Fire Survival of Longleaf Pine (Pinus palustris) Grass Stage Seedlings: The Role of Seedling Size, Root Collar Position, and Resprouting

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. The Studied Seedling Population
2.3. Data Collection
2.4. Data Analyses
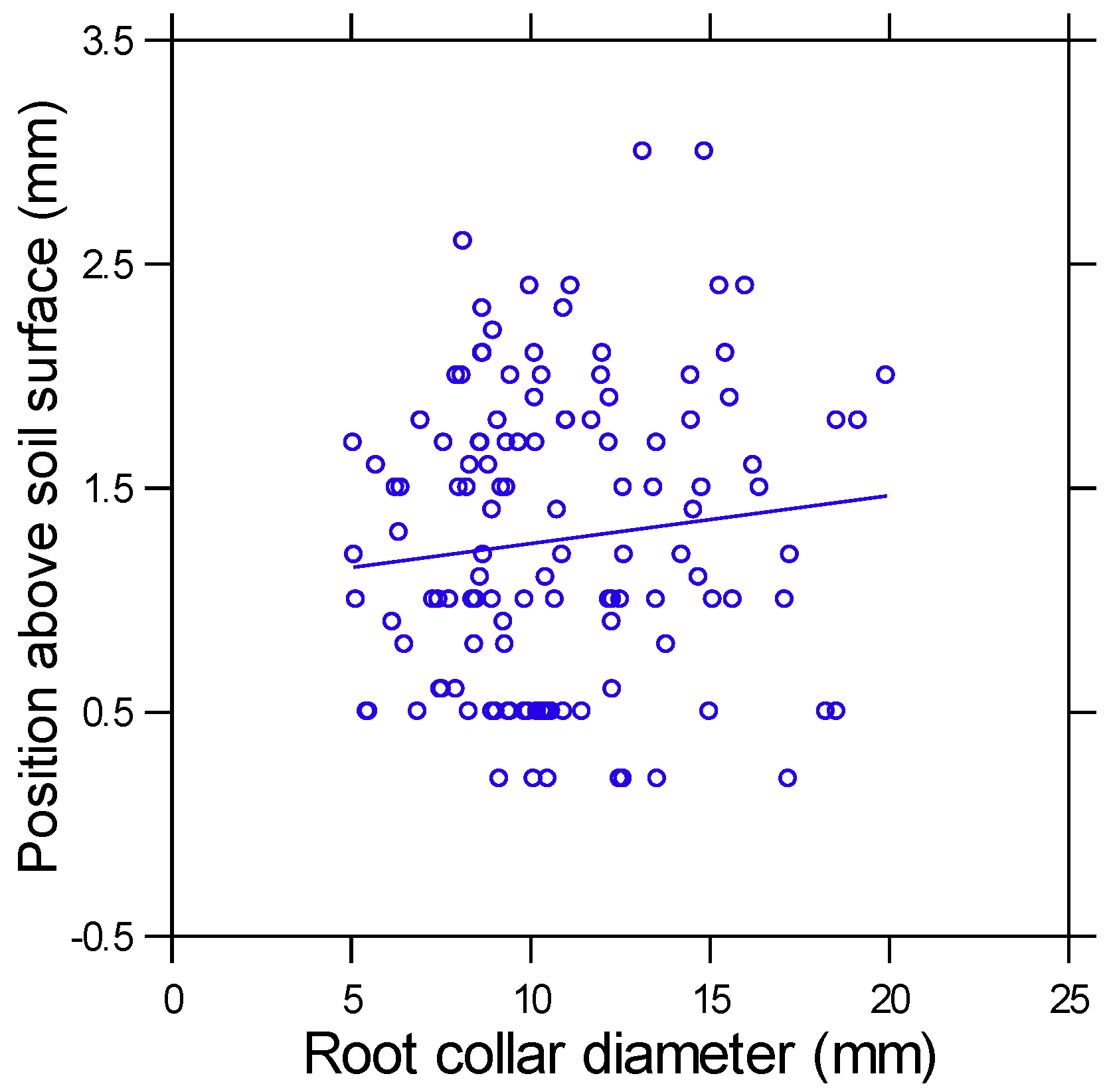
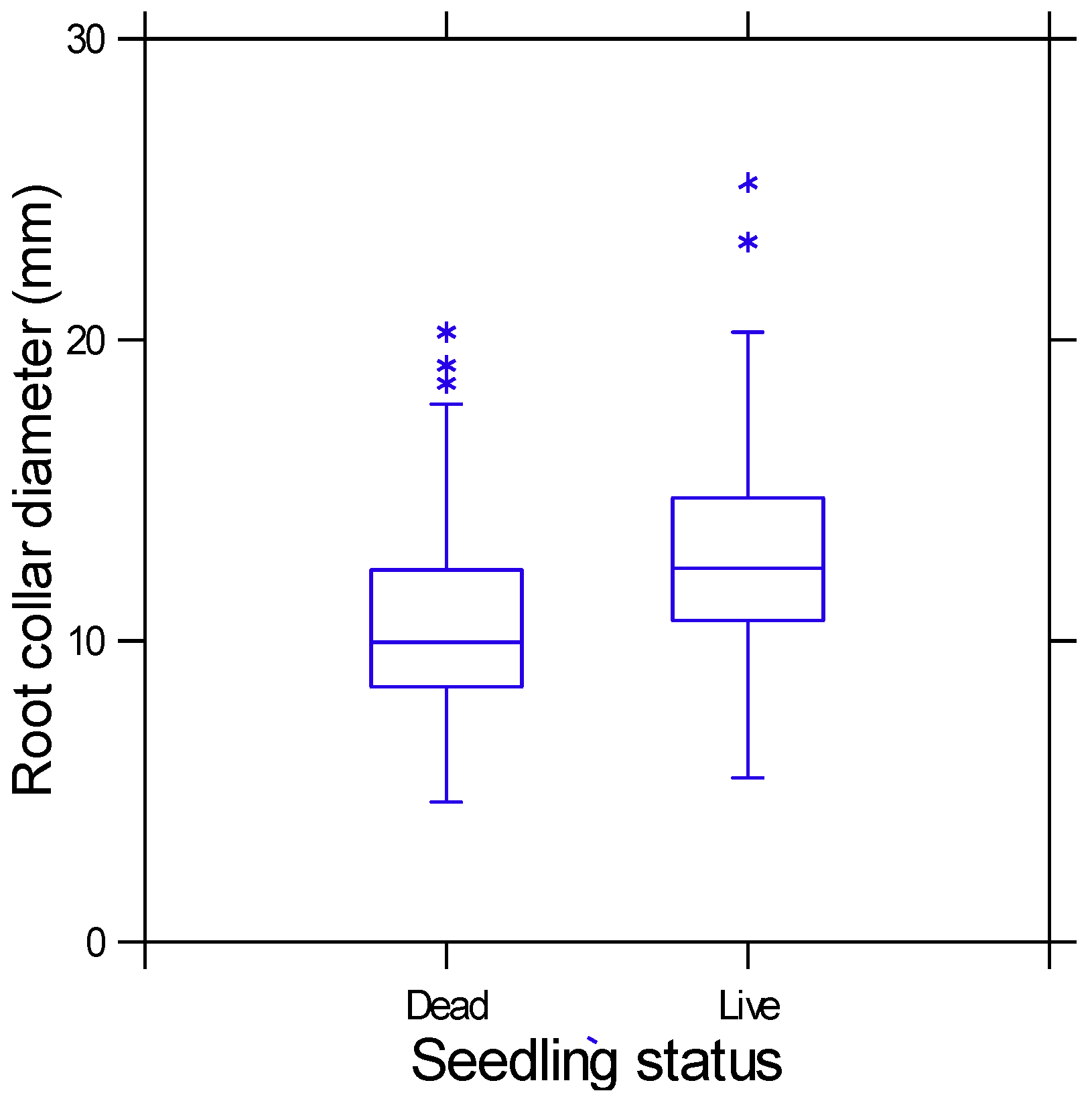
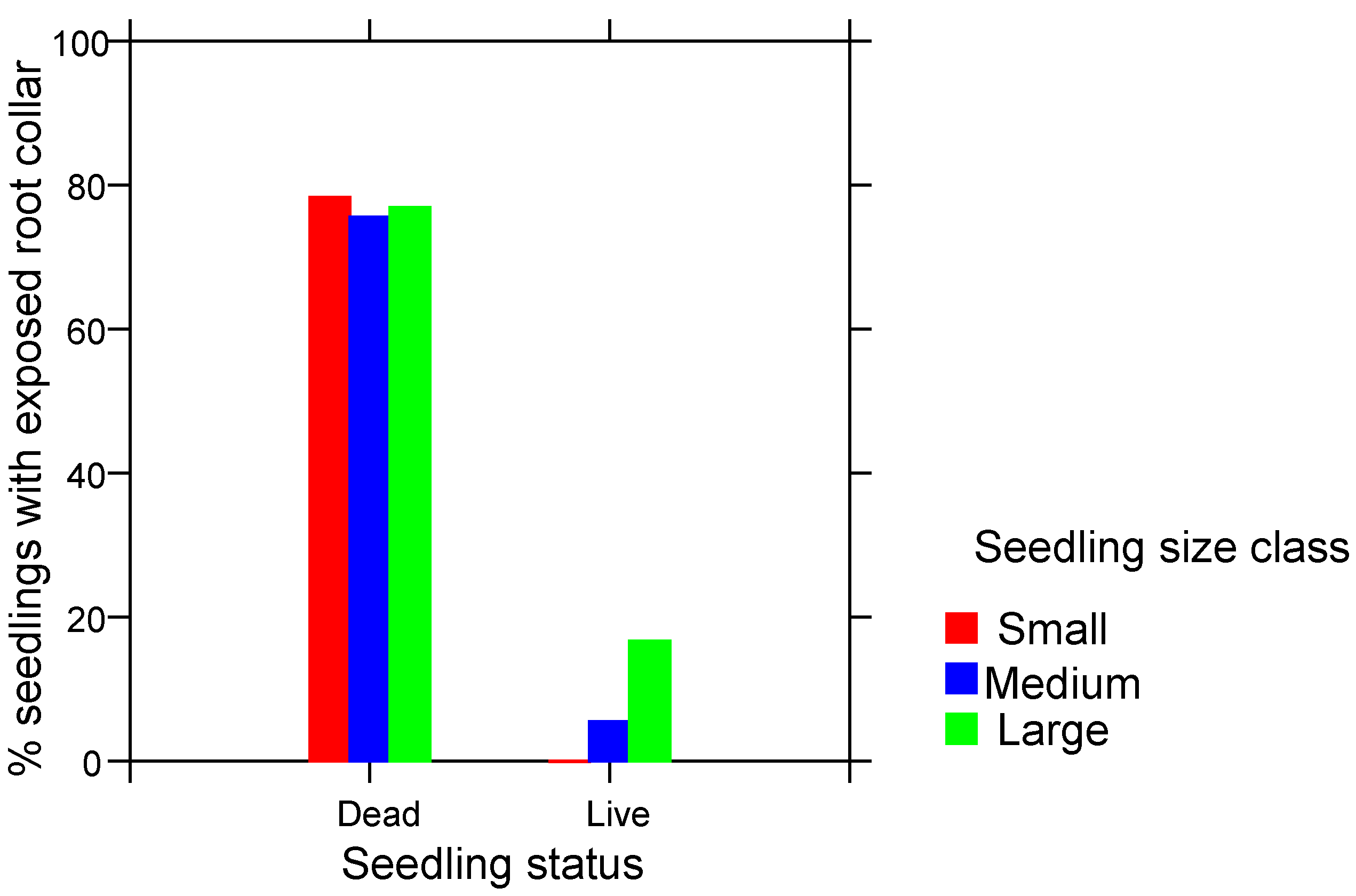
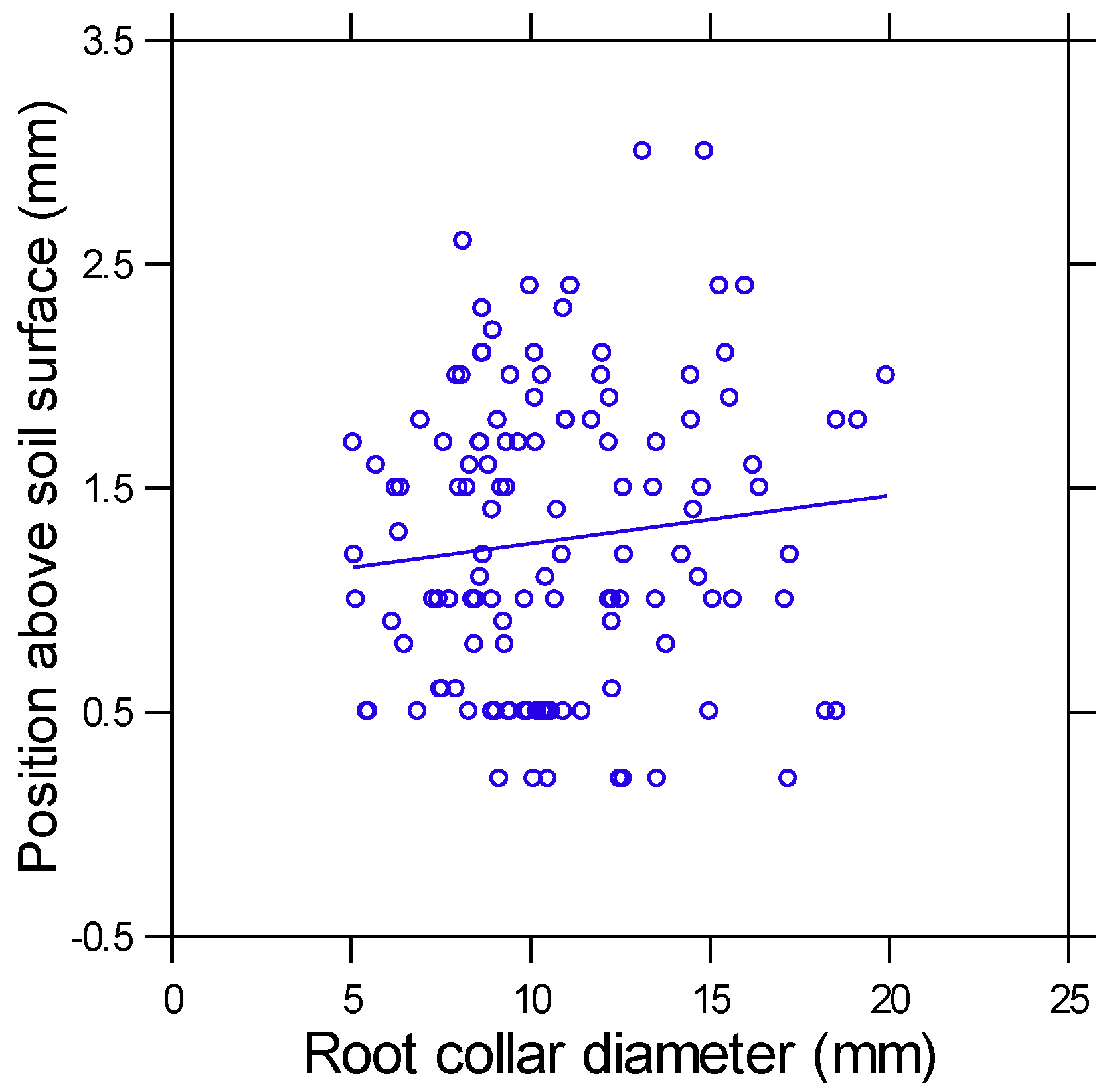
