Genome-Wide Analysis and Expression Profiling of the Heat Shock Factor Gene Family in Phyllostachys edulis during Development and in Response to Abiotic Stresses


Abstract
1. Introduction
2. Materials and Methods
2.1. Database Searches for Hsf Genes in Moso Bamboo and Analyses of Physicochemical Characteristics
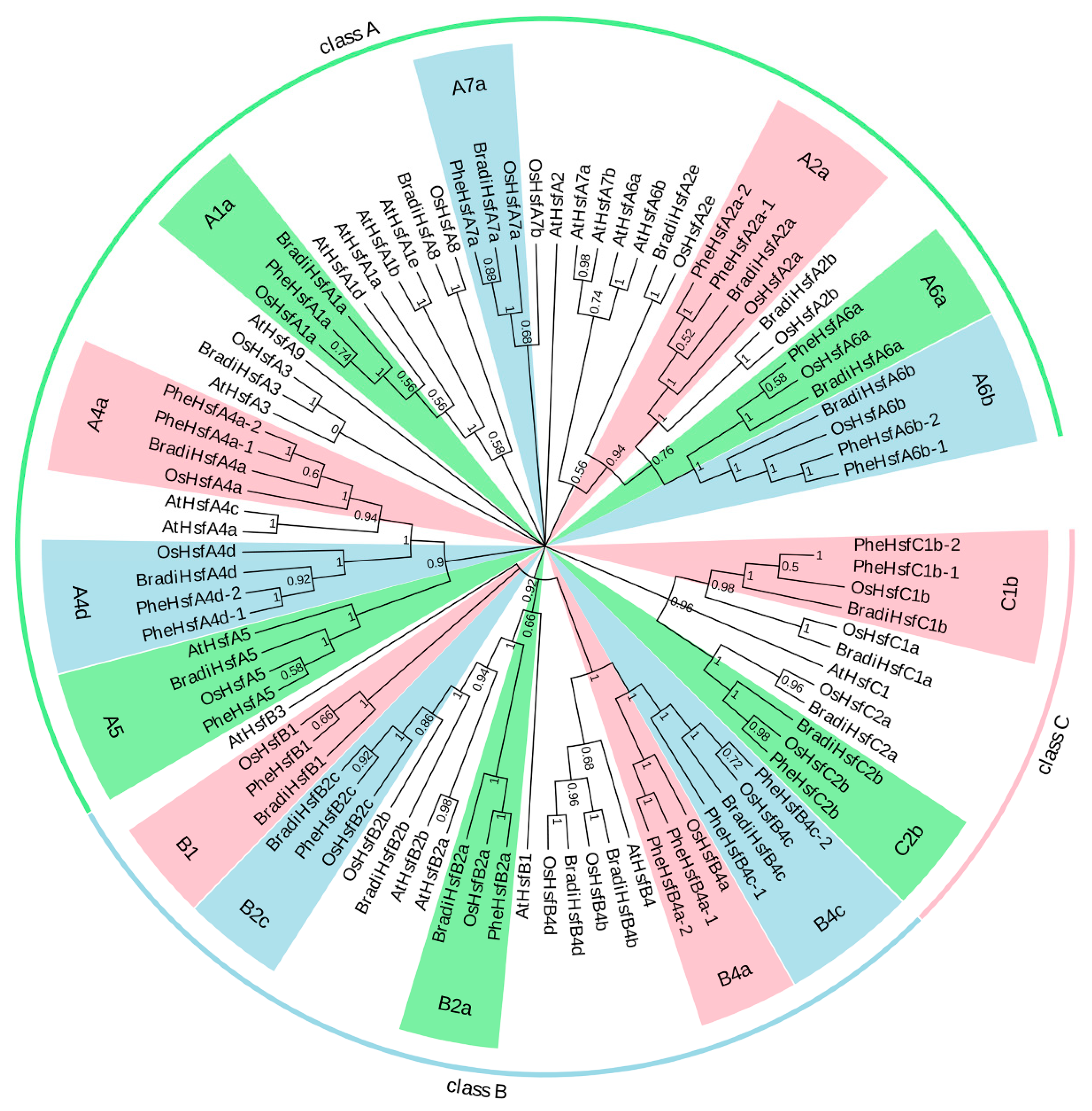
2.2. Phylogenetic Analysis
2.3. Structural and Motif Analyses of PheHsf Genes
2.4. Cis-Regulatory Element Analysis of PheHsf Genes
2.5. Plant Material
2.6. RNA Isolation and Relative Expression Level Analysis of PheHsfs
2.7. Co-Expression Network and Protein Interactions of PheHsfs
3. Results
3.1. The Hsf Family Genes in Moso Bamboo
3.2. Phylogenetic Relationships and Multiple Sequence Alignment of PheHsf Genes
3.3. Structure and Motif Analyses of PheHsf Genes
3.4. Cis-Regulatory Element Analysis in Promoters of PheHsfs
3.5. Expression Pattern of the PheHsf Genes in Shoot and Flower Development
3.6. PheHsfAs Expression in Moso Bamboo in Response to Various Stresses
3.7. Expression Correlation and Interaction Networks
4. Discussion
4.1. Characterization of the Moso Bamboo Hsf Genes Family
4.2. Cis-Regulatory Element Analysis in the Promoters of PheHsfs
4.3. PheHsfAs Involvement in Development Processes
4.4. PheHsfAs are Involved in Stress Responses
4.5. Expression Correlation and Interaction Networks
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Li, L.; Cheng, Z.; Ma, Y.; Bai, Q.; Li, X.; Cao, Z.; Wu, Z.; Gao, J. The association of hormone signaling genes, transcription, and changes in shoot anatomy during moso bamboo growth. Plant Biotechnol. J. 2017, 16, 72–85. [Google Scholar] [CrossRef] [PubMed]
- Lobovikov, M.; Paudel, S.; Piazza, M.; Ren, H.; Wu, J. World Bamboo Resources: A Thematic Study Prepared in the Framework of the Global Forest Resources Assessment 2005; Food and Agriculture Organization of the United Nations: Rome, Italy, 2007; pp. 1–73, 1020–3370. [Google Scholar]
- Song, X.; Zhou, G.; Jiang, H.; Yu, S.; Fu, J.; Li, W. Carbon sequestration by Chinese bamboo forests and their ecological benefits: Assessment of potential, problems, and future challenges. Environ. Rev. 2011, 19, 418–428. [Google Scholar] [CrossRef]
- Li, L.; Hu, T.; Li, X.; Mu, S.; Cheng, Z.; Ge, W.; Gao, J. Genome-wide analysis of shoot growth-associated alternative splicing in moso bamboo. Mol. Genet. Genom. 2016, 291, 1695–1714. [Google Scholar] [CrossRef]
- Zhou, G.M.; Wu, J.S.; Jiang, P.K. Effects of different management models on carbon storage in Phyllostachys pubescens forests. J. Beijing For. Univ. 2006, 28, 51–55. [Google Scholar] [CrossRef]
