Understanding the Critical Impact Path on Vegetation Growth under Climate Extremes and Human Influence


Abstract
1. Introduction
2. Materials and Methods
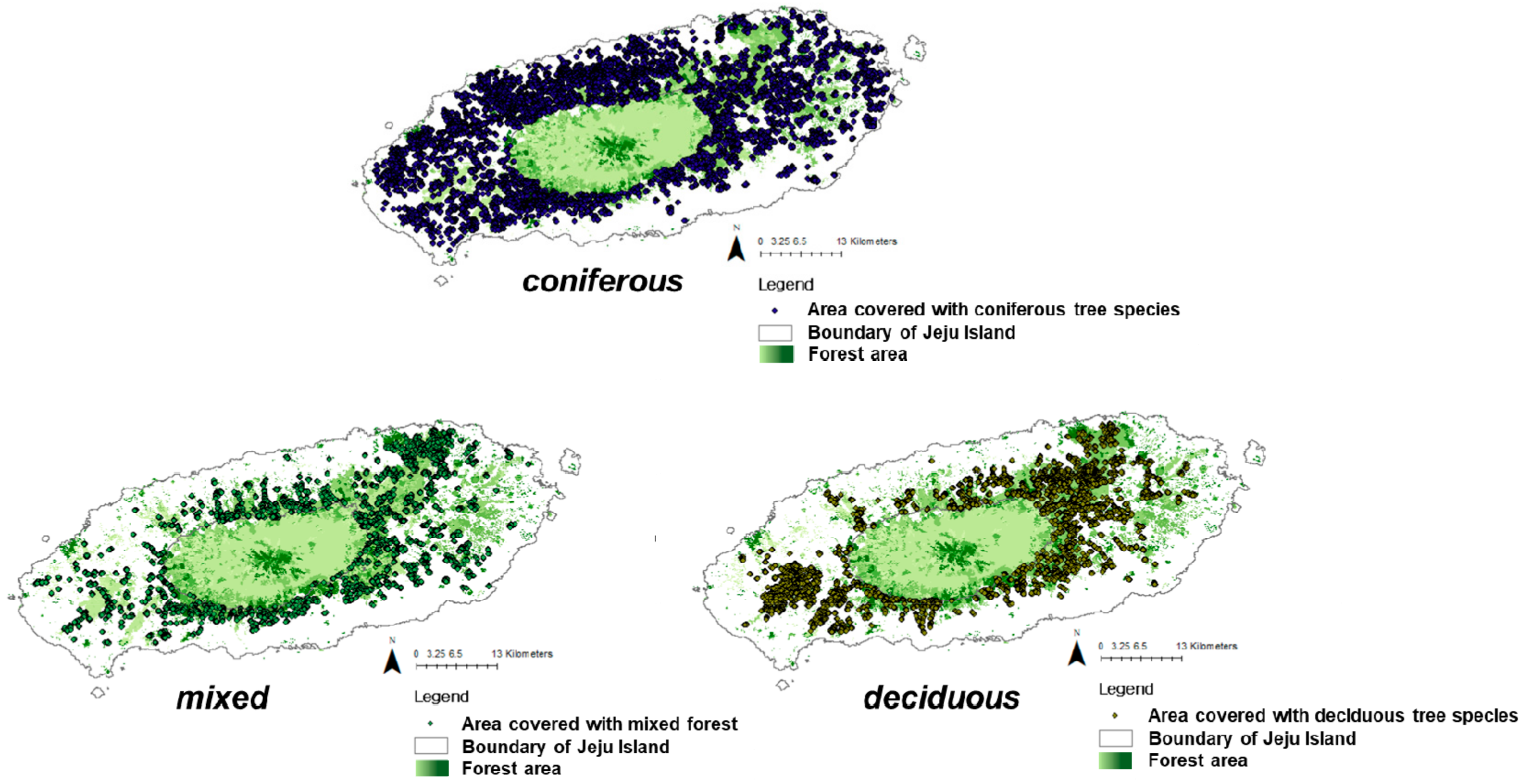
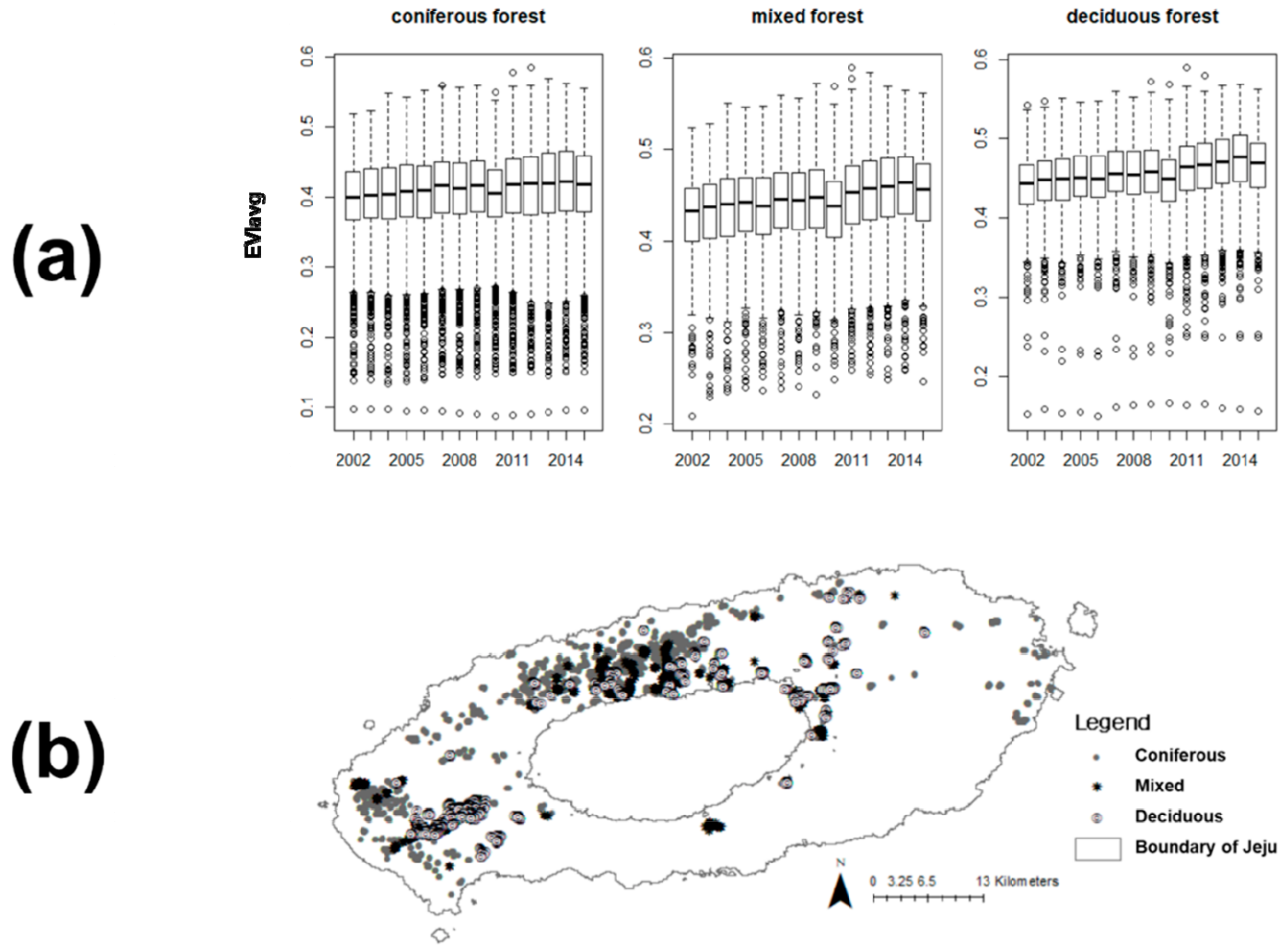
2.1. Study Site and Vegetation for Case Study
2.2. Data Processing for Annual Average EVI (EVIavg)
2.3. Data Processing for Land Conversion
2.4. Climate Extreme Indices
2.5. Construction of Bayesian Network
2.6. Validation of Developed Bayesian Networks
3. Results
3.1. Temporal Changes in EVI
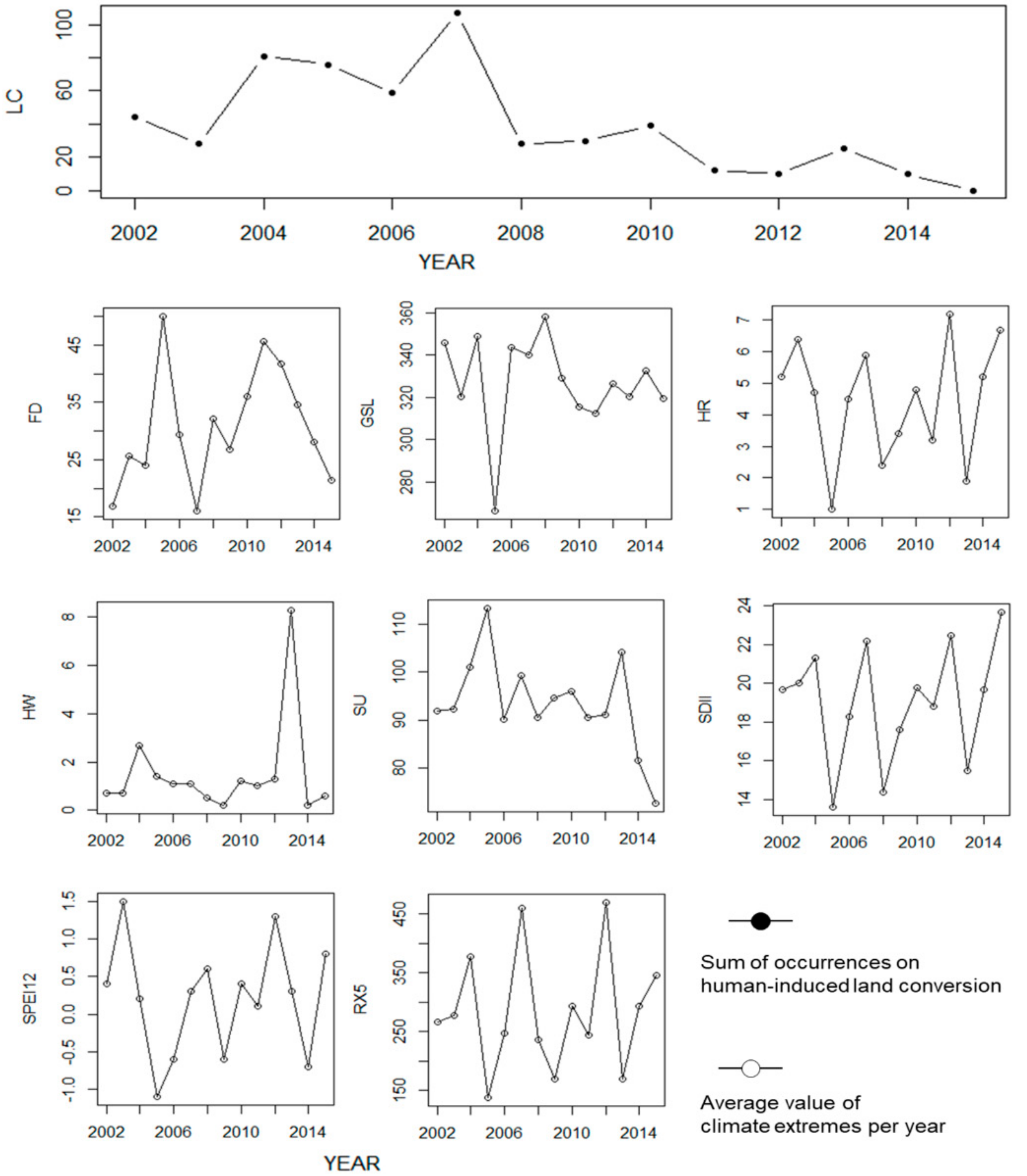
3.2. Temporal Changes in Human-Induced Land Conversion and Climate Extremes
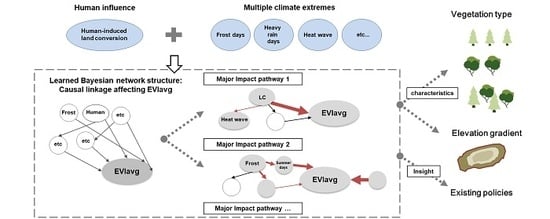
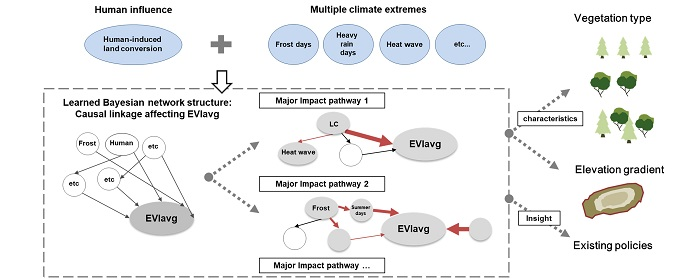
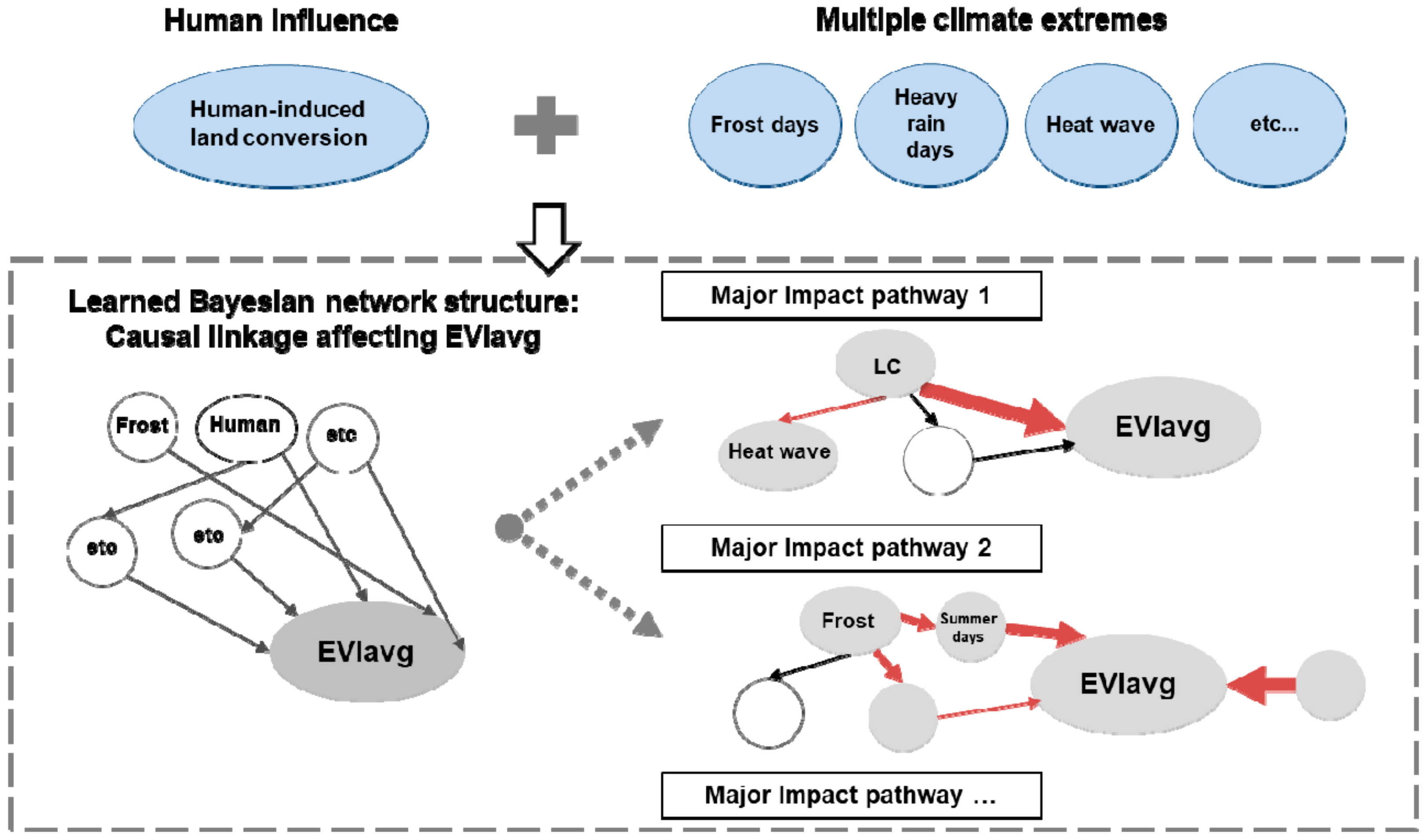
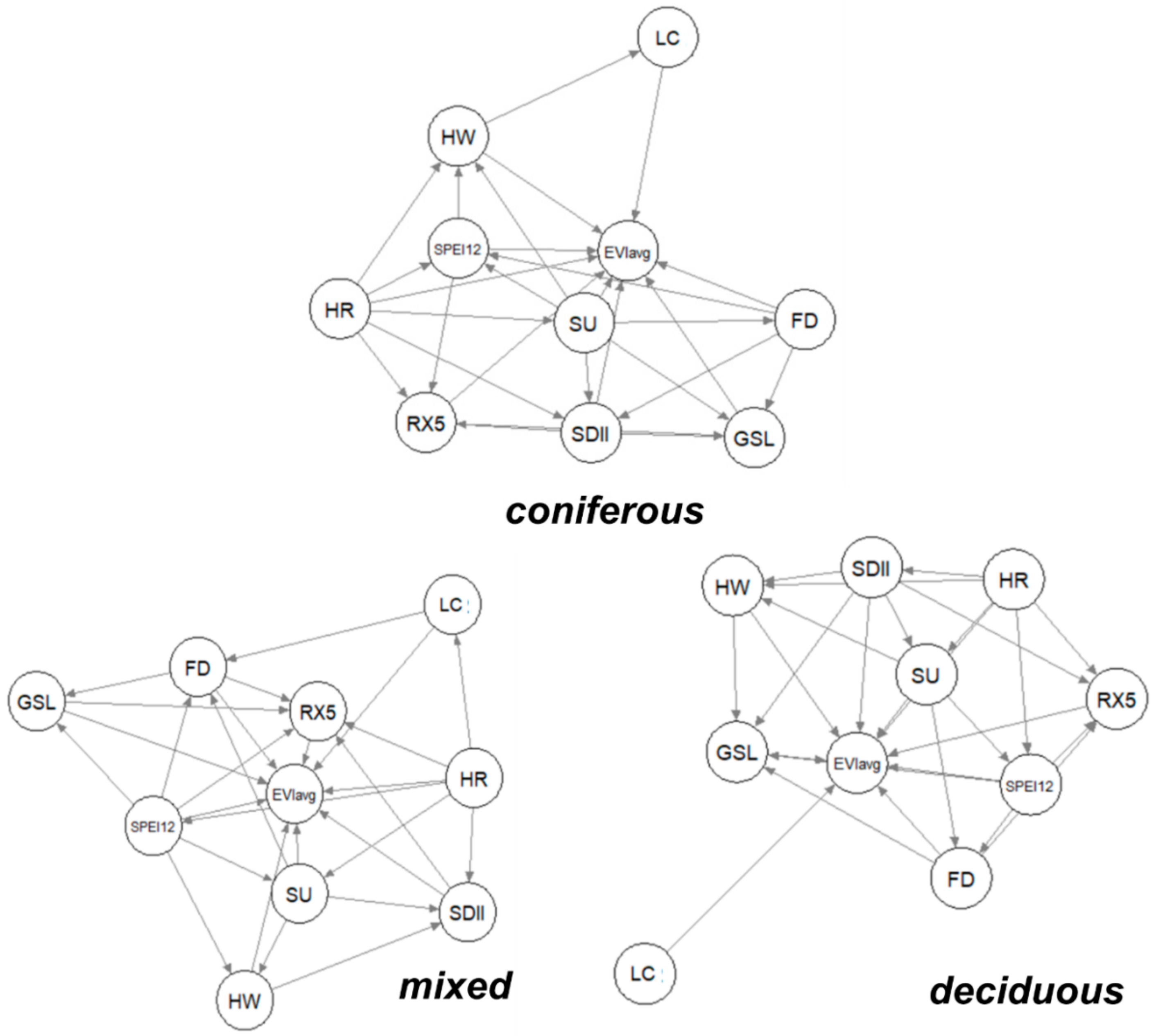
3.3. Causal Network among EVIavg, Climate Extremes, and Human-Induced Land Cover Change