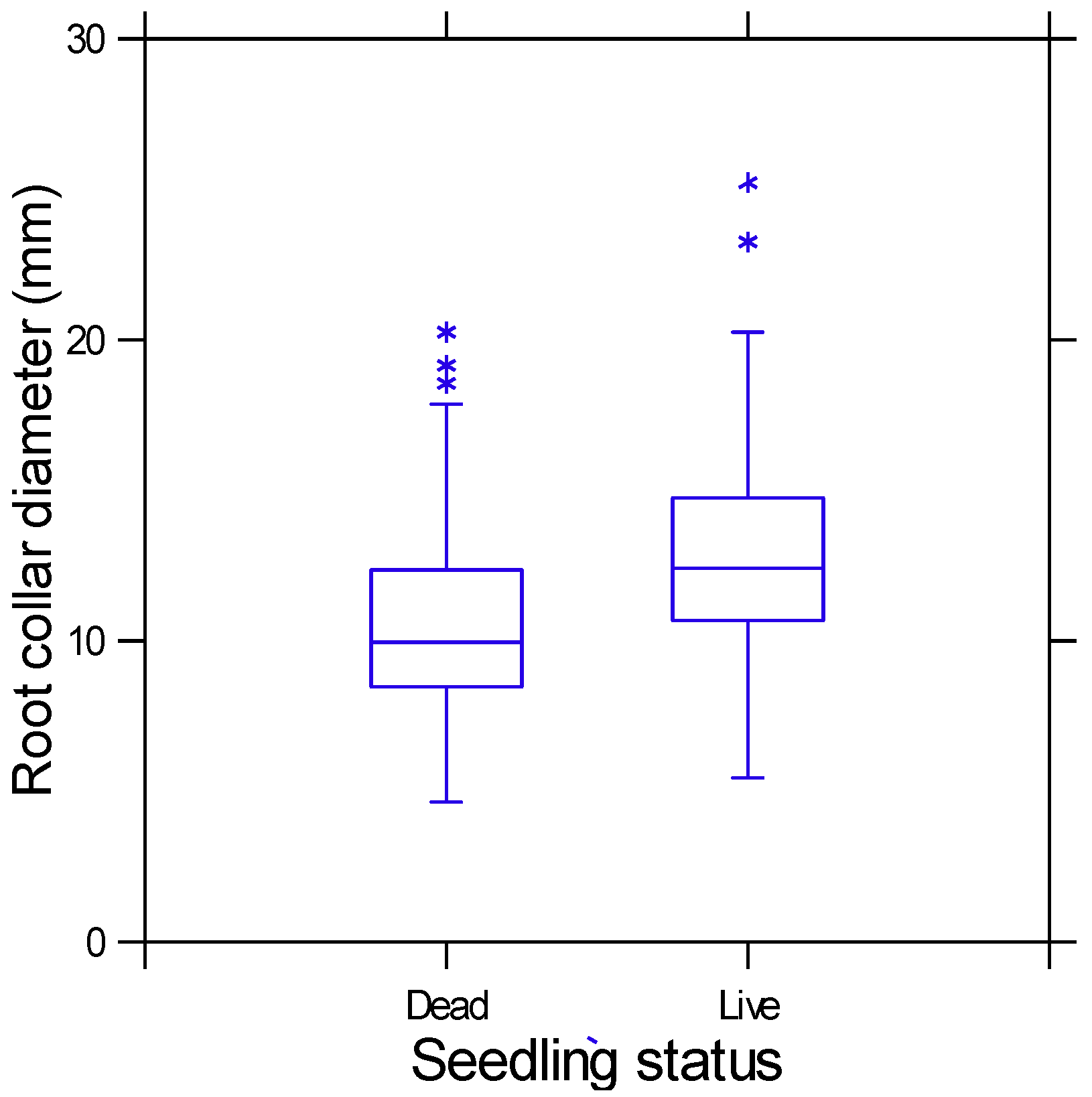
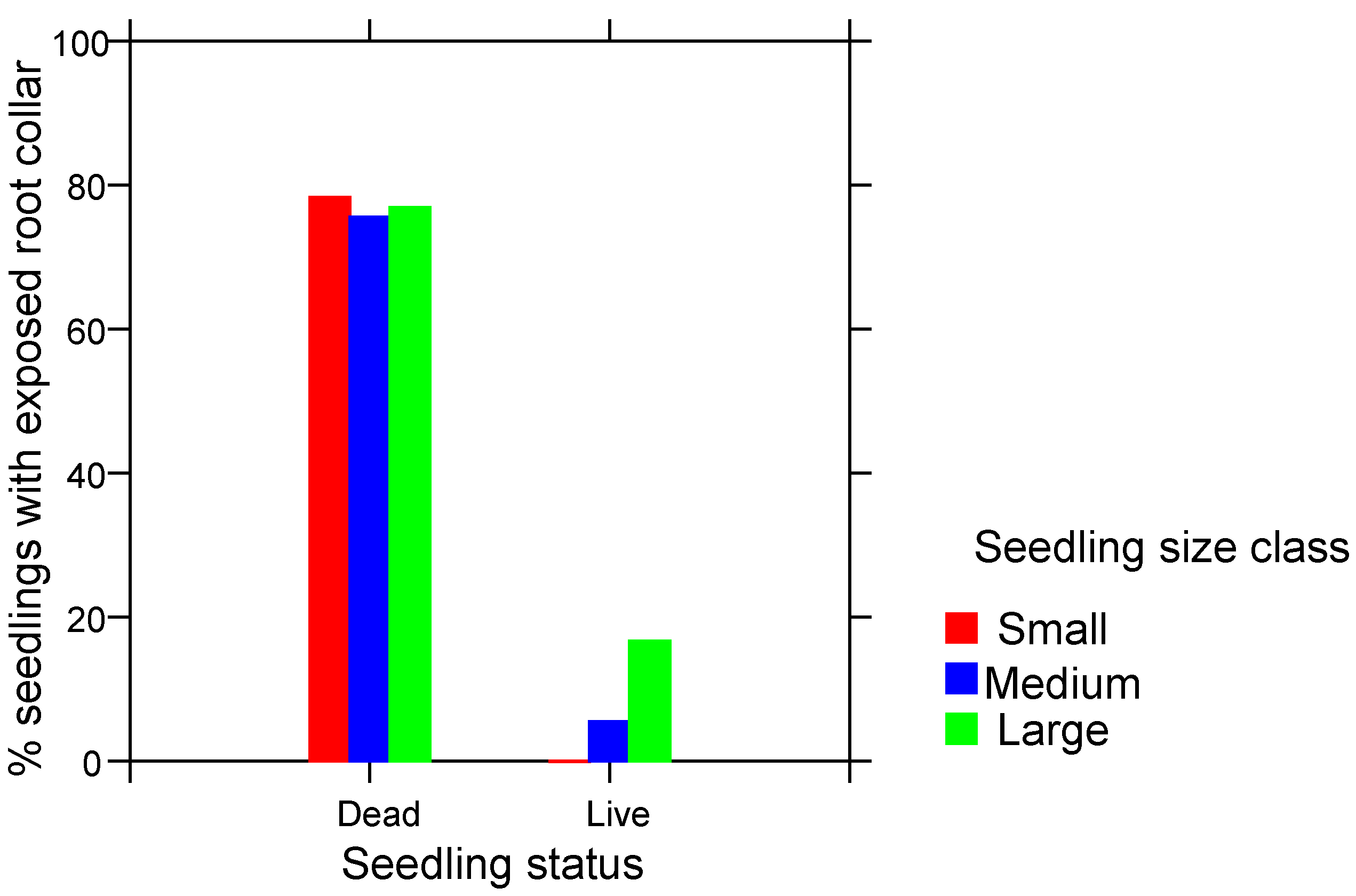
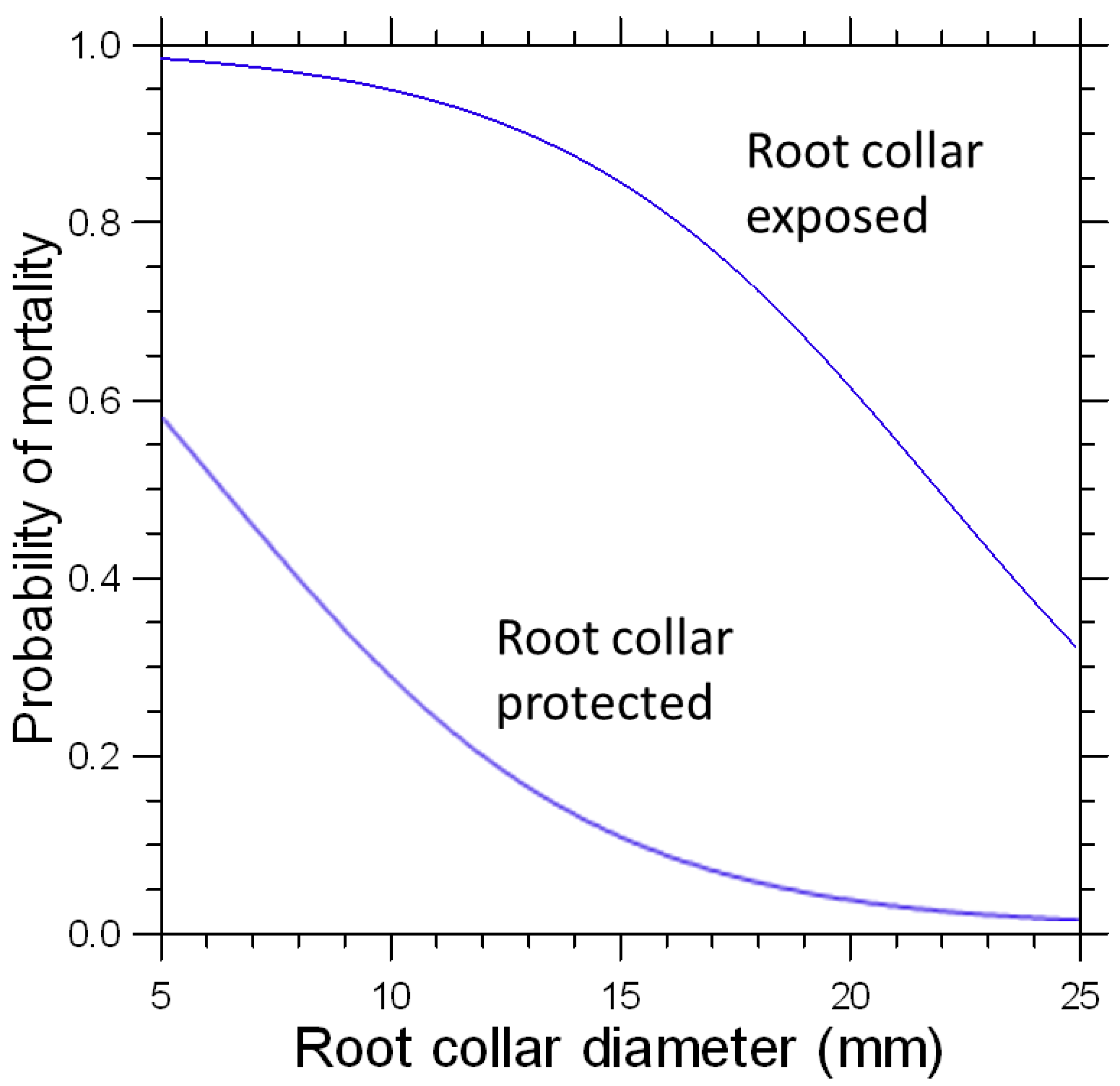
3. Results
4. Discussion
4.1. Seedling Size and Fire Survival
4.2. Root Collar Position and Fire Survival
4.3. Sprouting Ability and Fire Survival
5. Conclusions and Management Implication
Author Contributions
Funding
Conflicts of Interest
References
