Seven Decades of Spontaneous Forest Regeneration after Large-Scale Clear-Cutting in Białowieża Forest do not Ensure the Complete Recovery of Collembolan Assemblages



Abstract
1. Introduction
2. Materials and Methods
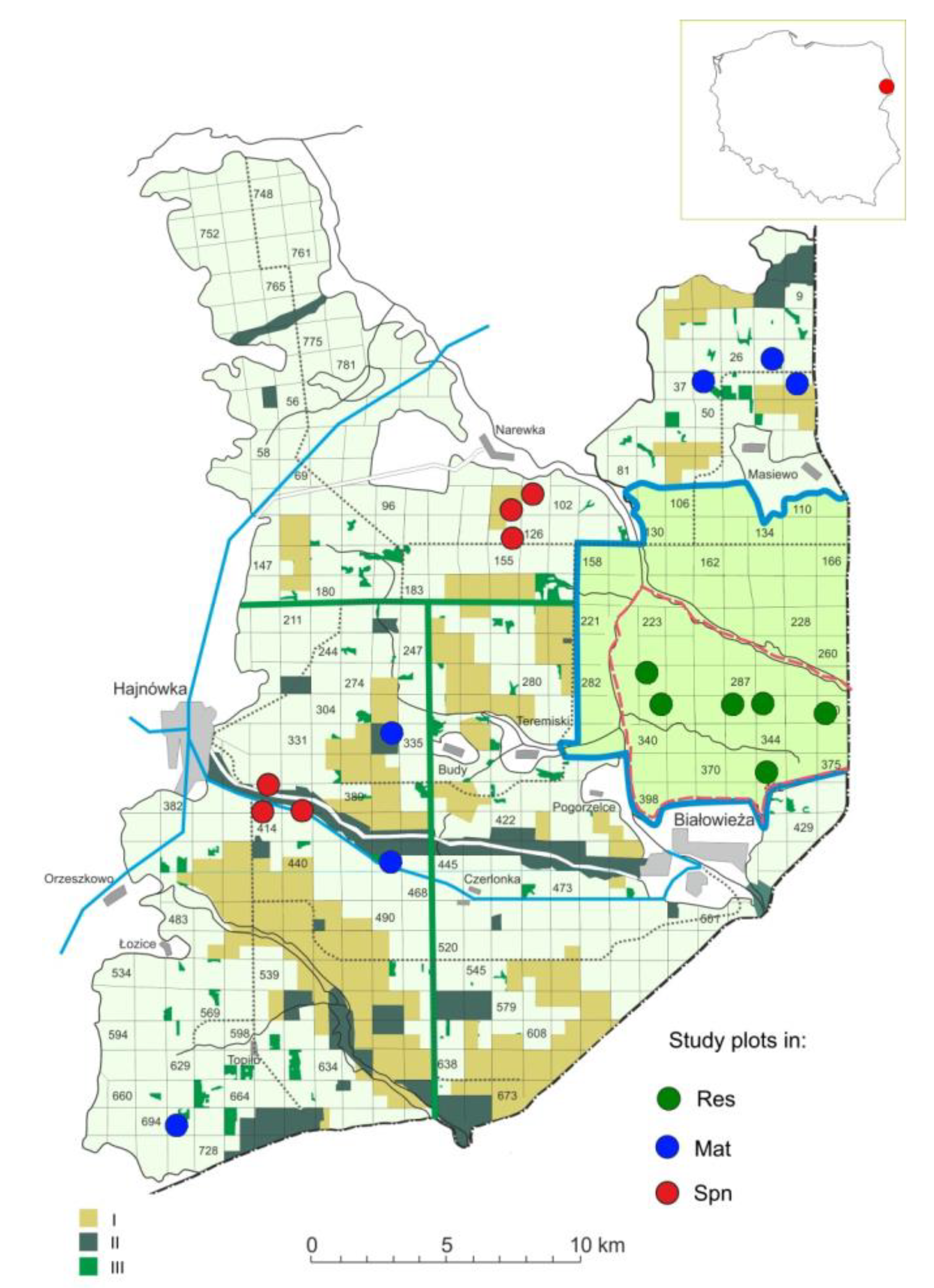
2.1. Study Area
2.2. Data Collection and Analysis
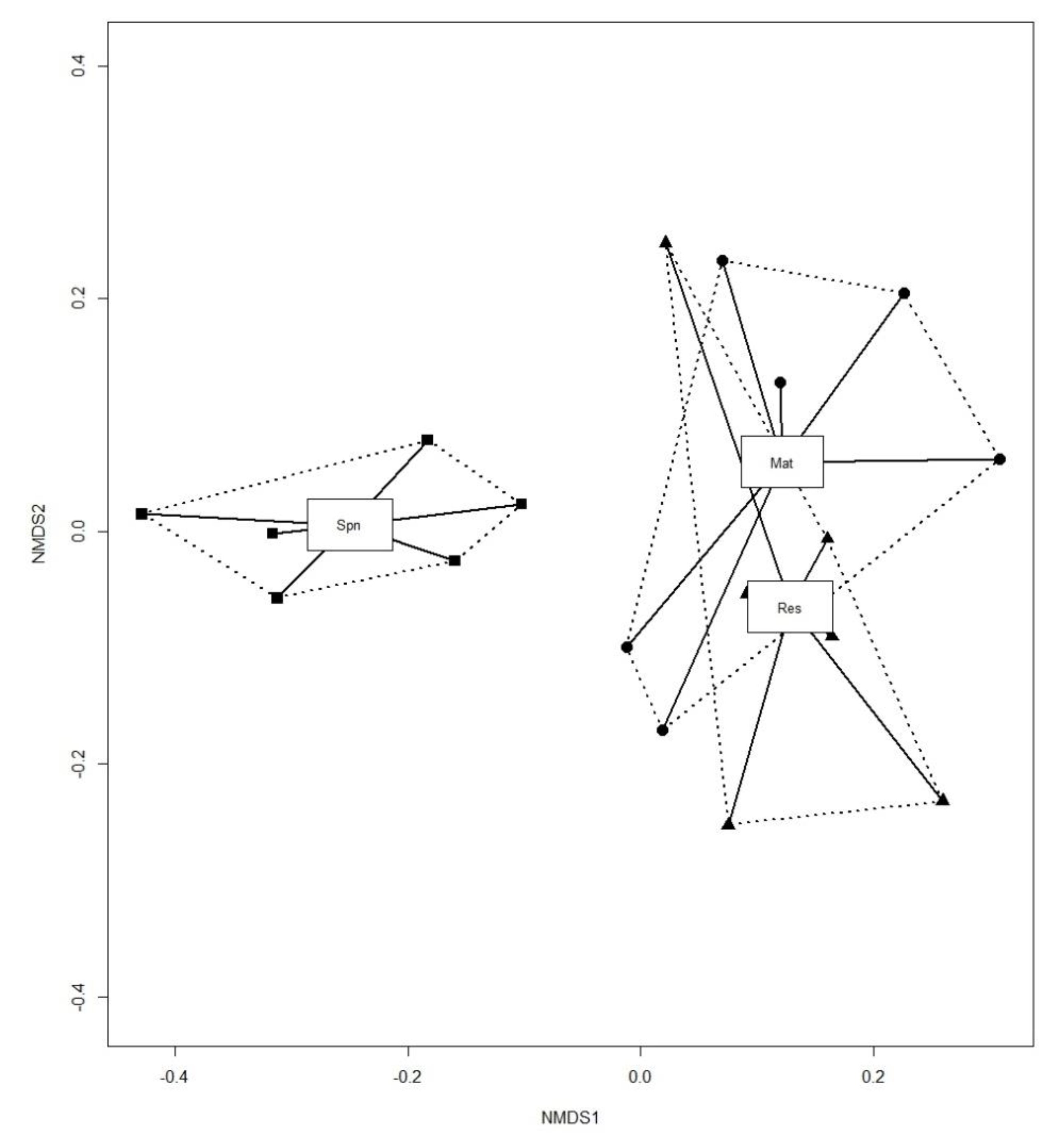
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix
| Taxa | Life Form | Spn | Mat | Res | Total |
| Ceratophysella engadinensis (Gisin, 1949) | ep | 1 | 56 | 40 | 97 |
| C. mosquensis (Becker, 1905) | ep | - | 1 | - | 1 |
| Ceratophysella sp. juv. | ep | 3 | 13 | 2 | 18 |
| Xenylla brevicauda (Tullberg, 1869) | h | 7 | - | - | 7 |
| Xenylla sp. juv. | h | 25 | - | - | 25 |
| Willemia anopthalma (Börner, 1901) | eu | 1 | - | 56 | 57 |
| W. denisi (Mills, 1932) sensu (Fjellberg 1985) | eu | - | 8 | 28 | 36 |
| Willemia sp. juv. | eu | - | 1 | 2 | 3 |
| Xenyllodes armatus (Axelson, 1903) | h | - | 1 | - | 1 |
| Friesea claviseta (Axelson, 1900) | ep | - | - | 56 | 56 |
| Friesea sp. juv. | ep | - | - | 1 | 1 |
| Pseudachorutes corticicolus (Schäffer, 1896) | ep | - | 3 | 10 | 13 |
| P. dubius (Krausbauer, 1898) | ep | 1 | - | - | 1 |
| P. parvulus (Börner, 1901) | ep | 1 | 1 | 11 | 13 |
| Pseudachorutes sp. juv. | ep | 3 | - | 3 | 6 |
| Micranurida pygmea (Börner, 1901) | h | 1 | - | 9 | 10 |
| Anurida granulata (Agrell, 1943) | h | - | 4 | 3 | 7 |
| Neanura muscorum (Templeton, 1835) | h | 23 | 5 | 9 | 37 |
| Micraphorura absoloni (Börner, 1901) | eu | 5 | 9 | 40 | 54 |
| Protaphorura bicampata (Gisin, 1956) | eu | - | 16 | - | 16 |
| P. pannonica (Haybach, 1960) | eu | 55 | 17 | 54 | 126 |
| P. subarmata (Gisin, 1957) | eu | - | 67 | 67 | 134 |
| P. tricampata (Gisin, 1956) | eu | 7 | 1 | - | 8 |
| Protaphorura sp. juv. | eu | 16 | 11 | 58 | 85 |
| Supraphorura furcifera (Börner, 1901) | eu | - | 11 | - | 11 |
| Hymenaphorura polonica (Pomorski, 1990) | eu | - | - | 25 | 25 |
| Mesaphorura critica (Ellis, 1976) | eu | - | 1 | 10 | 11 |
| M. hylophila (Rusek, 1982) | eu | 3 | 9 | 7 | 19 |
| M. italica (Rusek, 1971) | eu | - | - | 3 | 3 |
| M. macrochaeta (Rusek, 1976) | eu | 65 | 54 | 91 | 210 |
| M. sylvatica (Rusek, 1971) | eu | - | - | 1 | 1 |
| M. tenuisensillata (Rusek, 1974) | eu | - | 7 | 12 | 19 |
| M. yosii (Rusek, 1967) | eu | - | 6 | 35 | 41 |
| Mesaphorura sp. juv. | eu | 1 | 1 | 9 | 11 |
| Karlstejnia norvegica (Fjellberg, 1974) | eu | 1 | - | 1 | 2 |
| Stenaphorurella quadrispina (Börner, 1901) | eu | 1 | - | - | 1 |
| Anurophorus septentrionalis (Pallisa, 1966) | h | - | - | 30 | 30 |
| Anurophorus sp. juv. | h | 1 | - | 33 | 34 |
| Folsomia dovrensis (Fjellberg, 1976) | eu | - | 1 | 1 | 2 |
| F. fimetarioides (Axelson, 1903) | eu | - | 73 | - | 73 |
| F. lawrencei (Rusek, 1984) | eu | 4 | - | - | 4 |