3.3.1. Climate and Human Impacts per Vegetation Type
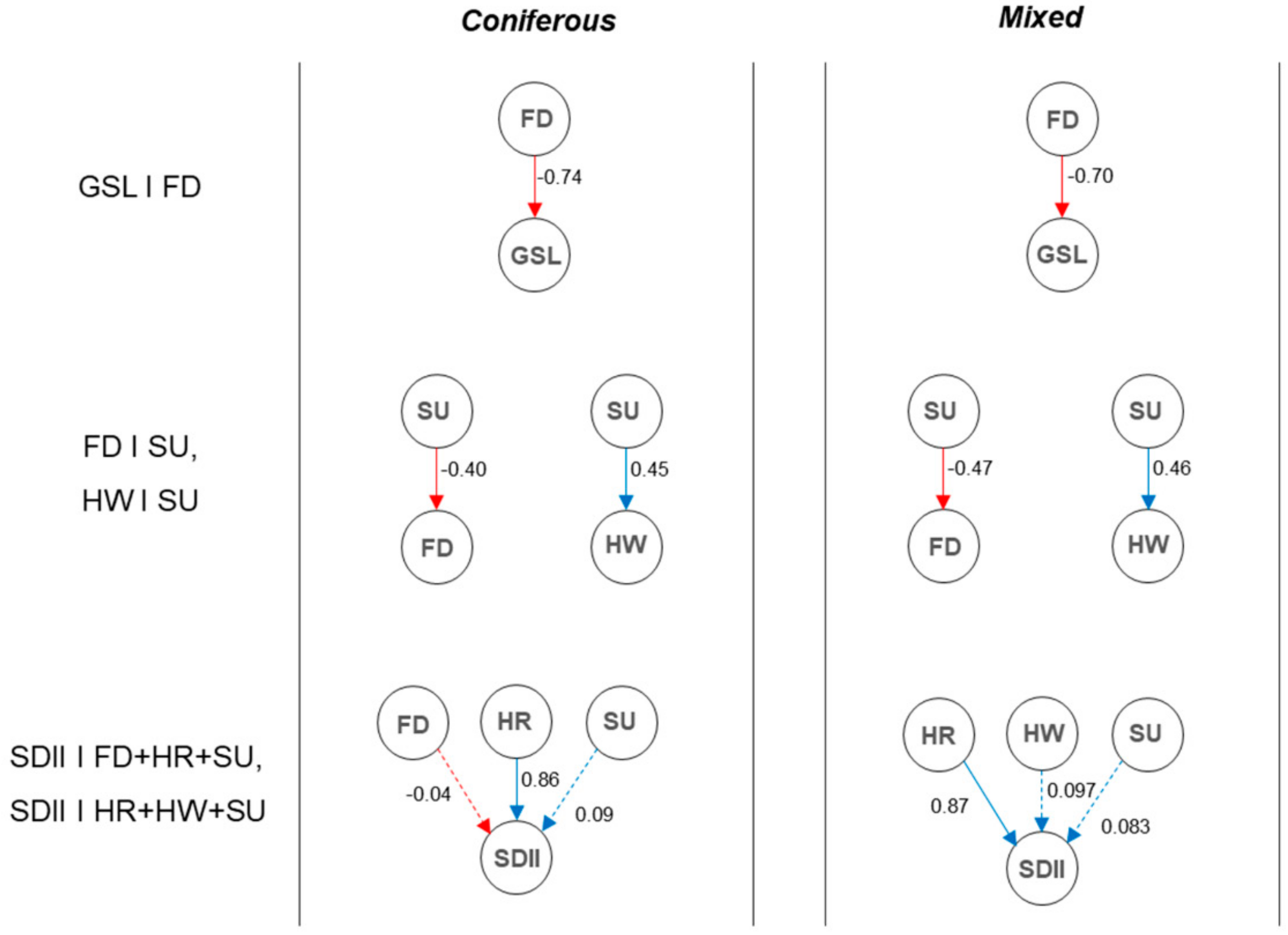
3.3.2. Critical Impact Path Regarding Human-Induced LC
3.3.3. Critical Impact Path Regarding Climate Extremes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Yu, D.; Shao, H.; Shi, P.; Zhu, W.; Pan, Y. Agricultural and Forest Meteorology How does the conversion of land cover to urban use affect net primary productivity? A case study in Shenzhen city, China. Agric. For. Meteorol. 2009, 149, 2054–2060. [Google Scholar] [CrossRef]
- Pimm, S.L.; Raven, P. Biodiversity: Extinction by numbers. Nature 2000, 403, 843. [Google Scholar] [CrossRef] [PubMed]
- Leemans, R.; De Groot, R.S. Millennium Ecosystem Assessment: Ecosystems and Human Well-Being: A Framework for Assessment; Island Press: Washington, DC, USA, 2003. [Google Scholar]