- Wahlenberg, W.G. Longleaf Pine: Its Use, Ecology, Regeneration, Protection, Growth, and Management; Charles Lathrop Pack Forestry Foundation and USDA Forest Service: Washington, DC, USA, 1946; p. 429. [Google Scholar]
- Frost, C.C. Four centuries of changing landscape patterns in the longleaf pine ecosystem. In Proceedings of the Tall Timbers Fire Ecology Conference; Hermann, S.M., Ed.; Tall Timbers Research Station: Tallahassee, FL, USA, 1993; Volume 18, pp. 17–43. [Google Scholar]
- Frost, C.C. History and future of the longleaf pine ecosystem. In The Longleaf Pine Ecosystem: Ecology, Silviculture, and Restoration; Jose, S., Jokela, E.J., Miller, D.L., Eds.; Springer: New York, NY, USA, 2006; pp. 9–42. [Google Scholar]
- Boyer, W.D. Pinus palustris Mill. Longleaf pine. In Silvics of North America; Burns, R.M., Honkala, B.H., Eds.; Conifers. USDA Handbook 654; USDA: Washington, DC, USA, 1990; Volume 1, pp. 405–412. [Google Scholar]
- van Lear, D.H.; Carroll, W.; Kapeluck, P.; Johnson, R. History and restoration of the longleaf pine-grassland ecosystem: Implications for species at risk. For. Ecol. Manag. 2005, 211, 150–165. [Google Scholar] [CrossRef]
- Mirov, N.T. The Genus Pinus; The Ronald Press: New York, NY, USA, 1967; p. 602. [Google Scholar]
- Knapp, B.O.; Wang, G.G.; Walker, J.L.; Cohen, S. Effects of site preparation treatments on early growth and survival of planted longleaf pine (Pinus palustris Mill.) seedlings in North Carolina. For. Ecol. Manag. 2006, 226, 122–128. [Google Scholar] [CrossRef]
- Croker, T.C.; Boyer, W.D. Regenerating Longleaf Pine Naturally; Research Paper SO-105; USDA Forest Service, Southern Forest Experiment Station: New Orleans, LA, USA, 1975.