| F. manolachei (Bagnal, 1939) | h | 10 | - | - | 10 |
| F. stella (Christansen & Tucker, 1977) | eu | - | 1 | 9 | 10 |
| F. quadrioculata (Tullberg, 1871) | h | 228 | 420 | 631 | 1279 |
| Appendisotoma juliannae (Traser, 1993) | ep | - | - | 190 | 190 |
| Proisotoma armeriae (Fjellberg, 1976) | h | - | 1 | - | 1 |
| P. minima (Tullberg, 1871) | h | 4 | 5 | 31 | 40 |
| Proisotoma sp. juv. | h | - | - | 1 | 1 |
| Isotomiella minor (Schäffer, 1896) | eu | 509 | 339 | 893 | 1741 |
| Parisotoma notabilis (Schäffer, 1896) | h | 970 | 310 | 323 | 1603 |
| Vertagopus sp. juv. | ep | - | - | 1 | 1 |
| Isotoma viridis (Bourlet, 1839) | ep | - | 41 | - | 41 |
| Isotoma sp. juv. | ep | 2 | 11 | 2 | 15 |
| Desoria divergens (Axelson, 1900) | ep | 8 | - | 12 | 20 |
| D. tigrina (Tullberg, 1871) | ep | - | 33 | 13 | 46 |
| D. violacea (Tullberg, 1876) | ep | - | 44 | 12 | 56 |
| Isotomidae juv. | ep | - | - | 30 | 30 |
| Tomocerus vulgaris (Tullberg, 1871) | ep | - | 1 | 2 | 3 |
| Pogonognathellus flavescens (Tullberg, 1871) | ep | 4 | 99 | 47 | 150 |
| Tomoceridae juv. | ep | 2 | 12 | 4 | 18 |
| Orchesella bifasciata (Nicolet, 1841) | a | 2 | - | - | 2 |
| O. flavescens (Bourlet, 1839) | a | 9 | 55 | 10 | 74 |
| Orchesella sp. juv. | a | 3 | 3 | - | 6 |
| Entomobrya corticalis (Nicolet, 1841) | a | 2 | - | - | 2 |
| E. nivalis (Linnaeus, 1758) | a | 2 | - | - | 2 |
| Entomobryides myrmecophilus (Reuter, 1886) | a | 2 | 1 | - | 3 |
| Willowsia buski (Lubbock, 1869) | a | 1 | - | - | 1 |
| Willowsia nigromaculata (Lubbock, 1876) | a | 1 | 4 | 7 | 12 |
| Lepidocyrtus lignorum (Fabricius, 1793) | ep | - | 37 | 47 | 84 |
| Lepidocyrtus sp. 1 | ep | 2 | - | - | 2 |
| Lepidocyrtus violaceus gr juv. | ep | 1 | - | - | 1 |
| Lepidocyrtus lignorum gr juv. | ep | 106 | 3 | 24 | 133 |
| Pseudosinella alba (Packard, 1873) | h | 65 | - | - | 65 |
| Pseudosinella zygophora (Schille, 1908) | h | 156 | 93 | 49 | 298 |
| Entomobyidae juv. | ep | 101 | 54 | 4 | 159 |
| Neelides minutus (Folsom, 1901) | eu | - | 1 | - | 1 |
| Megalothorax minimus (Willem, 1900) | eu | 40 | 33 | 16 | 89 |
| Sphaeridia pumilis (Krausbauer, 1898) | ep | 14 | 38 | 2 | 54 |
| Sminthurides malmgreni (Tullberg, 1876) | a | - | 2 | - | 2 |
| Arrhopalites caecus (Tullberg, 1871) | h | - | 2 | - | 2 |
| A. spinosus (Rusek, 1967) | h | - | 1 | 13 | 14 |
| Arrhopalites sp. juv. | h | 3 | 6 | 3 | 12 |
| Sminthurinus elegans (Fitch, 1863) | ep | 1 | 2 | - | 3 |
| Sminthurinus sp. juv. | ep | 13 | 19 | 9 | 41 |
| Ptenothrix sp. juv. | a | - | - | 7 | 7 |
| Dicyrtoma sp. juv. | a | - | 1 | 2 | 3 |
| Lipotrix lubbocki (Tullberg, 1872) | ep | 8 | 14 | 4 | 26 |
| Allacma fusca (Linnaeus, 1758) | a | 3 | - | - | 3 |
| Caprainea marginata (Schött, 1893) | ep | - | 10 | 10 | 20 |
| Sminthuridae juv. | a | - | - | 2 | 2 |
| Symphypleona juv. | a | 10 | 13 | 1 | 24 |