- DeFries, R.S.; Field, C.B.; Fung, I.; Collatz, G.J.; Bounoua, L. Combining satellite data and biogeochemical models to estimate global effects of human-induced land cover change on carbon emissions and primary productivity. Glob. Biogeochem. Cycles 1999, 13, 803–815. [Google Scholar] [CrossRef]
- Chapin, F.S.; Zavaleta, E.S.; Eviner, V.T.; Naylor, R.L.; Vitousek, P.M.; Reynolds, H.L.; Hooper, D.U.; Lavorel, S.; Sala, O.E.; Hobbie, S.E.; et al. Consequences of changing biodiversity. Nature 2000, 405, 234–242. [Google Scholar] [CrossRef]
- Breshears, D.D.; Cobb, N.S.; Rich, P.M.; Price, K.P.; Allen, C.D.; Balice, R.G.; Romme, W.H.; Kastens, J.H.; Floyd, M.L.; Belnap, J. Regional vegetation die-off in response to global-change-type drought. Proc. Natl. Acad. Sci. USA 2005, 102, 15144–15148. [Google Scholar] [CrossRef]
- Nezlin, N.P.; Kostianoy, A.G.; Li, B.-L. Inter-annual variability and interaction of remote-sensed vegetation index and atmospheric precipitation in the Aral Sea region. J. Arid Environ. 2005, 62, 677–700. [Google Scholar] [CrossRef]