- Maple, W.R. Mortality of Longleaf Pine Seedlings Following a Winter Burn against Brown-Spot Needle Blight; Research Note SO-195; U.S. Department of Agriculture, Forest Service, Southern Forest Experiment Station: New Orleans, LA, USA, 1975; p. 3.
- Boyer, W.D. Regenerating Longleaf Pine with Natural Seeding. In Proceedings of the Tall Timbers Fire Ecology Conference, No. 18, The Longleaf Pine Ecosystem: Ecology, Restoration and Management; Hermann, S.M., Ed.; Tall Timbers Research Station: Tallahassee, FL, USA, 1993. [Google Scholar]
- Brockway, D.G.; Outcalt, K.W.; Boyer, W.D. Longleaf pine regeneration ecology and methods. In The Longleaf Pine Ecosystem: Ecology, Silviculture, and Restoration; Shibu, J., Jokela, E.J., Miller, D.L., Eds.; Springer: New York, NY, USA, 2006; pp. 95–133. [Google Scholar]
- Pile, L.S.; Wang, G.G.; Knapp, B.O.; Liu, G.; Yu, D. Morphological and physiological adaptations to frequent surface fire regimes: A comparative study on seedlings of three Pinus species native to the southeast United States. Ann. For. Sci. 2017, 74, 68. [Google Scholar] [CrossRef]
- Wang, G.G.; Pile, L.S.; Knapp, B.O.; Hu, H. Longleaf pine adaptation to fire: Is early height growth pattern critical to fire survival? In Proceedings of the 18th Southern Silvicultural Research Conference, Knoxville, TN, USA, 2–5 March 2015. [Google Scholar]
- Kush, J.S. Natural Regeneration of Longleaf Pine: Adaptations to Site Conditions and Management Systems. Ph.D. Thesis, Auburn University, Auburn, AL, USA, 2002. [Google Scholar]
- Moule, B.M. How Mechanical Manipulation and Herbicide Treatments Measure up to the Effects of Prescribed Fire on an Established Longleaf Pine (Pinus palustris) Ecosystem; Clemson University: Clemson, SC, USA, 2013; p. 221. [Google Scholar]
- Knapp, B.O.; Pile, L.S.; Walker, J.L.; Wang, G.G. Fire effects on a fire-adapted species: Response of grass stage longleaf pine seedlings to experimental burning. Fire Ecol. 2018, 14, 2. [Google Scholar] [CrossRef]
- Bruce, D. Fire resistance of pine seedlings. J. For. 1951, 49, 739–740. [Google Scholar]
- Bruce, D. Mortality of pine seedlings after a winter fire. J. For. 1954, 52, 442–443. [Google Scholar]
- Grelen, H.E. May burning favors survival and early height growth of longleaf pine seedlings. South. J. Appl. For. 1983, 7, 16–20. [Google Scholar] [CrossRef]
- Maple, W.R. Prescribed Winter Fire Thins Dense Longleaf Seedling Stand; Research Note SO-104; U.S. Department of Agriculture, Forest Service, Southern Forest Experiment Station: New Orleans, LA, USA, 1970; p. 2.
- Jack, S.B.; Hiers, J.K.; Mitchell, R.J.; Gagnon, J.L. Fuel loading and fire intensity—Effects on longleaf pine seedling survival. In Proceedings of the 14th Biennial Southern Silvicultural Research Conference; Stanturf, J.A., Ed.; Gen. Tech. Rep. SRS-121; U.S. Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2010; pp. 275–279. [Google Scholar]
- Hiers, J.K.; O’Brien, J.J.; Mitchell, R.J.; Grego, J.M.; Loudermilk, E.L. The wildland fuel cell concept: An approach to characterize fine-scale variation in fuels and fire in frequently burned longleaf pine forests. Int. J. Wildland Fire 2009, 18, 315–325. [Google Scholar] [CrossRef]
- Wenk, E.S.; Wang, G.G.; Walker, J.L. Within-stand variation in understory vegetation affects fire behavior in longleaf pine xeric sandhills. Int. J. Wildland Fire 2011, 20, 866–875. [Google Scholar] [CrossRef]
- Bigelow, S.W.; Whelan, A.W. Longleaf pine proximity effects on air temperatures and hardwood top-kill from prescribed fire. Fire Ecol. 2019, 15. [Google Scholar] [CrossRef]
- Robertson, K.M.; Platt, W.J.; Faires, C.E. Patchy fires promote regeneration of longleaf pine (Pinus palustris Mill.) in pine savannas. Forests 2019, 10, 367. [Google Scholar] [CrossRef]
- Grace, S.L.; Platt, W.J. Effects of adult tree density and fire on the demography of pregrass stage juvenile longleaf pine (Pinus palustris Mill.). J. Ecol. 1995, 83, 75–86. [Google Scholar] [CrossRef]
- Schwarz, G.F. The Longleaf Pine in Virgin Forests: A Silvical Study; John Wiley and Sons: New York, NY, USA, 1907; p. 135. [Google Scholar]
- Stone, E.L.; Stone, M.H. Root collar sprouts in pine. J. For. 1954, 52, 487–491. [Google Scholar]
- Garin, G.I. Longleaf pines can form vigorous sprouts. J. For. 1958, 56, 430–431. [Google Scholar]
- Farrar, R.M., Jr. Sprouting ability of longleaf pine. For. Sci. 1975, 21, 189–190. [Google Scholar]
- Schafale, M.P. Guide to the Natural Communities of North Carolina, Fourth Approximation. NC Department of Natural Resources, Natural Heritage Program. 2012. Available online: http://cvs.bio.unc.edu/pubs/4thApproximationGuideFinalMarch2012.pdf (accessed on 10 October 2019).