| Total number of individuals | 2507 | 2086 | 3118 | 7711 | |
| Total number of taxa | 52 | 59 | 63 | 91 |
References
- Gardiner, B.; Blennow, K.; Carnus, J.-M.; Fleischer, P.; Ingemarson, F.; Landmann, G.; Lindner, M.; Marzano, M.; Nicoll, B.; Orazio, C.; et al. Destructive Storms in European Forests: Past and Forthcoming Impacts; Final Report to EC DG Environment; EFI Atlantic: Bordeaux, France, 2010. [Google Scholar]
- Brázdil, R.; Stucki, P.; Szabó, P.; Řezníčková, L.; Dolák, L.; Dobrovolný, P.; Tolasz, R.; Kotyza, O.; Chromá, K.; Suchánková, S. Windstorms and forest disturbances in the Czech Lands: 1801–2015. Agric. For. Meteorol. 2018, 250–251, 47–63. [Google Scholar] [CrossRef]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P.; Ayres, M.P.; Flannigan, M.D.; Hanson, P.J.; Irland, L.C.; Lugo, A.E.; Peterson, C.J.; et al. Climate change and forest disturbances. BioScience 2001, 51, 723–734. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.-J.; Lexer, M.J. Unraveling the drivers of intensifying forest disturbance regimes in Europe. Glob. Chang. Biol. 2011, 17, 2842–2852. [Google Scholar] [CrossRef]
- Rykowski, K. Hurricane in Forests: Calamity or Disturbance in Development? Pisz Forest District 4th July 2002: Case Study; Forest Research Institute: Sękocin Stary, Poland, 2012. [Google Scholar]
- Karpiński, J. Białowieża primeval forest and national park in Białowieża. In Forest Inspectorate; Białowieża: Warsaw, Poland, 1930. [Google Scholar]
- Więcko, E. Management in the Białowieża Forest between the first and the second world wars. Sylwan 1980, 8, 55–65. [Google Scholar]
- Paluch, R.; Bielak, K. Stand conversion by means of natural succession in the Białowieża primeval forest. For. Res. Pap. 2009, 70, 339–354. [Google Scholar] [CrossRef]
- Bird, G.A.; Chatarpaul, L. Effect of whole-tree and conventional forest harvest on soil microarthropods. Can. J. Zool. 1986, 64, 1986–1993. [Google Scholar] [CrossRef]
- Addison, J.A.; Barber, K. Response of Soil Invertebrates to Clear-Cutting and Partial Cutting in a Boreal Mixedwood Forest in Northern Ontario; Great Lakes Forestry Centre: Sault Ste. Marie, ON, Canada, 1997. [Google Scholar]
- Lindo, Z.; Visser, S. Forest floor microarthropod abundance and oribatid mite (Acari: Oribatida) composition following partial and clear-cut harvesting in the mixedwood boreal forest. Can. J. For. Res. 2004, 34, 998–1006. [Google Scholar] [CrossRef]
- Siira-Pietikäinen, A.; Haimi, J. Changes in soil fauna 10 years after forest harvestings: Comparison between clear felling and green-tree retention methods. For. Ecol. Manag. 2009, 258, 332–338. [Google Scholar] [CrossRef]
- Farská, J.; Prejzková, K.; Rusek, J. Management intensity affects traits of soil microarthropod community in montane spruce forest. Appl. Soil Ecol. 2014, 75, 71–79. [Google Scholar] [CrossRef]