- Liu, L.; Dong, D.; Li, Y.; Li, X.; Bureau, A.M. Investigation of moso bamboo forest under high temperature and drought disaster. World Bamboo Rattan 2014, 12, 24–27. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, J.; Zhang, H. Measures of water management and increasing drought resistance of moso forests in Anji County, Zhejiang Province. World Bamboo Rattan 2008, 6, 23–24. [Google Scholar] [CrossRef]
- Rizhsky, L.; Liang, H.; Mittler, R. The combined effect of drought stress and heat shock on gene expression in tobacco. Plant Physiol. 2002, 130, 1143–1151. [Google Scholar] [CrossRef]
- Chauhan, H.; Khurana, N.; Agarwal, P.; Khurana, P. Heat shock factors in rice (Oryza sativa L.): Genome-wide expression analysis during reproductive development and abiotic stress. Mol. Genet. Genom. 2011, 286, 171–187. [Google Scholar] [CrossRef]
- Hartl, F.U.; Hayerhartl, M. Molecular chaperones in the cytosol: From nascent chain to folded protein. Science 2002, 295, 1852–1858. [Google Scholar] [CrossRef]
- Queitsch, C.; Hong, S.W.; Vierling, E.; Lindquist, S. Heat shock protein 101 plays a crucial role in thermotolerance in Arabidopsis. Plant Cell 2000, 12, 479–492. [Google Scholar] [CrossRef]
- Döring, P.; Treuter, E.; Kistner, C.; Lyck, R.; Chen, A.; Nover, L. The role of AHA motifs in the activator function of tomato heat stress transcription factors HsfA1 and HsfA2. Plant Cell 2000, 12, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Pelham, H.R.; Bienz, M. A synthetic heat-shock promoter element confers heat-inducibility on the herpes simplex virus thymidine kinase gene. EMBO J. 1982, 1, 1473–1477. [Google Scholar] [CrossRef] [PubMed]
- Von Koskull-Döring, P.; Scharf, K.; Nover, L. The diversity of plant heat stress transcription factors. Trends Plant Sci. 2007, 12, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Scharf, K.D.; Berberich, T.; Ebersberger, I.; Nover, L. The plant heat stress transcription factor (Hsf) family: Structure, function and evolution. Biochim. Biophys. Acta 2012, 1819, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Kotak, S.; Port, M.; Ganguli, A.; Bicker, F.; von Koskull-Döring, P. Characterization of C-terminal domains of Arabidopsis heat stress transcription factors (Hsfs) and identification of a new signature combination of plant class A Hsfs with AHA and NES motifs essential for activator function and intracellular localization. Plant J. 2004, 39, 98–112. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.K.; Tripp, J.; Winkelhaus, S.; Tschiersch, B.; Theres, K.; Nover, L.; Scharf, K.D. In the complex family of heat stress transcription factors, HsfA1 has a unique role as master regulator of thermotolerance in tomato. Genes Dev. 2002, 16, 1555–1567. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, D.; Yamaguchi, K.; Nishiuchi, T. High-level overexpression of the Arabidopsis HsfA2 gene confers not only increased themotolerance but also salt/osmotic stress tolerance and enhanced callus growth. J. Exp. Bot. 2007, 12, 3373–3383. [Google Scholar] [CrossRef]