- The United States Department of Agriculture (USDA). Soil Survey of Aikens County Area South Carolina; USDA Soil Conservation Service: Washington, DC, USA, 1985; p. 134.
- Southeast Regional Climate Center. Aiken 4 NE, South Carolina (380074): Period of Record Monthly Climate Summary. 2011. Available online: http://www.sercc.com/cgi-bin/sercc/cliMAIN.pl?sc0074 (accessed on 8 May 2017).
- Pessin, L.J. Annual ring formation in Pinus palustris seedlings. Am. J. Bot. 1934, 21, 599–603. [Google Scholar] [CrossRef]
- SYSTAT Software Inc. SYSTAT 13: Statistics II; SYSTAT Software Inc.: Chicago, IL, USA, 2009. [Google Scholar]
- Knapp, B.O.; Wang, G.G.; Hu, H.; Walker, J.L.; Tennant, C. Restoring longleaf pine (Pinus palustris Mill.) in loblolly pine (Pinus taeda L.) stands: Effects of restoration treatments on natural loblolly pine regeneration. For. Ecol. Manag. 2011, 262, 1157–1167. [Google Scholar] [CrossRef]
- Loudermilk, E.L.; Hiers, J.K.; Pokswinski, S.; O’Brien, J.J.; Barnett, A.; Mitchell, R.J. The path back: Oaks (Quercus spp.) facilitate longleaf pine (Pinus palustris) seedling establishment in xeric sites. Ecosphere 2016, 7, e01361. [Google Scholar] [CrossRef]
- Planting Depth is Critical. Available online: https://longleafalliance.org/what-we-do/restoration-management/restoration/planting-longleaf-pine-seedlings/planting-depth-is-critical (accessed on 10 October 2019).
- Sayer, M.A.S.; Tyree, M.C.; Dillaway, D.N.; Rudd, B.M. Foliage re-establishment of Pinus palustris Mill. Saplings after spring or fall prescribed fire. New For. 2018, 49, 851–869. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Mean | Minimum | Maximum |
|---|---|---|---|
| Temperature (°F) | 57 | 41 | 73 |
| Humidity (%) | 58 | 35 | 82 |
| Wind speed (MPH) | 5 | 0 | 10 |
| Dew point (°F) | 40 | 34 | 48 |
| Precipitation (mm) | 0 | - | - |
| of Seedlings | Mortality (%) | |
|---|---|---|
| Size Class | ||
| RCD < 7.6 mm | 30 | 76.7% |
| 7.6 mm > RCD < 15 mm | 225 | 51.1% |
| RCD ≥ 15 mm | 43 | 30.2% |
| Root Collar Exposure Class | ||
| Exposed | 126 | 91.3% |
| Protected | 172 | 20.9% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, S.; Moule, B.; Yu, D.; Wang, G.G. Fire Survival of Longleaf Pine (Pinus palustris) Grass Stage Seedlings: The Role of Seedling Size, Root Collar Position, and Resprouting. Forests 2019, 10, 1070. https://doi.org/10.3390/f10121070
Jin S, Moule B, Yu D, Wang GG. Fire Survival of Longleaf Pine (Pinus palustris) Grass Stage Seedlings: The Role of Seedling Size, Root Collar Position, and Resprouting. Forests. 2019; 10(12):1070. https://doi.org/10.3390/f10121070
Chicago/Turabian StyleJin, Songheng, Brett Moule, Dapao Yu, and G. Geoff Wang. 2019. "Fire Survival of Longleaf Pine (Pinus palustris) Grass Stage Seedlings: The Role of Seedling Size, Root Collar Position, and Resprouting" Forests 10, no. 12: 1070. https://doi.org/10.3390/f10121070
APA StyleJin, S., Moule, B., Yu, D., & Wang, G. G. (2019). Fire Survival of Longleaf Pine (Pinus palustris) Grass Stage Seedlings: The Role of Seedling Size, Root Collar Position, and Resprouting. Forests, 10(12), 1070. https://doi.org/10.3390/f10121070