- Rousseau, L.; Venier, L.; Hazlett, P.; Fleming, R.; Morris, D.; Handa, I.T. Forest floor mesofauna communities respond to a gradient of biomass removal and soil disturbance in a boreal jack pine (Pinus banksiana) stand of northeastern Ontario (Canada). For. Ecol. Manag. 2018, 407, 155–165. [Google Scholar] [CrossRef]
- Rousseau, L.; Venier, L.; Aubin, I.; Gendreau-Berthiaume, B.; Moretti, M.; Salmon, S.; Handa, I.T. Woody biomass removal in harvested boreal forest leads to a partial functional homogenization of soil mesofaunal communities relative to unharvested forest. Soil Biol. Biochem. 2019, 133, 129–136. [Google Scholar] [CrossRef]
- Berch, S.M.; Battigelli, J.P.; Hope, G.D. Responses of soil mesofauna communities and oribatid mite species to site preparation treatments in high-elevation cutblocks in southern British Columbia. Pedobiologia 2007, 51, 23–32. [Google Scholar] [CrossRef]
- Addison, J. Impact of Retaining Woody Debris and Forest Floor Habitats on Stand. Level of Diversity of Soil Collembola; School of Environment and Sustainability, Royal Road University: Victoria, BC, Canada, 2006. [Google Scholar]
- Bengtsson, J.; Lundkvist, H.; Saetre, P.; Sohlenius, B.; Solbreck, B. Effects of organic matter removal on the soil food web: Forestry practices meet ecological theory. Appl. Soil Ecol. 1998, 9, 137–143. [Google Scholar] [CrossRef]
- Rousseau, L.; Venier, L.; Fleming, R.; Hazlett, P.; Morris, D.; Handa, I.T. Long-term effects of biomass removal on soil mesofaunal communities in northeastern Ontario (Canada) jack pine (Pinus banksiana) stands. For. Ecol. Manag. 2018, 421, 72–83. [Google Scholar] [CrossRef]
- Hopkin, S. Biology of the Springtails: (Insecta: Collembola); OUP: Oxford, UK, 1997. [Google Scholar]
- Rusek, J. Biodiversity of Collembola and their functional role in the ecosystem. Biodivers. Conserv. 1998, 7, 1207–1219. [Google Scholar] [CrossRef]
- Wolters, V. Biodiversity of soil animals and its function. Eur. J. Soil Biol. 2001, 37, 221–227. [Google Scholar] [CrossRef]
- Andre, H.M.; Ducarme, X.; Lebrun, P. Soil biodiversity: Myth, reality or conning? Oikos 2002, 96, 3–24. [Google Scholar] [CrossRef]
- Potapov, A.M.; Goncharov, A.A.; Semenina, E.E.; Korotkevich, A.Y.; Tsurikov, S.M.; Rozanova, O.L.; Anichkin, A.E.; Zuev, A.G.; Samoylova, E.S.; Semenyuk, I.I.; et al. Arthropods in the subsoil: Abundance and vertical distribution as related to soil organic matter, microbial biomass and plant roots. Eur. J. Soil Biol. 2017, 82, 88–97. [Google Scholar] [CrossRef]
- Juceviča, E.; Melecis, V. Long-term dynamics of Collembola in a pine forest ecosystem. Pedobiologia 2002, 46, 365–372. [Google Scholar] [CrossRef]
- Perez, G.; Decaëns, T.; Dujardin, G.; Akpa-Vinceslas, M.; Langlois, E.; Chauvat, M. Response of Collembolan assemblages to plant species successional gradient. Pedobiologia 2013, 56, 169–177. [Google Scholar] [CrossRef]
