sevik, H.; Yahyaoglu, Z.; Turna, I. Determination of genetic variation between populations of Abies nordmanniana subsp. bornmulleriana Mattf according to some seed characteristics. In Genetic Diversity in Plants; Caliskan, M., Ed.; InTech: Rijeka, Croatia, 2012; pp. 231–248. ISBN 9789535101857. [Google Scholar]
- Sabor, J. Relation between the weight and the germination capacity of seed of silver fir. Sylwan 1984, 4, 59–69. (In Polish) [Google Scholar]
- Załęski, A. Nasiennictwo Leśnych Drzew i Krzewów Iglastych (Management of Coniferous Forest Trees and Shrubs for Seed Production); Oficyna Edytorska “Wydawnictwo Świat”: Warszawa, Poland, 1995; pp. 1–178. ISBN 8385597271. (In Polish) [Google Scholar]
- Khan, M.L. Effects of seed mass on seedling success in Artocarpus heterophyllus L., a tropical tree species of north-east India. Acta Oecol. 2004, 25, 103–110. [Google Scholar] [CrossRef]
- Upadhaya, K.; Pandey, H.N.; Law, P.S. The effect of seed mass on germination, seedling survival and growth in Prunus jenkinsii Hook.f. & Thoms. Turk. J. Bot. 2007, 31, 31–36. [Google Scholar] [CrossRef]
- Norden, N.; Daws, M.I.; Antoine, C.; Gonzalez, M.A.; Garwood, N.C.; Chave, J. The relationship between seed mass and mean time to germination for 1037 tree species across five tropical forests. Funct. Ecol. 2009, 23, 203–210. [Google Scholar] [CrossRef]
- Ballian, D. Genetic overload of silver fir (Abies alba Mill.) from five populations from central Bosnia and Herzegovina. Folia For. Pol. Ser. A For. 2013, 55, 49–57. [Google Scholar] [CrossRef]
- Chaisurisri, K.; Edwards, D.G.W.; El-Kassaby, Y.A. Effects of seed size on seedling attributes in Sitka spruce. New For. 1994, 8, 81–87. [Google Scholar]
- Grochowicz, J. Maszyny do Czyszczenia i Sortowania Nasion (Seed Cleaning and Sorting Machines); Wydawnictwo Akademii Rolniczej: Lublin, Poland, 1994; pp. 25–33. ISBN 839016129X. (In Polish) [Google Scholar]
- Kaliniewicz, Z.; Poznański, A. Variability and correlation of selected physical attributes of small-leaved lime (Tilia cordata Mill.) seeds. Sylwan 2013, 157, 39–46. (In Polish) [Google Scholar]