- Wang, X.; Huang, W.; Liu, J.; Yang, Z.; Huang, B. Molecular regulation and physiological functions of a novel FaHsfA2c cloned from tall fescue conferring plant tolerance to heat stress. Plant Biotechnol. J. 2017, 15, 237–248. [Google Scholar] [CrossRef]
- Zhang, L.; Li, Y.; Xing, D.; Gao, C. Characterization of mitochondrial dynamics and subcellular localization of ROS reveal that HsfA2 alleviates oxidative damage caused by heat stress in Arabidopsis. J. Exp. Bot. 2009, 60, 2073–2091. [Google Scholar] [CrossRef]
- Sakuma, Y.; Maruyama, K.; Qin, F.; Osakabe, Y.; Shinozaki, K. Yamaguchi-Shino-zaki, K. Dual function of an Arabidopsis transcription factor DREB2A in water-stress-responsive and heat-stress-responsive gene expression. Proc. Natl. Acad. Sci. USA 2006, 103, 18822–18827. [Google Scholar] [CrossRef]
- Schramm, F.; Larkindale, J.; Kiehlmann, E.; Ganguli, A.; Englich, G.; Vierling, E.; von Koskull-Döring, P. A cascade of transcription factor DREB2A and heat stress transcription factor HsfA3 regulates the heat stress response of Arabidopsis. Plant J. 2008, 53, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Sakuma, Y.; Todaka, D.; Maruyama, K.; Qin, F.; Mizoi, J.; Kidokoro, S.; Fujita, Y.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of an Arabidopsis heat-shock transcription factor HsfA3 in the transcriptional cascade downstream of the DREB2A stress-regulatory system. Biochem. Biophys. Res. Commun. 2008, 368, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, L.; Wang, A.; Xu, X.; Li, J. Ectopic Overexpression of SlHsfA3, a Heat Stress Transcription Factor from Tomato, Confers Increased Thermotolerance and Salt Hypersensitivity in Germination in Transgenic Arabidopsis. PLoS ONE 2013, 8, e54880. [Google Scholar] [CrossRef] [PubMed]
- Lang, S.; Liu, X.; Xue, H.; Li, X.; Wang, X. Functional characterization of BnHSFA4a as a heat shock transcription factor in controlling the re-establishment of desiccation tolerance in seeds. J. Exp. Bot. 2017, 68, 2361–2375. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhang, H.; Zhao, H.; Gao, T.; Song, A.; Jiang, J.; Chen, F.; Chen, S. Chrysanthemum CmHSFA4 gene positively regulates salt stress tolerance in transgenic chrysanthemum. Plant Biotechnol. J. 2017, 16, 1311–1321. [Google Scholar] [CrossRef] [PubMed]