- Daghighi, E.; Koehler, H.; Kesel, R.; Filser, J. Long-term succession of Collembola communities in relation to climate change and vegetation. Pedobiologia 2017, 64, 25–38. [Google Scholar] [CrossRef]
- Gisin, H. Okologie und Levensgemenischaften der Collembolen im schweizerischen Exkursionsgebiet Basels. Rev. Suisse Zool. 1943, 50, 131–224. [Google Scholar]
- Potapov, A.A.; Semenina, E.E.; Korotkevich, A.Y.; Kuznetsova, N.A.; Tiunov, A.V. Connecting taxonomy and ecology: Trophic niches of Collembolans as related to taxonomic identity and life forms. Soil Biol. Biochem. 2016, 101, 20–31. [Google Scholar] [CrossRef]
- Malmström, A. Life-history traits predict recovery patterns in Collembola species after fire: A 10 year study. Appl. Soil Ecol. 2012, 56, 35–42. [Google Scholar] [CrossRef]
- Pey, B.; Nahmani, J.; Auclerc, A.; Capowiez, Y.; Cluzeau, D.; Cortet, J.; Decaëns, T.; Deharveng, L.; Dubs, F.; Joimel, S.; et al. Current use of and future needs for soil invertebrate functional traits in community ecology. Basic Appl. Ecol. 2014, 15, 194–206. [Google Scholar] [CrossRef]
- Mori, A.S.; Ota, A.T.; Fujii, S.; Seino, T.; Kabeya, D.; Okamoto, T.; Ito, M.T.; Kaneko, N.; Hasegawa, M. Biotic homogenization and differentiation of soil faunal communities in the production forest landscape: Taxonomic and functional perspectives. Oecologia 2015, 177, 533–544. [Google Scholar] [CrossRef]
- da Silva, P.M.; Carvalho, F.; Dirilgen, T.; Stone, D.; Creamer, R.; Bolger, T.; Sousa, J.P. Traits of Collembolan life-form indicate land use types and soil properties across an European transect. Appl. Soil Ecol. 2016, 97, 69–77. [Google Scholar] [CrossRef]
- Materna, J. Collembolan succession on afforested colliery spoil heaps in two contrasting postmining landscapes. In Soil Zoological Problems in Central Europe; Tajovský, K., Pižl, V., Eds.; Institute of Soil Biology, Academy of Sciences of the Czech Republic: České Budéjovice, Czech Republic, 1999; pp. 223–231. [Google Scholar]
- Addison, J.A.; Trofymow, J.A.; Marshall, V.G. Abundance, species diversity, and community structure of Collembola in successional coastal temperate forests on Vancouver Island, Canada. Appl. Soil Ecol. 2003, 24, 233–246. [Google Scholar] [CrossRef]
- Chauvat, M.; Zaitsev, A.S.; Wolters, V. Successional changes of Collembola and soil microbiota during forest rotation. Oecologia 2003, 137, 269–276. [Google Scholar] [CrossRef]
- Chauvat, M.; Trap, J.; Perez, G.; Delporte, P.; Aubert, M. Assemblages of Collembola across a 130-year chronosequence of beech forest. Soil Org. 2011, 83, 405–418. [Google Scholar]
- Dunger, W.; Schulz, H.-J.; Zimdars, B.; Hohberg, K. Changes in Collembolan species composition in Eastern German mine sites over fifty years of primary succession. Pedobiologia 2004, 48, 503–517. [Google Scholar] [CrossRef]