- Kaliniewicz, Z.; Markowski, P.; Anders, A.; Jadwisieńczak, B.; Rawa, T.; Szczechowicz, D. Basic physical properties of Norway spruce (Picea abies (L.) Karst.) seed. Tech. Sci. 2016, 19, 103–115. [Google Scholar]
- Mohsenin, N.N. Physical Properties of Plant and Animal Materials; Gordon and Breach Science Public: New York, NY, USA, 1986; pp. 1–891. ISBN 9780677213705. [Google Scholar]
- Kaliniewicz, Z. Analysis of frictional properties of cereal seed. Afr. J. Agricult. Res. 2013, 8, 5611–5621. [Google Scholar] [CrossRef]
- Rabiej, M. Statystyka z Programem Statistica (Statisctics in Statistica Software); Helion: Gliwice, Poland, 2012; pp. 1–344. ISBN 9788324641109. (In Polish) [Google Scholar]
- Kaliniewicz, Z.; Mańkowski, S.; Tylek, P.; Krzysiak, Z.; Peda, W. Correlations between the physical properties of silver fir seeds. Acta Agroph. 2018, 25, 197–212. (In Polish) [Google Scholar] [CrossRef]
- Tylek, P. Problems of pneumatic selection of forest tree seeds. Sylwan 1999, 12, 65–72. (In Polish) [Google Scholar]
- Tylek, P. Efficiency of some basic criteria of silver fir seeds separation. Acta Agroph. 2003, 2, 857–866. (In Polish) [Google Scholar]
- Tracz, M.; Barzdajn, W. The morphological traits of cones and seeds of Abies alba in the Middle Sudeten. Dendrobiology 2007, 58, 59–65. [Google Scholar]
- Velioğlu, E.; Tayanç, Y.; Çengel, B.; Kandemir, G. Genetic variability of seed characteristics of Abies populations from Turkey. J. Kast. Forf. 2012, 3, 27–35. [Google Scholar]
- Masoodi, H.U.R.; Thapliyal, M.; Singh, V.R.R. Studies on the variation in germination and seedling growth of Abies pindrow Spach. (Royle) in Garhwal region of Uttarakhand, India. J. Appl. Nat. Sci. 2014, 6, 711–715. [Google Scholar] [CrossRef]
- Song, J.-H.; Jang, K.-H.; Hur, S.-D. Variation of seed and germination characteristics of natural populations of Abies koreana Wilson, a Korean endemic species. J. Korean For. Soc. 2010, 99, 849–854. [Google Scholar]