- Shim, D.; Hwang, J.U.; Lee, J.; Lee, S.; Choi, Y.; An, G.; Martinoia, E.; Lee, Y. Orthologs of the class A4 heat shock transcription factor HsfA4a confer cadmium tolerance in wheat and rice. Plant Cell 2009, 21, 4031–4043. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.S.; Jiang, J.; Han, X.J.; Zhang, Y.X.; Zhuo, R.Y. Identification, expression analysis of the Hsf family, and characterization of class A4 in Sedum Alfredii hance under cadmium stress. Int. J. Mol. Sci. 2018, 19, 1216. [Google Scholar] [CrossRef] [PubMed]
- Kotak, S.; Vierling, E.; Bäumlein, H.; von Koskull-Döring, P. A novel transcriptional cascade regulating expression of heat stress proteins during seed development of Arabidopsis. Plant Cell 2007, 19, 182–195. [Google Scholar] [CrossRef]
- Ikeda, M.; Ohme-Takagi, M. Arabidopsis HsfB1 and HsfB2b act as repressors of the expression of heat-inducible Hsfs but positively regulate the acquired thermotolerance. Plant Physiol. 2011, 157, 1243–1254. [Google Scholar] [CrossRef]
- Zhuang, L.L.; Cao, W.; Wang, J.; Yu, J.J.; Yang, Z.M.; Huang, B.G. Characterization and functional analysis of FaHsfC1b from Festuca arundinacea conferring heat tolerance in Arabidopsis. Int. J. Mol. Sci. 2018, 19, 2702. [Google Scholar] [CrossRef]
- Peng, Z.; Lu, Y.; Li, L.; Zhao, Q.; Feng, Q.; Gao, Z.; Lu, H.; Hu, T.; Yao, N.; Liu, K.; et al. The draft genome of the fast-growing non-timber forest species moso bamboo (Phyllostachys heterocycla). Nat. Genet. 2013, 45, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Hou, D.; Cheng, Z.; Xie, L.; Li, X.; Li, J.; Mu, S.; Gao, J. The R2R3MYB Gene Family in Phyllostachys edulis: Genome-Wide Analysis and Identification of Stress or Development-Related R2R3MYBs. Front. Plant Sci. 2018, 9, 738. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Wang, H.; Cai, D.; Gao, Y.; Zhang, H.; Wang, Y.; Lin, C.; Ma, L.; Gu, L. Comprehensive profiling of rhizome-associated alternative splicing and alternative polyadenylation in moso bamboo (Phyllostachys edulis). Plant J. 2017, 91, 684–699. [Google Scholar] [CrossRef]
- Jin, J.; He, K.; Tang, X.; Li, Z.; Lv, L.; Zhao, Y.; Luo, J.; Gao, G. An Arabidopsis transcriptional regulatory map reveals distinct functional and evolutionary features of novel transcription factors. Mol. Biol. Evol. 2015, 32, 1767–1773. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Tian, F.; Yang, D.C.; Meng, Y.Q.; Kong, L.; Luo, J.; Gao, G. PlantTFDB 4.0: Toward a central hub for transcription factors and regulatory interactions in plants. Nucleic Acids Res. 2017, 45, 1040–1045. [Google Scholar] [CrossRef]
- Yu, C.S.; Lin, C.J.; Hwang, J.K. Predicting subcellular localization of proteins for Gram-negative bacteria by support vector machines based on n-peptide compositions. Protein Sci. 2004, 13, 1402–1406. [Google Scholar] [CrossRef]