- Dunger, W.; Voigtländer, K. Soil fauna (Lumbricidae, Collembola, Diplopoda and Chilopoda) as indicators of soil eco-subsystem development in post-mining sites of eastern Germany—A review. Soil Org. 2009, 81, 1–51. [Google Scholar]
- Zeppelini, D.; Bellini, B.C.; Creão-Duarte, A.J.; Hernández, M.I.M. Collembola as bioindicators of restoration in mined sand dunes of Northeastern Brazil. Biodivers. Conserv. 2009, 18, 1161–1170. [Google Scholar] [CrossRef]
- Michalczuk, C. Forest habitat and tree stands of Białowieża National Park. Phytocenosis Suppl. Cartogr. Geobot. 2001, 13, 1–22. [Google Scholar]
- Babenko, A.; Chernova, N.; Potapov, M.; Stebaeva, S. Collembola of Russia and Adjacent Countries: Family Hypogastruridae; Nauka: Moscow, Russia, 1994. [Google Scholar]
- Fjellberg, A. The Collembola of Fennoscandia and Denmark Part 1: Poduromorpha; Fauna Entomologica Scandinavica; Brill: Leiden, The Netherlands, 1998. [Google Scholar]
- Fjellberg, A. The Collembola of Fennoscandia and Denmark Part 2: Entomobryomorpha and Symphypleona; Fauna Entomologica Scandinavica; Brill: Leiden, The Netherlands, 2007. [Google Scholar]
- Pomorski, R.J. Onychiurinae of Poland (Collembola: Onychiuridae). Genus 1998, 9, 1–201. [Google Scholar]
- Bretfeld, G. Symphypleona. In Synopses on Palearctic Collembola; Dunger, W., Ed.; Staatliches Museum für Naturkunde Görlitz: Görlitz, Germany, 1999; pp. 1–318. [Google Scholar]
- Potapov, M. Isotomidae. In Synopses on Palaearctic Collembola; Dunger, W., Ed.; Staatliches Museum fr Naturkunde Görlitz: Görlitz: Germany, 2001; pp. 1–603. [Google Scholar]
- Thibaud, J.M.; Schulz, H.J.; Assalino, M.M.D.G. Hypogastruridae. In Synopses on Palaearctic Collembola; Dunger, W., Ed.; Staatliches Museum für Naturkunde Görlitz: Görlitz, Germany, 2004; pp. 1–287. [Google Scholar]
- Dunger, W.; Schlitt, B. Synopses on Palearctic Collembola: Tullbergiidae; Staatliches Museum für Naturkunde Görlitz: Görlitz, Germany, 2011. [Google Scholar]
- Petersen, H. General aspects of Collembolan ecology at the turn of the millennium. Pedobiologia 2002, 46, 246–260. [Google Scholar] [CrossRef]
- Chao, A.; Jost, L. Coverage-based rarefaction and extrapolation: Standardizing samples by completeness rather than size. Ecology 2012, 93, 2533–2547. [Google Scholar] [CrossRef]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef]
- Hsieh, T.C.; Ma, K.H.; Chao, A.; McInerny, G. iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 8 January 2019).
- Oksanen, J.; Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.; O’Hara, R.; Simpson, G.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package. Available online: http://CRAN.Rproject. org/package=vegan (accessed on 8 January 2019).