- Oleksyn, J.; Reich, P.B.; Tjoelker, M.G.; Chalupka, W. Biogeographic differences in shoot elongation pattern among European Scots pine populations. For. Ecol. Manag. 2001, 148, 207–220. [Google Scholar] [CrossRef]
- Moles, A.T.; Westoby, M. Latitude, seed predation and seed mass. J. Biogeogr. 2003, 30, 105–128. [Google Scholar] [CrossRef]
- Gradečki-Poštenjak, M.; Ćelepirović, N. The influence of crown defoliation on the variability of some physiological and morphological properties of silver fir (Abies alba) seeds in the seed zone of Dinaric beech-fir forests in Croatia. Period. Biol. 2015, 117, 479–492. [Google Scholar] [CrossRef]
- Tylek, P. Agrophysical characteristics of silver fir seeds after pneumatic separation. Acta Sci. Pol. Silv. Colendar. Rat. Ind. Lignar. 2005, 4, 97–105. (In Polish) [Google Scholar]
- Veech, J.A.; Charlet, D.A.; Jenkins, S.H. Interspecific variation in seed mass and the co-existence of conifer species: A null model test. Evol. Ecol. Res. 2000, 2, 353–363. [Google Scholar]
- Arista, M.; Talavera, S. Density effect on the fruit-set, seed crop viability and seedling vigour of Abies pinsapo. Ann. Bot. 1996, 77, 187–192. [Google Scholar] [CrossRef]
- Andersen, U.S.; Cordova, J.P.; Nielsen, U.B.; Kollmann, J. Provenance variation in germination and seedling growth of Abies guatemalensis Rehder. Forest Ecol. Manag. 2008, 255, 1831–1840. [Google Scholar] [CrossRef]
- Kurt, Y.; Frampton, J.; Isik, F.; Landgren, C.; Chastagner, G. Variation in needle and cone characteristics and seed germination ability of Abies bornmuelleriana and Abies equi-trojani populations from Turkey. Turk. J. Agric. For. 2016, 40, 169–176. [Google Scholar] [CrossRef]
- Nielsen, U.B.; Hansen, O.K. Response to selfing in seed set, seedling establishment and nursery growth based on controlled crosses of Abies nordmanniana clones. Silvae Genet. 2009, 59, 90–98. [Google Scholar] [CrossRef]