- Wang, J.; Wang, K.; Zhang, M.; Zhang, C. Impacts of climate change and human activities on vegetation cover in hilly Southern China. Ecol. Eng. 2015, 81, 451–461. [Google Scholar] [CrossRef]
- Jiang, L.; Jiapaer, G.; Bao, A.; Guo, H.; Ndayisaba, F. Vegetation dynamics and responses to climate change and human activities in Central Asia. Sci. Total Environ. 2017, 599, 967–980. [Google Scholar] [CrossRef]
- CBD (Convention of Biological Diversity). Decision Adopted by the Conference of the Parties to the Convention on BiologicaL Diversity: CBD/COP/DEC/XIII/3. Available online: https://www.cbd.int/doc/decisions/cop-13/cop-13-dec-03-en.pdf (accessed on 20 October 2018).
- Secretariat of the Convention on Biological Diversity. GBO 4 Global Biodiversity Outlook. A Mid-term Assessment of Progress towards the Implementation of the Strategic Plan for Biodiversity 2011–2020. Available online: https://www.cbd.int/gbo/gbo4/publication/gbo4-en.pdf (accessed on 10 October 2018).
- Settele, J.; Scholes, R.; Betts, R.A. Terrestrial and inland water systems. In Climate Change 2014 Impacts, Adaptation and Vulnerability: Part A: Global and Sectoral Aspects; Field, C.B., Barros, V.R., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2015; pp. 271–360. [Google Scholar]
- Hoegh-Guldberg, O.; Jacob, D.; Taylor, M. Impacts of 1.5 °C of Global Warming on Natural and Human Systems. In Global Warming of 1.5 °C. An IPCC Special Report on the Impacts of Global Warming of 1.5 °C above Pre-industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty, 1st ed.; Masson-Delmotte, V., Zhai, P., Eds.; Springer: Berlin, Germany, 2018; pp. 175–312. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species Version 2018-1. Available online: https://www.iucnredlist.org (accessed on 1 September 2018).