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Anderson, M.J.; Walsh, D.C.I. PERMANOVA, ANOSIM, and the Mantel test in the face of heterogeneous dispersions: What null hypothesis are you testing? Ecol. Monogr. 2013, 83, 557–574. [Google Scholar] [CrossRef]
- Dufrene, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P.; Moretti, M. Improving indicator species analysis by combining groups of sites. Oikos 2010, 119, 1674–1684. [Google Scholar] [CrossRef]
- Beasley, T.M.; Schumacker, R.E. Multiple regression approach to analyzing contingency tables: Post Hoc and planned comparison procedures. J. Exp. Educ. 1995, 64, 79–93. [Google Scholar] [CrossRef]
- Covington, W.W. Changes in forest floor organic matter and nutrient content following clear cutting in Northern Hardwoods. Ecology 1981, 62, 41–48. [Google Scholar] [CrossRef]
- Seastedt, T.R.; Crossley, D.A. Microarthropod response following cable logging and clear-cutting in the southern appalachians. Ecology 1981, 62, 126–135. [Google Scholar] [CrossRef]
- Blair, J.M.; Crossley, D.A. Litter decomposition, nitrogen dynamics and litter microarthropods in a southern Appalachian hardwood forest 8 years following clearcutting. J. Appl. Ecol. 1988, 25, 683–698. [Google Scholar] [CrossRef]
- Sterzyńska, M. Collembola in the process of secondary succession of the pine forests of Puszcza Białowieska. Fragm. Faun. 1995, 38, 353–364. [Google Scholar] [CrossRef]
- Malmström, A.; Persson, T.; Ahlström, K.; Gongalsky, K.B.; Bengtsson, J. Dynamics of soil meso- and macrofauna during a 5-year period after clear-cut burning in a boreal forest. Appl. Soil Ecol. 2009, 43, 61–74. [Google Scholar] [CrossRef]
- Siira-Pietikäinen, A.; Haimi, J.; Siitonen, J. Short-term responses of soil macroarthropod community to clear felling and alternative forest regeneration methods. For. Ecol. Manag. 2003, 172, 339–353. [Google Scholar] [CrossRef]
- Kopeszki, H. Collembolan fauna, in Vienna beech wood in relation to litter accumulation and depletion. Pol. Pismo Entomol. 1995, 64, 357–362. [Google Scholar]
- Setälä, H.; Haimi, J.; Siira-Pietikäinen, A. Sensitivity of soil processes in northern forest soils: Are management practices a threat? For. Ecol. Manag. 2000, 133, 5–11. [Google Scholar] [CrossRef]
- Huhta, V.; Karppinen, E.; Nurminen, M.; Valpas, A. Effect of silvicultural practices upon Arthropos, Annelid and Nematode populations in coniferous forest soil. Ann. Zool. Fenn. 1967, 4, 87–145. [Google Scholar]
- Huhta, V.; Nurminen, M.; Valpas, A. Further notes on the effect of silvicultural practices upon the fauna of coniferous forest soil. Ann. Zool. Fenn. 1969, 6, 327–334. [Google Scholar]
- Huhta, V. Effects of clear-cutting on numbers, biomass and community respiration of soil invertebrates. Ann. Zool. Fenn. 1976, 13, 63–80. [Google Scholar]
- Bird, S.B.; Coulson, R.N.; Fisher, R.F. Changes in soil and litter arthropod abundance following tree harvesting and site preparation in a loblolly pine (Pinus taeda L.) plantation. For. Ecol. Manag. 2004, 202, 195–208. [Google Scholar] [CrossRef]
- Cassagne, N.; Gers, C.; Gauquelin, T. Relationships between Collembola, soil chemistry and humus types in forest stands (France). Biol. Fertil. Soils 2003, 37, 355–361. [Google Scholar] [CrossRef]
- Kováč, L.u.; Kostúrová, N.; Miklisová, D. Comparison of collembolan assemblages (Hexapoda, Collembola) of thermophilous oak woods and Pinus nigra plantations in the Slovak Karst (Slovakia). Pedobiologia 2005, 49, 29–40. [Google Scholar] [CrossRef]
- Salmon, S.; Artuso, N.; Frizzera, L.; Zampedri, R. Relationships between soil fauna communities and humus forms: Response to forest dynamics and solar radiation. Soil Biol. Biochem. 2008, 40, 1707–1715. [Google Scholar] [CrossRef]
- Sławska, M.; Bruckner, A.; Sławski, M. Edaphic Collembola assemblages of European temperate primeval forests gradually change along a forest-type gradient. Eur. J. Soil Biol. 2017, 80, 92–101. [Google Scholar] [CrossRef]
- Zanella, A.; Ponge, J.-F.; Briones, M.J.I. Humusica 1, article 8: Terrestrial humus systems and forms—Biological activity and soil aggregates, space-time dynamics. Appl. Soil Ecol. 2018, 122, 103–137. [Google Scholar] [CrossRef]
- Hasegawa, M.; Fukuyama, K.; Makino, S.I.; Okochi, I.; Goto, H.; Mizoguchi, T.; Sakata, T.; Tanaka, H. Collembolan community dynamics during deciduous forests regeneration in Japan. Pedobiologia 2006, 50, 117–126. [Google Scholar] [CrossRef]
- Čuchta, P.; Miklisová, D.; Kováč, Ľ. The succession of soil Collembola communities in spruce forests of the High Tatra Mountains five years after a windthrow and clear–cut logging. For. Ecol. Manag. 2019, 433, 504–513. [Google Scholar] [CrossRef]