- McCartan, S.A.; Jinks, R.L. Upgrading seed lots of European silver fir (Abies alba Mill.) using imbibition-drying-separation. Tree Planters’ Notes 2015, 58, 21–27. [Google Scholar]
- Tylek, P. Analysis of aerodynamic properties of common fir and common beech. Inżynieria Rolnicza 2011, 6, 247–253. (In Polish) [Google Scholar]
- Aniszewska, M.; Gendek, A.; Śliwińka, J. Variability of silver fir (Abies alba Mill.) cones—Variability structure of scale surface area. For. Res. Pap. 2017, 78, 5–13. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Fir Species | Property | |||||
|---|---|---|---|---|---|---|
| Terminal Velocity v | Thickness T | Width W | Length L | Angle of External Friction γ | Mass m | |
| (m s−1) | (mm) | (mm) | (mm) | (°) | (mg) | |
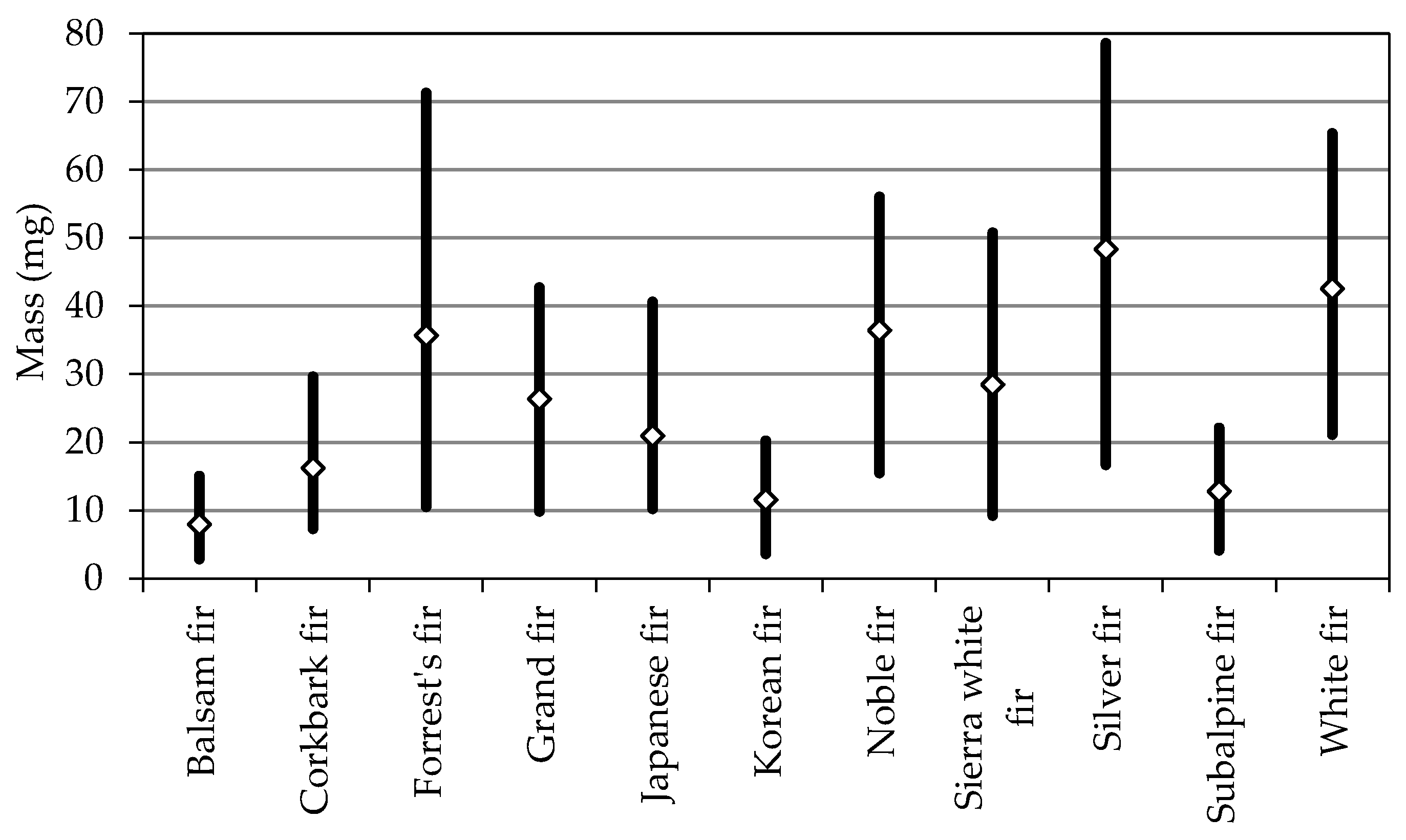
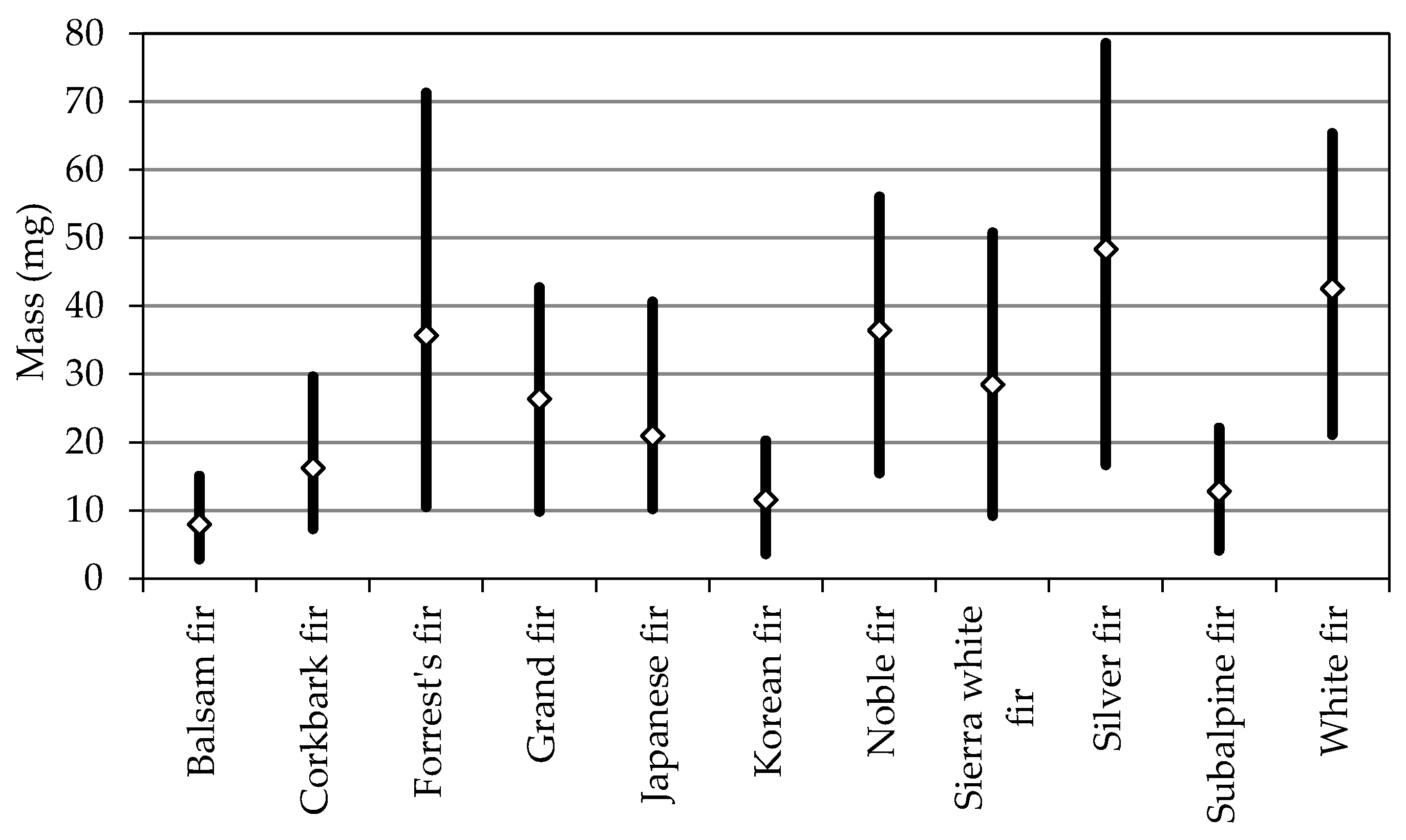
| Balsam fir | 4.8 ± 0.7 a | 1.78 ± 0.19 a | 3.29 ± 0.46 a | 5.44 ± 0.46 a | 33 ± 5 e | 7.9 ± 2.4 a |
| Corkbark fir | 5.7 ± 0.6 cd | 1.99 ± 0.27 c | 3.90 ± 0.52 b | 6.90 ± 0.72 c | 26 ± 3 a | 16.2 ± 4.8 c |
| Forrest’s fir | 5.7 ± 0.9 d | 2.32 ± 0.33 d | 5.39 ± 0.84 e | 10.95 ± 1.54 f | 27 ± 3 ab | 35.7 ± 13.2 f |
| Grand fir | 5.4 ± 0.8 b | 2.31 ± 0.24 d | 4.81 ± 0.64 d | 9.41 ± 0.85 d | 31 ± 4 d | 26.4 ± 6.5 e |
| Japanese fir | 5.4 ± 0.8 b | 1.99 ± 0.29 bc | 3.94 ± 0.68 b | 9.63 ± 1.06 d | 30 ± 5 d | 20.9 ± 6.1 d |
| Korean fir | 5.8 ± 0.9 d | 1.76 ± 0.22 a | 3.46 ± 0.57 a | 5.79 ± 0.68 b | 28 ± 3 bc | 11.6 ± 3.4 b |
| Noble fir | 6.1 ± 0.6 e | 2.59 ± 0.28 e | 4.43 ± 0.69 c | 11.06 ± 1.07 f | 27 ± 3 ab | 36.5 ± 9.6 f |
| Sierra white fir | 5.5 ± 0.7 bc | 2.29 ± 0.24 d | 4.91 ± 0.78 d | 9.48 ± 1.03 d | 27 ± 4 ab | 28.5 ± 7.4 e |
| Silver fir | 7. 2 ± 0.9 g | 3.22 ± 0.39 f | 5.57 ± 0.83 e | 10.83 ± 1.02 f | 28 ± 5 bc | 48.3 ± 12.9 h |
| Subalpine fir | 4.9 ± 0.7 a | 1.91 ± 0.27 b | 3.95 ± 0.62 b | 7.04 ± 0.82 c | 29 ± 4 c | 12.9 ± 3.7 b |
| White fir | 6.4 ± 0.7 f | 2.58 ± 0.30 e | 5.40 ± 0.85 e | 10.41 ± 1.12 e | 27 ± 3 ab | 42.6 ± 9.4 g |
| Fir Species | Indicator | |||||
|---|---|---|---|---|---|---|