- Di Vittorio, A.V.; Mao, J.; Shi, X.; Chini, L.; Hurtt, G.; Collins, W.D. Quantifying the Effects of Historical Land Cover Conversion Uncertainty on Global Carbon and Climate Estimates. Geophys. Res. Lett. 2018, 45, 974–982. [Google Scholar] [CrossRef]
- Pan, N.; Feng, X.; Fu, B.; Wang, S.; Ji, F.; Pan, S. Increasing global vegetation browning hidden in overall vegetation greening: Insights from time-varying trends. Remote Sens. Environ. 2018, 214, 59–72. [Google Scholar] [CrossRef]
- Liu, R.; Xiao, L.; Liu, Z.; Dai, J. Quantifying the relative impacts of climate and human activities on vegetation changes at the regional scale. Ecol. Indic. 2018, 93, 91–99. [Google Scholar] [CrossRef]
- Qi, X.; Jia, J.; Liu, H.; Lin, Z. Relative importance of climate change and human activities for vegetation changes on China’s silk road economic belt over multiple timescales. Catena 2019, 180, 224–237. [Google Scholar] [CrossRef]
- Zheng, K.; Wei, J.Z.; Pei, J.Y.; Cheng, H.; Zhang, X.L.; Huang, F.Q.; Li, F.M.; Ye, J.S. Impacts of climate change and human activities on grassland vegetation variation in the Chinese Loess Plateau. Sci. Total Environ. 2019, 660, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Conroy, M.J.; Peterson, J.T. Decision Making in Natural Resource Management: A Structured, Adaptive Approach; Wiley-Blackwell: West Succex, UK, 2013; ISBN 978-047-067-175-7. [Google Scholar]
- Darling-Hammond, L. The implications of testing policy for quality and equality. Phi Delta Kappan 1991, 73, 220–225. [Google Scholar]
- Clemen, R.T.; Reilly, T. Making Hard Decisions with Decision Tools Suite, 2nd ed.; Duxbury Thomson Learning: Pacific Grove, CA, USA, 2001; ISBN 053-436-597-3. [Google Scholar]
- Augspurger, C.K. Reconstructing patterns of temperature, phenology, and frost damage over 124 years: Spring damage risk is increasing. Ecology 2013, 94, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Park, C.; Koo, K.A.; Lee, M.K.; Lee, D.K. Evaluating multiple bioclimatic risks using Bayesian belief network to support urban tree management under climate change. Urban For. Urban Green. 2019, 43. [Google Scholar] [CrossRef]
- Easterling, D.R.; Meehl, G.A.; Parmesan, C.; Changnon, S.A.; Karl, T.R.; Mearns, L.O. Climate extremes: Observations, modeling, and impacts. Science 2000, 289, 2068–2074. [Google Scholar] [CrossRef]
- Gonzalez-Roglich, M.; Zvoleff, A.; Noon, M.; Liniger, H.; Fleiner, R.; Harari, N.; Garcia, C. Synergizing global tools to monitor progress towards land degradation neutrality: Trends. Earth and the World Overview of Conservation Approaches and Technologies sustainable land management database. Environ. Sci. Policy 2019, 93, 34–42. [Google Scholar] [CrossRef]
- De Long, J.R.; Kardol, P.; Sundqvist, M.K.; Veen, G.F.; Wardle, D.A. Plant growth response to direct and indirect temperature effects varies by vegetation type and elevation in a subarctic tundra. Oikos 2015, 124, 772–783. [Google Scholar] [CrossRef]
- He, D.; Yi, G.; Zhang, T.; Miao, J.; Li, J.; Bie, X. Temporal and spatial characteristics of EVI and its response to climatic factors in recent 16 years based on grey relational analysis in inner Mongolia Autonomous Region, China. Remote Sens. 2018, 10, 961. [Google Scholar] [CrossRef]
- Mccann, R.K.; Marcot, B.G.; Ellis, R. Bayesian belief networks: Applications in ecology and natural resource management. Can. J. For. Res. 2006, 36, 3053–3063. [Google Scholar] [CrossRef]
- Barton, D.N.; Kuikka, S.; Varis, O.; Uusitalo, L.; Henriksen, J.; Borsuk, M.; De Hera, A.; Farmani, R.; Johnson, S.; Linnell, J.D. Bayesian Networks in Environmental and Resource Management. Integr. Environ. Assess Manag. 2012, 8, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, P.A.; Fernández, A.; Fernández, R.; Rumí, R.; Salmerón, A. Bayesian networks in environmental modelling. Environ. Model. Softw. 2011, 26, 1376–1388. [Google Scholar] [CrossRef]
- Woo-Seok, K.; Watts, P. The Plant Geography of Korea: With an Emphasis on the Alpine Zones, 1st ed.; Kluwer Academic Publishers: Dordrecht, Netherlands, 1993; ISBN 940-104-708-1. [Google Scholar]
- Schaaf, T.; Rodrigues, D.C. Managing MIDAs: Harmonising the Management of Multi-internationnaly Designated Areas: Ramsar Sites, World Heritage Sites, Biosphere Reserves and UNESCO Global Geoparks; IUCN International Union for Conservation of Nature and Natural Resources: Gland, Switzerland, 2016; ISBN 978-2-8317-1793-7. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Release 10.6.; ESRI, Inc.: Redlands, CA, USA, 2018. [Google Scholar]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Wu, S.; Liang, Z.; Li, S. Relationships between urban development level and urban vegetation states: A global perspective. Urban For. Urban Green. 2019, 38, 215–222. [Google Scholar] [CrossRef]
- Waring, R.H.; Coops, N.C.; Fan, W.; Nightingale, J.M. MODIS enhanced vegetation index predicts tree species richness across forested ecoregions in the contiguous USA. Remote Sens. Environ. 2006, 103, 218–226. [Google Scholar] [CrossRef]
- Zhang, Y.; Song, C.; Band, L.E.; Sun, G.; Li, J. Reanalysis of global terrestrial vegetation trends from MODIS products: Browning or greening? Remote Sens. Environ. 2017, 191, 145–155. [Google Scholar] [CrossRef]