- Guo, J.; Wu, J.; Ji, Q.; Wang, C.; Luo, L.; Yuan, Y.; Wang, Y.; Wang, J. Genome-wide analysis of heat shock transcription factor families in rice and Arabidopsis. J. Genet. Genom. 2008, 35, 105–118. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Sievers, F.; Higgins, D.G. Clustal Omega, accurate alignment of very large numbers of sequences. Methods Mol. Biol. 2014, 1079, 105–116. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.Y.; Li, W.W.; Noble, W.S. MEME Suite: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Cheng, Z.C.; Hou, D.; Liu, J.; Li, X.Y.; Xie, L.H.; Ma, Y.J.; Gao, J. Characterization of moso bamboo (Phyllostachys edulis) Dof factors in floral development and abiotic stress responses. Genome 2018, 61, 151–156. [Google Scholar] [CrossRef]
- Liu, A.L.; Zou, J.; Zhang, X.W.; Zhou, X.Y.; Wang, W.F.; Xiong, X.Y.; Chen, L.Y.; Chen, X.B. Expression profiles of class A rice heat shock transcription factor genes under abiotic stresses. J. Plant Biol. 2010, 53, 142–149. [Google Scholar] [CrossRef]
- Gao, J.; Zhang, Y.; Zhang, C.; Qi, F.; Li, X.; Mu, S.; Peng, Z. Characterization of the floral transcriptome of moso bamboo (Phyllostachys edulis) at different flowering developmental stages by transcriptome sequencing and RNA-Seq analysis. PLoS ONE 2014, 9, e98910. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Fan, C.; Ma, J.; Guo, Q.; Li, X.; Wang, H.; Lu, M. Selection of reference genes for quantitative real-time PCR in bamboo (Phyllostachys edulis). PLoS ONE 2013, 8, e56573. [Google Scholar] [CrossRef]
- Livak, K.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Smita, S.; Katiyar, A.; Chinnusamy, V.; Pandey, D.M.; Bansal, K.C. Transcriptional regulatory network analysis of MYB transcription factor family genes in rice. Front. Plant Sci. 2015, 6, 1157. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Sultan, S.; Ali, M.; Nawaz, S.; Ali, M.A.; Shahzad, A. Genome wide analysis of heat shock factors (HSF) gene family of Arabidopsis Thaliana. J. Biol. Agric. Healthc. 2016, 6, 69–77. [Google Scholar]
- Li, P.S.; Zheng, W.J.; Zhou, Y.B.; Chen, M.; Chai, S.C.; Ma, L.J.; Xu, Z.S. Genome-wide identification, classification and high temperature responses of Hsf family in Brachypodium distachyon. J. China Agric. Univ. 2015, 20, 8–18. [Google Scholar] [CrossRef]
- Nover, L.; Bharti, K.; Döring, P.; Mishra, S.K.; Ganguli, A.; Scharf, K. Arabidopsis and the heat stress transcription factor world: How many heat stress transcription factors do we need? Cell Stress Chaperones 2001, 6, 177–189. [Google Scholar] [CrossRef]