| Geometric Mean Diameter D | Aspect | Aspect | Aspect | Sphericity Index Φ | Specific Mass mD | |
| (mm) | Ratio T/W (%) | Ratio T/L (%) | Ratio W/L (%) | (%) | (g m−1) | |
| Balsam fir | 3.16 ± 0.27 a | 54.97 ± 7.91 d | 32.80 ± 3.17 g | 60.58 ± 8.29 d | 58.20 ± 3.73 h | 2.48 ± 0.62 a |
| Corkbark fir | 3.76 ± 0.37 b | 51.77 ± 8.45 c | 28.98 ± 3.55 e | 56.79 ± 7.42 c | 54.62 ± 3.51 f | 4.26 ± 0.94 c |
| Forrest’s fir | 5.13 ± 0.57 f | 43.76 ± 8.04 a | 21.41 ± 3.39 a | 49.75 ± 7.66 b | 47.18 ± 3.95 c | 6.82 ± 2.01 g |
| Grand fir | 4.69 ± 0.36 d | 48.78 ± 7.69 b | 24.71 ± 3.14 c | 51.29 ± 3.64 b | 50.07 ± 3.34 d | 5.60 ± 1.20 e |
| Japanese fir | 4.20 ± 0.41 c | 51.78 ± 11.29 c | 20.78 ± 3.26 a | 41.35 ± 7.98 a | 43.86 ± 4.12 a | 4.92 ± 1.12 d |
| Korean fir | 3.26 ± 0.36 a | 51.80 ± 8.41 c | 30.61 ± 4.11 f | 59.85 ± 8.08 d | 56.60 ± 4.12 g | 3.49 ± 0.80 b |
| Noble fir | 5.01 ± 0.45 e | 59.83 ± 10.64 e | 23.58 ± 2.88 b | 40.11 ± 5.59 a | 45.40 ± 3.03 b | 7.22 ± 1.54 h |
| Sierra white fir | 4.72 ± 0.43 d | 47.66 ± 8.45 b | 24.31 ± 3.00 bc | 52.01 ± 7.84 b | 50.00 ± 3.64 d | 5.98 ± 1.30 f |
| Silver fir | 5.77 ± 0.51 g | 58.94 ± 10.26 e | 29.83 ± 3.33 ef | 51.61 ± 7.54 b | 53.40 ± 3.45 e | 8.29 ± 1.76 i |
| Subalpine fir | 3.74 ± 0.36 b | 49.62 ± 10.39 bc | 27.37 ± 4.07 d | 56.51 ± 9.27 c | 53.41 ± 4.32 e | 3.39 ± 0.77 b |
| White fir | 5.24 ± 0.50 f | 48.77 ± 8.15 b | 25.03 ± 3.40 c | 52.02 ± 7.13 b | 50.50 ± 3.61 d | 8.08 ± 1.36 i |
| Property | v | T | W | L | γ | m | D | T/W | T/L | W/L | Φ |
|---|---|---|---|---|---|---|---|---|---|---|---|
| T | 0.64 * | 1 | |||||||||
| W | 0.33 * | 0.60 * | 1 | ||||||||
| L | 0.39 * | 0.69 * | 0.69 * | 1 | |||||||
| γ | −0.28 * | −0.25 * | −0.26 * | −0.23 * | 1 | ||||||
| m | 0.70 * | 0.84 * | 0.75 * | 0.82 * | −0.28 * | 1 | |||||
| D | 0.51 * | 0.86 * | 0.87 * | 0.91 * | −0.28 * | 0.92 * | 1 | ||||
| T/W | 0.31 * | 0.38 * | −0.48 * | −0.03 | 0.02 | 0.05 | −0.05 | 1 | |||
| T/L | 0.16 * | 0.14 * | −0.30 * | −0.60 * | 0.04 | −0.22 * | −0.30 * | 0.47 * | 1 | ||