- Chen, J.; Jönsson, P.; Tamura, M.; Gu, Z.; Matsushita, B.; Eklundh, L. A simple method for reconstructing a high-quality NDVI time-series data set based on the Savitzky–Golay filter. Remote Sens. Environ. 2004, 91, 332–344. [Google Scholar] [CrossRef]
- Peng, D.; Wu, C.; Zhang, B.; Huete, A.; Zhang, X.; Sun, R.; Lei, L.; Huang, W.; Liu, L.; Liu, X.; et al. The influences of drought and land-cover conversion on inter-annual variation of NPP in the Three-North Shelterbelt Program zone of China based on MODIS data. PLoS ONE 2016, 11, e0158173. [Google Scholar] [CrossRef]
- RStudio, T. RStudio: Integrated Development Environment for R; RStudio, Inc.: Boston, MA, USA, 2016. [Google Scholar]
- Zhang, X.; Alexander, L.; Hegerl, G.C.; Jones, P.; Tank, A.K.; Peterson, T.C.; Trewin, B.; Zwiers, F.W. Indices for monitoring changes in extremes based on daily temperature and precipitation data. Wiley Interdiscip. Rev. Clim. Chang. 2011, 2, 851–870. [Google Scholar] [CrossRef]
- Beguería, S.; Vicente-Serrano, S.M.; Reig, F.; Latorre, B. Standardized precipitation evapotranspiration index (SPEI) revisited: Parameter fitting, evapotranspiration models, tools, datasets and drought monitoring. Int. J. Climatol. 2014, 34, 3001–3023. [Google Scholar] [CrossRef]
- Scutari, M. Learning Bayesian Networks with the bnlearn R Package. J. Stat. Softw. 2010, 35, 1–22. [Google Scholar] [CrossRef]
- Hair, J.F.; Anderson, R.E.; Tatham, R.L. Multivariate Data Analysis with Readings; Petroleum Publishing: Tulsa, OK, USA, 1984. [Google Scholar]
- Aliferis, C.F.; Tsamardinos, I.; Statnikov, A.R.; Brown, L.E. Causal Explorer: A Causal Probabilistic Network Learning Toolkit for Biomedical Discovery. In Proceedings of the METMBS, Las Vegas, NV, USA, 3 June 2003; Volume 3, pp. 371–376. [Google Scholar]
- Tsamardinos, I.; Brown, L.E.; Aliferis, C.F. The max-min hill-climbing Bayesian network structure learning algorithm. Mach. Learn. 2006, 65, 31–78. [Google Scholar] [CrossRef]
- Reyer, C.; Bathgate, S.; Blennow, K.; Borges, J.G.; Bugmann, H.; Delzon, S.; Faias, S.P.; Garcia-Gonzalo, J.; Gardiner, B.; Gonzalez-Olabarria, J.R.; et al. Are forest disturbances amplifying or canceling out climate change-induced productivity changes in European forests? Environ. Res. Lett. 2017, 12, 034027. [Google Scholar] [CrossRef]
- Danneyrolles, V.; Dupuis, S.; Fortin, G.; Leroyer, M.; De Römer, A.; Terrail, R.; Vellend, M.; Boucher, Y.; Laflamme, J.; Bergeron, Y. Stronger influence of anthropogenic disturbance than climate change on century-scale compositional changes in northern forests. Nat. Commun. 2019, 10, 1265. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.-Z.; Wang, C.-J.; Qu, H.; Liu, R.; Zhang, Z.-X. Vulnerability of forest vegetation to anthropogenic climate change in China. Sci. Total Environ. 2018, 621, 1633–1641. [Google Scholar] [CrossRef]
- Schelhaas, M.; Nabuurs, G.; Schuck, A. Natural disturbances in the European forests in the 19th and 20th centuries. Glob. Chang. Biol. 2003, 9, 1620–1633. [Google Scholar] [CrossRef]
- Matías, L.; Jump, A.S. Interactions between growth, demography and biotic interactions in determining species range limits in a warming world: The case of Pinus sylvestris. For. Ecol. Manag. 2012, 282, 10–22. [Google Scholar] [CrossRef]
- Milad, M.; Schaich, H.; Konold, W. How is adaptation to climate change reflected in current practice of forest management and conservation? A case study from Germany. Biodivers. Conserv. 2013, 22, 1181–1202. [Google Scholar] [CrossRef]
- Thuiller, W.; Lavorel, S.; Araújo, M.B.; Sykes, M.T.; Prentice, I.C. Climate change threats to plant diversity in Europe. Proc. Natl. Acad. Sci. USA 2005, 102, 8245–8250. [Google Scholar] [CrossRef]
- Nolan, C.; Overpeck, J.T.; Allen, J.R.M.; Anderson, P.M.; Betancourt, J.L.; Binney, H.A.; Brewer, S.; Bush, M.B.; Chase, B.M.; Cheddadi, R. Past and future global transformation of terrestrial ecosystems under climate change. Science 2018, 361, 920–923. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Xia, J.; Wang, Y.; Ahlström, A.; Chen, J.; Cook, R.B.; Cui, E.; Fang, Y.; Fisher, J.B.; Huntzinger, D.N. Enhanced peak growth of global vegetation and its key mechanisms. Nat. Ecol. Evol. 2018, 2, 1897. [Google Scholar] [CrossRef] [PubMed]
- Snyder, R.L.; Melo Abreu, J.D. Frost Protection: Fundamentals, Practice and Economics; FAO: Rome, Italy, 2005; ISBN 925-105-329-4. [Google Scholar]
- Liu, Q.; Piao, S.; Janssens, I.A.; Fu, Y.; Peng, S.; Lian, X.; Ciais, P.; Myneni, R.B.; Peñuelas, J.; Wang, T. Extension of the growing season increases vegetation exposure to frost. Nat. Commun. 2018, 9, 426. [Google Scholar] [CrossRef] [PubMed]
- Teskey, R.; Wertin, T.; Bauweraerts, I.; Ameye, M.; McGuire, M.A.; Steppe, K. Responses of tree species to heat waves and extreme heat events. Plant. Cell Environ. 2015, 38, 1699–1712. [Google Scholar] [CrossRef]
- Guha, A.; Han, J.; Cummings, C.; McLennan, D.A.; Warren, J.M. Differential ecophysiological responses and resilience to heat wave events in four co-occurring temperate tree species. Environ. Res. Lett. 2018, 13, 65008. [Google Scholar] [CrossRef]