- Korboulewsky, N.; Perez, G.; Chauvat, M. How tree diversity affects soil fauna diversity: A review. Soil Biol. Biochem. 2016, 94, 94–106. [Google Scholar] [CrossRef]
- Russell, D.J.; Gergócs, V. Forest-management types similarly influence soil collembolan communities throughout regions in Germany—A data bank analysis. For. Ecol. Manag. 2019, 434, 49–62. [Google Scholar] [CrossRef]
- Skarżyński, D.; Piwnik, A.; Krzysztofiak, A. Saproxylic springtails (Collembola) of the Wigry National Park. For. Res. Pap. 2016, 77, 186–203. [Google Scholar] [CrossRef]
- Sławski, M. Analysis of forest structure in relation to age—Scots pine case study. Sylwan 2011, 155, 10–20. [Google Scholar]
- Marshall, V.G. Impacts of forest harvesting on biological processes in northern forest soils. For. Ecol. Manag. 2000, 133, 43–60. [Google Scholar] [CrossRef]
- Auclerc, A.; Ponge, J.F.; Barot, S.; Dubs, F. Experimental assessment of habitat preference and dispersal ability of soil springtails. Soil Biol. Biochem. 2009, 41, 1596–1604. [Google Scholar] [CrossRef]
- Picchio, R.; Venanzi, R.; Tavankar, F.; Luchenti, I.; Iranparast Bodaghi, A.; Latterini, F.; Nikooy, M.; Di Marzio, N.; Naghdi, R. Changes in soil parameters of forests after windstorms and timber extraction. Eur. J. For. Res. 2019, 138, 875–888. [Google Scholar] [CrossRef]
- Gomez, A.; Powers, R.F.; Singer, M.J.; Horwath, W.R. Soil compaction effects on growth of young ponderosa pine following litter removal in California’s Sierra Nevada. Soil Sci. Soc. Am. J. 2002, 66, 1334–1343. [Google Scholar] [CrossRef]
- Cambi, M.; Hoshika, Y.; Mariotti, B.; Paoletti, E.; Picchio, R.; Venanzi, R.; Marchi, E. Compaction by a forest machine affects soil quality and Quercus robur L. seedling performance in an experimental field. For. Ecol. Manag. 2017, 384, 406–414. [Google Scholar] [CrossRef]
- Geissen, V.; Kim, R.Y.; Schöning, A.; Schütte, S.; Brümmer, G.W. Effects of strip wise tillage in combination with liming on chemical and physical properties of acidic spruce forest soils after clear cutting. For. Ecol. Manag. 2003, 180, 75–83. [Google Scholar] [CrossRef]
- Battigelli, J.P.; Spence, J.R.; Langor, D.W.; Berch, S.M. Short-term impact of forest soil compaction and organic matter removal on soil mesofauna density and oribatid mite diversity. Can. J. For. Res. 2004, 34, 1136–1149. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Indicator Species | Forest Type | Specificity | Fidelity | IndVal Test Statistics | Significance |
|---|---|---|---|---|---|
| Micraphorura absoloni | Res | 0.74 | 1 | 0.861 | 0.014 |
| Mesaphorura yosii | Res | 0.85 | 0.83 | 0.843 | 0.014 |
| Arrhopalites spinosus | Res | 0.93 | 0.67 | 0.787 | 0.011 |
| Pogonognatellus flavescens | Mat + Res | 0.97 | 0.92 | 0.945 | 0.023 |
| Lepidocyrtus lignorum | Mat + Res | 1 | 0.83 | 0.913 | 0.004 |
| Protaphorura subarmata | Mat + Res | 1 | 0.67 | 0.816 | 0.036 |
| Protaphorura pannonica | Spn + Res | 0.8651 | 1 | 0.93 | 0.019 |
| Life Form | Spn | Mat | Res | Total N |
|---|---|---|---|---|
| Atmobiotic | 1% (−2.07)NS | 4% (7.66)*** | 1% (−4.96)*** | 143 |
| Epedaphic | 11% (−9.83)*** | 24% (−9.62)*** | 17% (0.67)NS | 1299 |
| Hemiedaphic | 60% (17.73)*** | 41% (−4.76)*** | 36% (−12.61)*** | 3476 |
| Euedaphic | 28% (−10.12)*** | 32% (−4.72)*** | 45% (13.93)*** | 2793 |
| Total N | 2507 | 2086 | 3118 | 7711 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sławski, M.; Sławska, M. Seven Decades of Spontaneous Forest Regeneration after Large-Scale Clear-Cutting in Białowieża Forest do not Ensure the Complete Recovery of Collembolan Assemblages. Forests 2019, 10, 948. https://doi.org/10.3390/f10110948
Sławski M, Sławska M. Seven Decades of Spontaneous Forest Regeneration after Large-Scale Clear-Cutting in Białowieża Forest do not Ensure the Complete Recovery of Collembolan Assemblages. Forests. 2019; 10(11):948. https://doi.org/10.3390/f10110948
Chicago/Turabian StyleSławski, Marek, and Małgorzata Sławska. 2019. "Seven Decades of Spontaneous Forest Regeneration after Large-Scale Clear-Cutting in Białowieża Forest do not Ensure the Complete Recovery of Collembolan Assemblages" Forests 10, no. 11: 948. https://doi.org/10.3390/f10110948
APA StyleSławski, M., & Sławska, M. (2019). Seven Decades of Spontaneous Forest Regeneration after Large-Scale Clear-Cutting in Białowieża Forest do not Ensure the Complete Recovery of Collembolan Assemblages. Forests, 10(11), 948. https://doi.org/10.3390/f10110948