Does Drought Stress on Seedlings Have Longer Term Effects on Sapling Phenology, Reshooting, Growth and Plant Architecture in Quercus robur, Q. petraea and Their Morphological Intermediates?

 ,
,
Abstract
1. Introduction
2. Materials and Methods
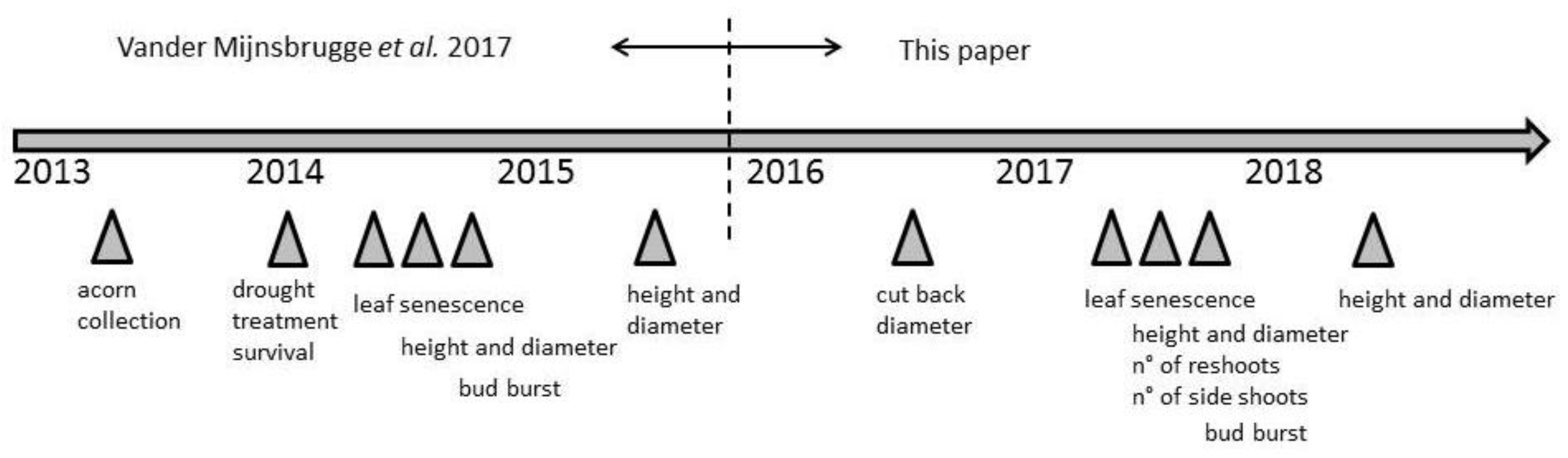
2.1. Source Material and Experimental Set-Up
2.2. Measurements and Scoring
2.3. Data Analysis
3. Results
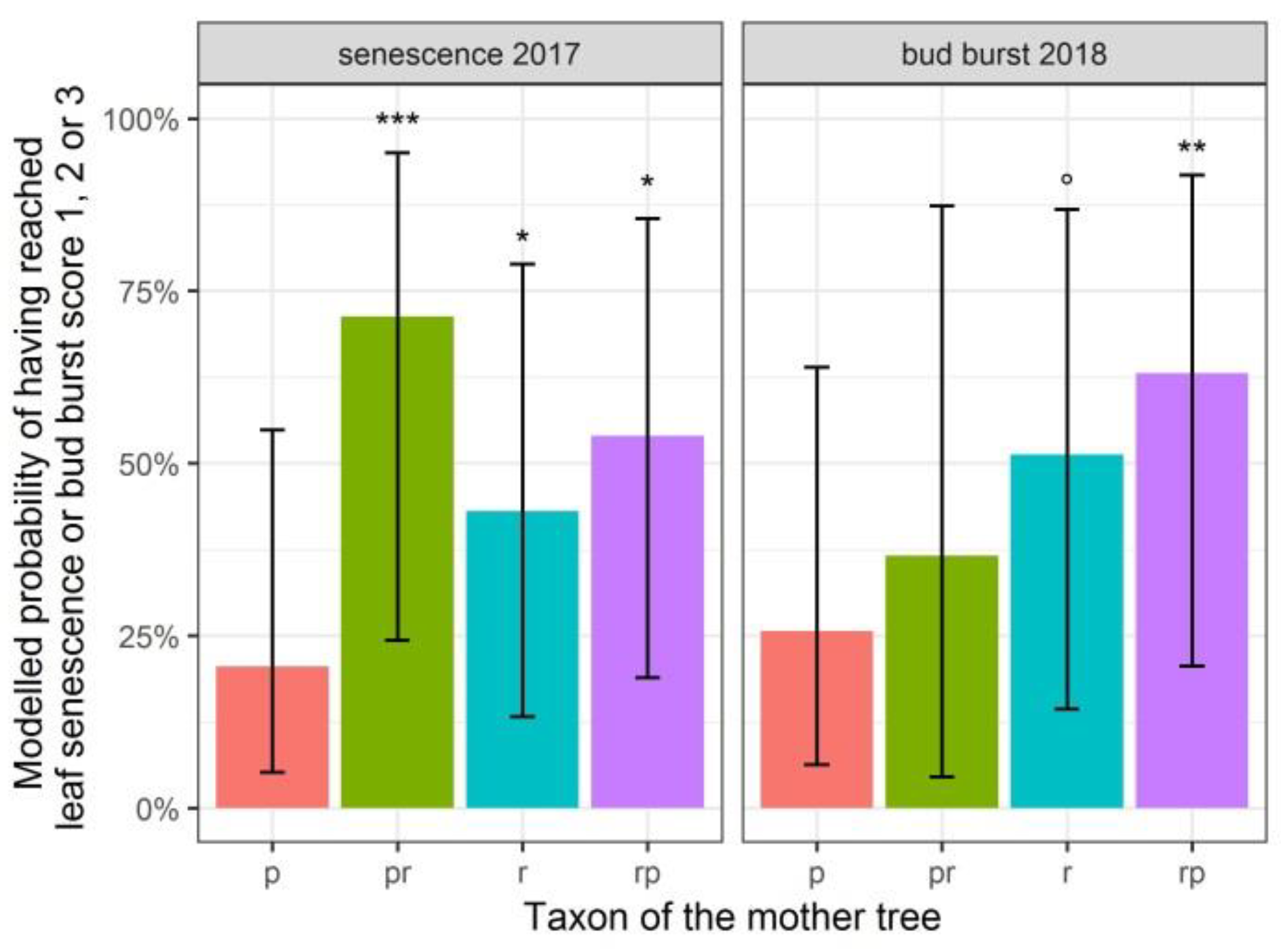