- Van der Molen, M.K.; Dolman, A.J.; Ciais, P.; Eglin, T.; Gobron, N.; Law, B.E.; Meir, P.; Peters, W.; Phillips, O.L.; Reichstein, M. Drought and ecosystem carbon cycling. Agric. For. Meteorol. 2011, 151, 765–773. [Google Scholar] [CrossRef]
- Song, Y.; Njoroge, J.B. Drought impact assessment from monitoring the seasonality of vegetation condition using long-term time-series satellite images: A case study of Mt. Kenya region. Environ. Monit. Assess. 2013, 185, 4117–4124. [Google Scholar] [CrossRef]
- Stocker, B.D.; Zscheischler, J.; Keenan, T.F.; Prentice, I.C.; Seneviratne, S.I.; Peñuelas, J. Drought impacts on terrestrial primary production underestimated by satellite monitoring. Nat. Geosci. 2019, 12, 264. [Google Scholar] [CrossRef]
- Spittlehouse, D.; Stewart, R.B. Adaptation to climate change in forest management. JEM 2003, 4, 1–11. [Google Scholar]
- Quétier, F.; Lavorel, S. Assessing ecological equivalence in biodiversity offset schemes: Key issues and solutions. Biol. Conserv. 2011, 144, 2991–2999. [Google Scholar] [CrossRef]
- Piao, S.; Friedlingstein, P.; Ciais, P.; Zhou, L.; Chen, A. Effect of climate and CO2 changes on the greening of the Northern Hemisphere over the past two decades. Geophys. Res. Lett. 2006, 33, L23402. [Google Scholar] [CrossRef]
- Vicca, S. Global vegetation’s CO2 uptake. Nat. Ecol. Evol. 2018, 2, 1840. [Google Scholar] [CrossRef] [PubMed]
- Los, S.O. Analysis of trends in fused AVHRR and MODIS NDVI data for 1982–2006: Indication for a CO2 fertilization effect in global vegetation. Glob. Biogeochem. Cycles 2013, 27, 318–330. [Google Scholar] [CrossRef]
- Schimel, D.; Schneider, F.D.; Participants, J.C.A.E.; Bloom, A.; Bowman, K.; Cawse-Nicholson, K.; Elder, C.; Ferraz, A.; Fisher, J.; Hulley, G.; et al. Flux towers in the sky: Global ecology from space. New Phytol. 2019, 224, 570–584. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Full Name | Definition |
|---|---|---|
| GSL | Growing season length | Annual count between first span of at least 6 days with TG > 5 °C and first span after July 1 of 6 days with TG < 5 °C (TG: Temperature for growing season) |
| SU25 | Summer days | Annual count when TX (daily maximum) > 25 °C |
| FD | Frost days | Annual count when TN (daily minimum) < 0 °C |
| HW | Heat wave days | Annual count when TX (daily maximum) > 33 °C |
| SDII | Simple daily intensity index | Annual total precipitation divided by the number of wet days (defined as precipitation ≥ 1.0 mm) in the year |
| HR | Heavy rain days | Annual count when PRCP ≥ 80 mm |
| RX5 | Maximum consecutive 5-day precipitation | Let RRkj be the precipitation amount for the 5-day interval ending k, period j. Then, the maximum 5-day values for period j are Rx5dayj = max (RRkj) |
| SPEI12 | Standardized precipitation-evapotranspiration index | A site-specific drought indicator of deviations from average water balance (precipitation minus potential evapotranspiration). |
| LC | FD | GSL | HR | HW | SU | RX5 | SDII | SPEI12 | |
|---|---|---|---|---|---|---|---|---|---|
| CON | −0.27 | 0.46 | 0.05 | 0.01 | −0.20 | 0.03 | −0.13 | 0.29 | −0.02 |
| MIX | −0.34 | 0.19 | 0.02 | 0.08 | −0.08 | −0.09 | −0.08 | −0.07 | −0.04 |
| DEC | −0.53 | −0.01 | −0.05 | −0.05 | −0.06 | −0.11 | −0.03 | −0.06 | −0.03 |
| Elevation | CON | MIX | DEC |
|---|---|---|---|
| TOTAL | −0.27 | −0.34 | −0.53 |
| 0–200 m | −0.31 | −0.34 | −0.44 |
| 201–400 m | −0.10 | NA | NA |
| 401–500 m | NA | NA | NA |
| Elevation | Top 1 | Top 2 | |||
|---|---|---|---|---|---|
| CON | TOTAL | FD | 0.46 | SDII | 0.29 |
| 0–200 m | FD | 0.27 | HW | −0.27 | |
| 201–400 m | SDII | 0.46 | FD | 0.19 | |
| 401–500 m | SDII | 0.27 | SU | −0.16 | |
| MIX | TOTAL | FD | 0.19 | SU | −0.09 |
| 0–200 m | HW | −0.15 | HR | 0.15 | |
| 201–400 m | SDII | 0.33 | GSL | 0.11 | |
| 401–500 m | SDII | 0.18 | SU | −0.17 | |
| Relevant Policies | Planned Project or Regulation |
|---|---|
| Jeju climate change adaptation action plan (2012–2016) | Afforestation of tree species having economic benefit |
| Monitoring on protected area | |
| Maintenance of blight for Pinus species | |
| Prevention of and action on forest fires | |
| Research on forest succession | |
| Biodiversity offset scheme | Restrict development based on quality of biodiversity in spatial context |
| Landscape management decision-making considering the concept of biodiversity offset |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.J.; Song, Y.K.; Lee, D.K. Understanding the Critical Impact Path on Vegetation Growth under Climate Extremes and Human Influence. Forests 2019, 10, 947. https://doi.org/10.3390/f10110947
Kim YJ, Song YK, Lee DK. Understanding the Critical Impact Path on Vegetation Growth under Climate Extremes and Human Influence. Forests. 2019; 10(11):947. https://doi.org/10.3390/f10110947
Chicago/Turabian StyleKim, Yoon Jung, Young Keun Song, and Dong Kun Lee. 2019. "Understanding the Critical Impact Path on Vegetation Growth under Climate Extremes and Human Influence" Forests 10, no. 11: 947. https://doi.org/10.3390/f10110947
APA StyleKim, Y. J., Song, Y. K., & Lee, D. K. (2019). Understanding the Critical Impact Path on Vegetation Growth under Climate Extremes and Human Influence. Forests, 10(11), 947. https://doi.org/10.3390/f10110947