- Guo, M.; Liu, J.H.; Ma, X.; Luo, D.X.; Gong, Z.H.; Lu, M.H. The plant heat stress transcription factors (HSFs): Structure, regulation, and function in response to abiotic stresses. Front. Plant Sci. 2016, 7, 114. [Google Scholar] [CrossRef]
- Xue, G.; Sadat, S.; Drenth, J.; McIntyre, C.L. The heat shock factor family from Triticum aestivum in response to heat and other major abiotic stresses and their role in regulation of heat shock protein genes. J. Exp. Bot. 2014, 65, 539–557. [Google Scholar] [CrossRef]
- Zhao, H.; Peng, Z.; Fei, B.; Li, L.; Hu, T.; Gao, Z.; Jiang, Z. BambooGDB: A bamboo genome database with functional annotation and an analysis platform. Database 2014, 2014, bau006. [Google Scholar] [CrossRef]
- Giorno, F.; Guerriero, G.; Baric, S.; Mariani, C. Heat shock transcriptional factors in Malus domestica: Identification, classification and expression analysis. BMC Genom. 2012, 13, 639. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Busch, W.; Wunderlich, M.; Schöffl, F. Identification of novel heat shock factor-dependent genes and biochemical pathways in Arabidopsis thaliana. Plant J. 2010, 41, 1–14. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Y.; Jia, H.X.; Li, J.B.; Huang, J.; Lu, M.Z.; Hu, J.J. The heat shock factor gene family in Salix suchowensis: A genome-wide survey and expression profiling during development and abiotic stresses. Front. Plant Sci. 2015, 6, 748. [Google Scholar] [CrossRef]
- Mittal, D.; Chakrabarti, S.; Sarkar, A.; Singh, A.; Grover, A. Heat shock factor gene family in rice: Genomic organization and transcript expression profiling in response to high temperature, low temperature and oxidative stresses. Plant Physiol. Biochem. 2009, 47, 785–795. [Google Scholar] [CrossRef]
- Liu, A.L.; Zou, J.; Liu, C.F.; Zhou, X.Y.; Zhang, X.W.; Luo, G.Y.; Chen, X.B. Over-expression of OsHsfA7 enhanced salt and drought tolerance in transgenic rice. BMB Rep. 2013, 46, 31–36. [Google Scholar] [CrossRef]
- Zhao, H.; Lou, Y.; Sun, H.; Li, L.; Wang, L.; Dong, L.; Gao, Z. Transcriptome and comparative gene expression analysis of Phyllostachys edulis in response to high light. BMC Plant Biol. 2016, 16, 34. [Google Scholar] [CrossRef]
- Pérez-Salamó, I.; Szabados, L. The heat shock factor A4A confers salt tolerance and is regulated by oxidative stress and the mitogen-activated protein kinases MPK3 and MPK6. Plant Physiol. 2014, 165, 319–334. [Google Scholar] [CrossRef]
- Xie, K.; Chen, J.; Wang, Q.; Yang, Y. Direct phosphorylation and activation of a mitogen-activated protein kinase by a calcium-dependent protein kinase in rice. Plant Cell 2014, 26, 3077–3089. [Google Scholar] [CrossRef]