| W/L | −0.16 * | −0.24 * | 0.19 * | −0.55 * | 0.02 | −0.26 * | −0.24 * | −0.52 * | 0.49 * | 1 | |
| Φ | 0.01 | −0.06 | −0.05 | −0.67 * | 0.03 | −0.27 * | −0.31 * | −0.03 | 0.86 * | 0.86 * | 1 |
| mD | 0.75 * | 0.78 * | 0.67 * | 0.79 * | −0.30 * | 0.99 * | 0.85 * | 0.08 * | −0.24 * | −0.31 * | −0.31 * |
| Fir Species | Coefficient of Correlation between Seed Mass m and: | ||||
|---|---|---|---|---|---|
| Terminal Velocity v | Thickness T | Width W | Length L | Angle of External Friction γ | |
| Balsam fir | 0.797 * | 0.669 * | 0.416 * | 0.650 * | −0.499 * |
| Corkbark fir | 0.626 * | 0.675 * | 0.579 * | 0.698 * | −0.149 |
| Forrest’s fir | 0.786 * | 0.617 * | 0.494 * | 0.691 * | −0.119 |
| Grand fir | 0.677 * | 0.486 * | 0.308 * | 0.321 * | −0.218 * |
| Japanese fir | 0.659 * | 0.659 * | 0.398 * | 0.503 * | −0.556 * |
| Korean fir | 0.587 * | 0.705 * | 0.541 * | 0.647 * | −0.232 * |
| Noble fir | 0.718 * | 0.645 * | 0.517 * | 0.474 * | −0.079 |
| Sierra white fir | 0.582 * | 0.580 * | 0.496 * | 0.432 * | −0.258 * |
| Silver fir | 0.578 * | 0.625 * | 0.487 * | 0.596 * | −0.042 |
| Subalpine fir | 0.657 * | 0.552 * | 0.430 * | 0.624 * | −0.282 * |
| White fir | 0.453 * | 0.472 * | 0.540 * | 0.649 * | −0.196 * |
| Fir Species | Seed Fraction | Percentage (%) | Mass (mg) | Coefficient of Variation (%) of Seed Mass | |
|---|---|---|---|---|---|
| Fraction | Total | ||||
| Balsam fir | I (v < 4.5 m s−1) | 30.7 | 5.46 | 25.91 | 30.10 |
| II (v = 4.5–5.5 m s−1) | 56.4 | 8.63 | 18.10 | ||
| III (v > 5.5 m s−1) | 12.9 | 10.77 | 19.31 | ||
| Corkbark fir | I (v < 5.5 m s−1) | 28.8 | 12.54 | 31.13 | 29.49 |
| II (v = 5.5–6.0 m s−1) | 44.2 | 15.24 | 20.09 | ||
| III (v > 6.0 m s−1) | 27.0 | 21.01 | 18.59 | ||
| Forrest’s fir | I (v < 5.0 m s−1) | 16.8 | 20.67 | 32.37 | 36.90 |
| II (v = 5.0–6.0 m s−1) | 42.1 | 31.51 | 29.62 | ||
| III (v > 6.0 m s−1) | 41.1 | 46.17 | 21.23 | ||
| Grand fir | I (v < 5.0 m s−1) | 19.4 | 19.10 | 27.29 | 24.45 |
| II (v = 5.0–5.5 m s−1) | 35.2 | 25.72 | 16.44 | ||
| III (v > 5.5 m s−1) | 45.4 | 30.06 | 18.34 | ||