- Shi, B.; Ni, L.; Liu, Y.; Zhang, A.; Tan, M.; Jiang, M. OsDMI3-mediated activation of OsMPK1 regulates the activities of antioxidant enzymes in abscisic acid signalling in rice. Plant Cell Environ. 2014, 37, 341–352. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene ID | Scaffolds | Number (aa) | MW (kDa) | PI | CELLO Localization (Reliability) |
|---|---|---|---|---|---|---|
| PheHsfA4a-1 | PH01000018G0140 | PH01000018 | 443 | 49.07 | 5.22 | Nuclear (3.717) |
| PheHsfB2c | PH01000314G0470 | PH01000314 | 382 | 40.12 | 4.86 | Nuclear (2.907) |
| PheHsfC1b-1 | PH01000333G0160 | PH01000333 | 247 | 26.77 | 8.71 | Cytoplasmic (1.219) |
| Mitochondrial (1.205) | ||||||
| Chloroplast (1.221) | ||||||
| PheHsfA1a | PH01000371G0730 | PH01000371 | 679 | 75.05 | 6.06 | Nuclear (2.682) |
| PheHsfB4a-1 | PH01000700G0690 | PH01000700 | 385 | 41.99 | 8.87 | Cytoplasmic (1.061) |
| Nuclear (1.481) | ||||||
| PheHsfC1b-2 | PH01000701G0030 | PH01000701 | 296 | 32.24 | 8.84 | Cytoplasmic (1.557) |
| PheHsfA4a-2 | PH01000746G0370 | PH01000746 | 444 | 49.50 | 5.36 | Nuclear (3.999) |
| PheHsfA4d-1 | PH01000814G0580 | PH01000814 | 554 | 61.26 | 4.95 | Nuclear (2.593) |
| PheHsfC2b | PH01000849G0330 | PH01000849 | 276 | 29.22 | 7.46 | Extracellular (1.087) |
| PheHsfA5 | PH01001018G0530 | PH01001018 | 499 | 55.33 | 5.21 | Nuclear (3.607) |
| PheHsfA7a | PH01001028G0180 | PH01001028 | 381 | 42.51 | 5.94 | Cytoplasmic (1.751) |
| PheHsfB4c-2 | PH01001228G0360 | PH01001228 | 276 | 29.72 | 8.34 | Nuclear (3.293) |
| PheHsfB4c-1 | PH01001554G0080 | PH01001554 | 420 | 45.70 | 9.81 | Nuclear (1.699) |
| PheHsfA4d-2 | PH01002437G0340 | PH01002437 | 453 | 51.34 | 5.18 | Nuclear (4.154) |
| PheHsfB4a-2 | PH01004959G0060 | PH01004959 | 275 | 30.24 | 8.37 | Chloroplast (1.119) |
| Nuclear (1.604) | ||||||
| PheHsfB2a | PH01000000G3800 | PH01000000 | 292 | 31.22 | 4.98 | Nuclear (3.984) |
| PheHsfA6b-2 | PH01000081G0140 | PH01000081 | 362 | 41.44 | 4.97 | Nuclear (3.466) |
| PheHsfA2a-2 | PH01000174G0590 | PH01000174 | 342 | 38.47 | 5.32 | Nuclear (4.565) |
| PheHsfA6a | PH01000194G0800 | PH01000194 | 378 | 42.59 | 4.70 | Nuclear (1.785) |
| PheHsfA6b-1 | PH01000208G0690 | PH01000208 | 348 | 40.04 | 4.89 | Cytoplasmic (2.207) |
| Nuclear (2.182) | ||||||
| PheHsfA2a-1 | PH01003916G0010 | PH01003916 | 358 | 40.31 | 5.49 | Nuclear (3.869) |
| PheHsfB1 | PH01000149G1320 | PH01000149 | 297 | 32.42 | 8.55 | Nuclear (3.609) |
| Gene | DBD | HR-A/B | NLS | AHA | RD | NES |
|---|---|---|---|---|---|---|
| PheHsfA1a | 1–84 | 121–171 | (201) ANKKRRLPKQ | (410) SFWEQFLVA | nd | nd |
| PheHsfA2a-1 | 39–128 | 151–202 | (237) ISKKRRRRID | (314) DDFWEDLLHE | nd | nd |
| PheHsfA2a-2 | 40–129 | 156–197 | (238) ISKKRRRRID | (315) DDFWEDLLHE | nd | (165) LLM |
| PheHsfA4a-1 | 14–103 | 124–165 | (204) DHHRKKRRLPKPISF | (379) GFWQQFLTE | nd | nd |
| PheHsfA4a-2 | 14–103 | 124–172 | (204) DHHRKKRRLPKPISF | (380) GFWQQFLTE | nd | nd |
| PheHsfA4d-1 | 13–96 | 112–140 | (199) FSKKRRAPKI | (363) LFWERFLTE | nd | nd |
| PheHsfA4d-2 | 16–105 | 120–149 | (208) FSKKRRVPKI | (382) LFWERFLTE | nd | (252) MELAL |
| PheHsfA5 | 30–134 | 152–179 | (241) FQKKRRLTGL | (446) KFWEQFLTE | nd | nd |
| PheHsfA6a | 46–135 | 165–206 | (247) ISKKRRRPID | (319) DDFWAELLVE | nd | (288) LENLAL |
| PheHsfA6b-1 | 45–134 | 163–204 | (133) LLKMIKRRRLLYY | (318) DDFWEELLNE | nd | nd |
| PheHsfA6b-2 | 45–134 | 163–204 | (135) KMIKRRRPLS | (318) EDFWEELLNE | nd | nd |
| PheHsfA7a | 74–165 | 193–227 | (166) KNIKRRRASK | (329) DDVWEELDAI | nd | nd |
| PheHsfB1 | 30–117 | 176–205 | (253) EDATRKRKRCEEAAARERPFKMIRI | nd | (246) KLFGVLL | nd |
| PheHsfB2a | 7–100 | 160–183 | nd | nd | (203) TLFGVTI | (264) LDVLALSL |
| PheHsfB2c | 29–118 | 199–232 | nd | nd | (304) RLFGVSI | nd |
| PheHsfB4a-1 | 19–112 | 110–139 | (237) RKRLLQEQPPTSPEWKRSMV | nd | (215) KLFGVNL | nd |
| PheHsfB4a-2 | 19–103 | 201–230 | (347) RKRSLQEQPPTSPDWKRSMV | nd | (325) KLFGVDL | nd |
| PheHsfB4c-1 | 26–116 | 256–285 | nd | nd | (323) KLFGVHI | nd |
| PheHsfB4c-2 | 26–116 | 124–143 | nd | nd | (213) KLFGVHL | (251) LESDDLSL |
| PheHsfC1b-1 | 25–114 | 136–165 | (209) PGKKRRIGAE | nd | nd | nd |
| PheHsfC1b-2 | 72–161 | 186–213 | (257) TPGKRRRIG | nd | nd | nd |
| PheHsfC2b | 12–105 | 140–176 | nd | nd | nd | (173) LKV |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, L.; Li, X.; Hou, D.; Cheng, Z.; Liu, J.; Li, J.; Mu, S.; Gao, J. Genome-Wide Analysis and Expression Profiling of the Heat Shock Factor Gene Family in Phyllostachys edulis during Development and in Response to Abiotic Stresses. Forests 2019, 10, 100. https://doi.org/10.3390/f10020100
Xie L, Li X, Hou D, Cheng Z, Liu J, Li J, Mu S, Gao J. Genome-Wide Analysis and Expression Profiling of the Heat Shock Factor Gene Family in Phyllostachys edulis during Development and in Response to Abiotic Stresses. Forests. 2019; 10(2):100. https://doi.org/10.3390/f10020100
Chicago/Turabian StyleXie, Lihua, Xiangyu Li, Dan Hou, Zhanchao Cheng, Jun Liu, Juan Li, Shaohua Mu, and Jian Gao. 2019. "Genome-Wide Analysis and Expression Profiling of the Heat Shock Factor Gene Family in Phyllostachys edulis during Development and in Response to Abiotic Stresses" Forests 10, no. 2: 100. https://doi.org/10.3390/f10020100
APA StyleXie, L., Li, X., Hou, D., Cheng, Z., Liu, J., Li, J., Mu, S., & Gao, J. (2019). Genome-Wide Analysis and Expression Profiling of the Heat Shock Factor Gene Family in Phyllostachys edulis during Development and in Response to Abiotic Stresses. Forests, 10(2), 100. https://doi.org/10.3390/f10020100