| Japanese fir | I (v < 5.0 m s−1) | 24.5 | 15.79 | 24.23 | 29.13 |
| II (v = 5.0–5.5 m s−1) | 30.4 | 18.77 | 22.87 | ||
| III (v > 5.5 m s−1) | 45.1 | 25.21 | 20.55 | ||
| Korean fir | I (v < 5.0 m s−1) | 17.0 | 7.62 | 29.59 | 29.54 |
| II (v = 5.0–6.0 m s−1) | 36.8 | 11.19 | 27.38 | ||
| III (v > 6.0 m s−1) | 46.2 | 13.27 | 20.34 | ||
| Noble fir | I (v < 5.5 m s−1) | 18.3 | 23.75 | 21.80 | 26.35 |
| II (v = 5.5–6.5 m s−1) | 69.2 | 37.88 | 19.53 | ||
| III (v > 6.5 m s−1) | 12.5 | 47.36 | 12.03 | ||
| Sierra white fir | I (v < 5.0 m s−1) | 15.7 | 20.56 | 35.59 | 26.13 |
| II (v = 5.0–5.5 m s−1) | 39.2 | 27.08 | 19.47 | ||
| III (v > 5.5 m s−1) | 45.1 | 32.41 | 20.10 | ||
| Silver fir | I (v < 6.5 m s−1) | 26.3 | 37.49 | 32.85 | 26.61 |
| II (v = 6.5–7.5 m s−1) | 51.7 | 49.65 | 21.21 | ||
| III (v > 7.5 m s−1) | 22.0 | 58.21 | 14.85 | ||
| Subalpine fir | I (v < 4.5 m s−1) | 24.5 | 9.67 | 32.08 | 28.70 |
| II (v = 4.5–5.0 m s−1) | 33.0 | 11.93 | 22.76 | ||
| III (v > 5.0 m s−1) | 42.5 | 15.40 | 18.35 | ||
| White fir | I (v < 6.0 m s−1) | 26.9 | 36.37 | 22.01 | 22.16 |
| II (v = 6.0–6.5 m s−1) | 37.5 | 41.88 | 21.46 | ||
| III (v > 6.5 m s−1) | 35.6 | 48.04 | 16.18 | ||
| Fir Species | Seed Fraction | Percentage (%) | Mass (mg) | Coefficient of Variation (%) of Seed Mass | |
|---|---|---|---|---|---|
| Fraction | Total | ||||
| Balsam fir | I (T ≤ 1.60 mm) | 18.8 | 5.99 | 21.30 | 30.10 |
| II (T = 1.61–1.80 mm) | 40.6 | 7.09 | 27.06 | ||
| III (T > 1.80 mm) | 40.6 | 9.68 | 21.15 | ||
| Corkbark fir | I (T ≤ 1.80 mm) | 23.1 | 11.75 | 28.20 | 29.49 |
| II (T = 1.81–2.10 mm) | 43.3 | 15.85 | 23.62 | ||
| III (T > 2.10 mm) | 33.6 | 19.81 | 20.38 | ||
| Forrest’s fir | I (T ≤ 2.10 mm) | 26.2 | 24.36 | 35.05 | 36.90 |
| II (T = 2.11–2.50 mm) | 41.1 | 36.04 | 28.01 | ||
| III (T > 2.50 mm) | 32.7 | 44.40 | 29.43 | ||
| Grand fir | I (T ≤ 2.10 mm) | 22.2 | 22.00 | 30.51 | 24.45 |
| II (T = 2.11–2.40 mm) | 42.6 | 26.36 | 19.08 | ||
| III (T > 2.40 mm) | 35.2 | 29.23 | 21.88 | ||
| Japanese fir | I (T ≤ 1.80 mm) | 27.5 | 15.22 | 18.78 | 29.13 |
| II (T = 1.81–2.10 mm) | 32.4 | 21.28 | 25.12 | ||
| III (T > 2.10 mm) | 40.1 | 24.58 | 21.92 | ||
| Korean fir | I (T ≤ 1.60 mm) | 19.8 | 8.00 | 31.17 | 29.54 |
| II (T = 1.61–1.80 mm) | 45.3 | 11.38 | 24.40 | ||
| III (T > 1.80 mm) | 34.9 | 13.77 | 20.44 | ||
| Noble fir | I (T ≤ 2.40 mm) | 28.8 | 28.72 | 26.12 | 26.35 |
| II (T = 2.41–2.70 mm) | 37.5 | 35.85 | 22.27 | ||
| III (T > 2.70 mm) | 33.7 | 43.83 | 16.36 | ||
| Sierra white fir | I (T ≤ 2.10 mm) | 25.5 | 24.54 | 25.33 | 26.13 |
| II (T = 2.11–2.40 mm) | 45.1 | 27.41 | 23.28 | ||
| III (T > 2.40 mm) | 29.4 | 33.47 | 22.11 | ||
| Silver fir | I (T ≤ 3.00 mm) | 26.3 | 38.09 | 31.36 | 26.61 |
| II (T = 3.01–3.40 mm) | 44.9 | 47.32 | 20.12 | ||
| III (T > 3.40 mm) | 28.8 | 59.28 | 16.03 | ||
| Subalpine fir | I (T ≤ 1.80 mm) | 34.9 | 10.25 | 27.33 | 28.70 |
| II (T = 1.81–2.10 mm) | 36.8 | 13.40 | 25.43 | ||
| III (T > 2.10 mm) | 28.3 | 15.34 | 19.38 | ||
| White fir | I (T ≤ 2.40 mm) | 30.8 | 37.69 | 18.58 | 22.16 |
| II (T = 2.41–2.70 mm) | 32.7 | 42.69 | 19.92 | ||
| III (T > 2.70 mm) | 36.5 | 46.61 | 21.98 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaliniewicz, Z.; Markowski, P.; Anders, A.; Jadwisieńczak, K.; Żuk, Z.; Krzysiak, Z. Physical Properties of Seeds of Eleven Fir Species. Forests 2019, 10, 142. https://doi.org/10.3390/f10020142
Kaliniewicz Z, Markowski P, Anders A, Jadwisieńczak K, Żuk Z, Krzysiak Z. Physical Properties of Seeds of Eleven Fir Species. Forests. 2019; 10(2):142. https://doi.org/10.3390/f10020142
Chicago/Turabian StyleKaliniewicz, Zdzisław, Piotr Markowski, Andrzej Anders, Krzysztof Jadwisieńczak, Zbigniew Żuk, and Zbigniew Krzysiak. 2019. "Physical Properties of Seeds of Eleven Fir Species" Forests 10, no. 2: 142. https://doi.org/10.3390/f10020142
APA StyleKaliniewicz, Z., Markowski, P., Anders, A., Jadwisieńczak, K., Żuk, Z., & Krzysiak, Z. (2019). Physical Properties of Seeds of Eleven Fir Species. Forests, 10(2), 142. https://doi.org/10.3390/f10020142