3.1. Leaf Senescence and Bud Burst
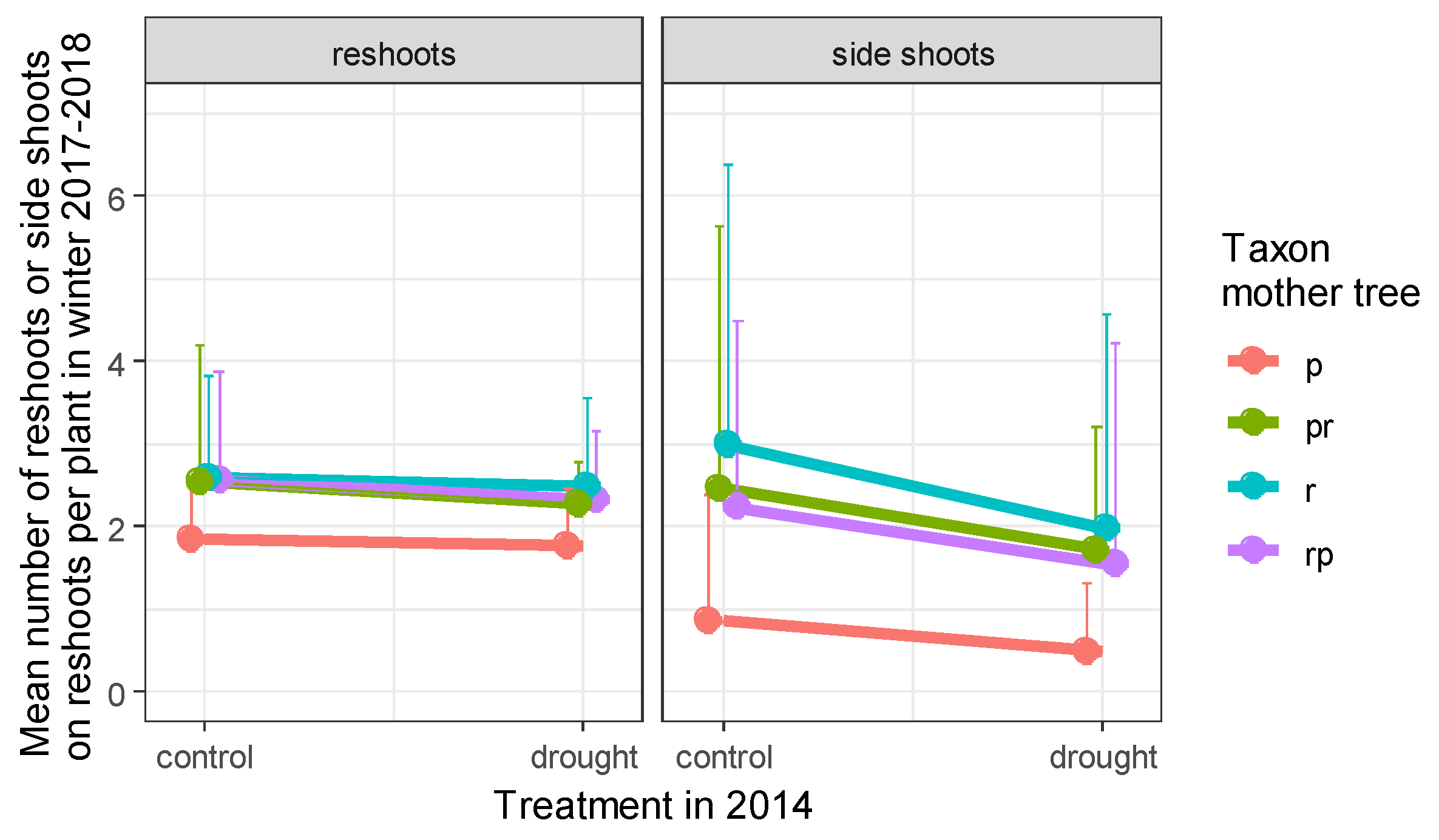
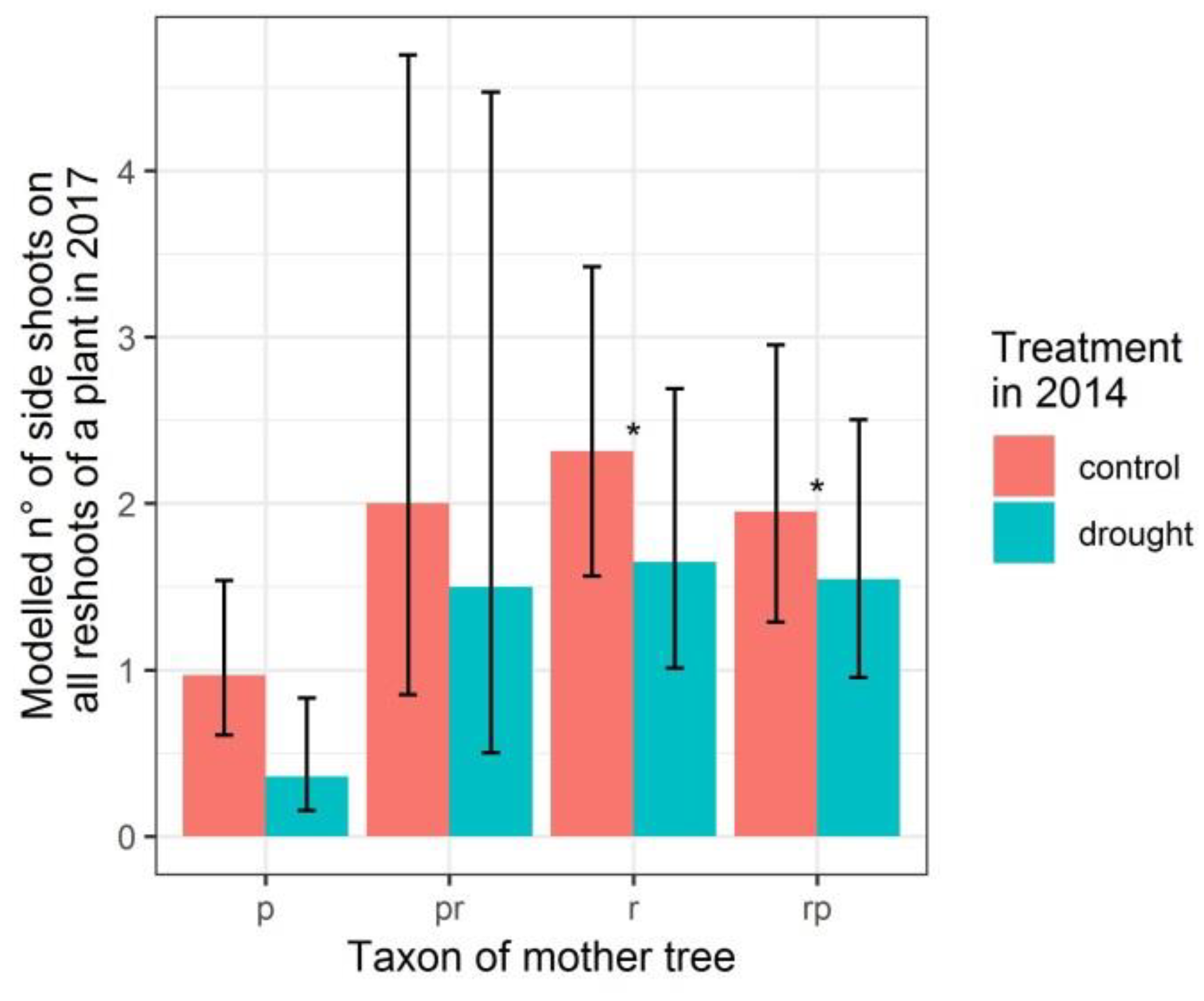
3.2. Number of Reshoots and Number of Side Shoots on Reshoots Per Plant
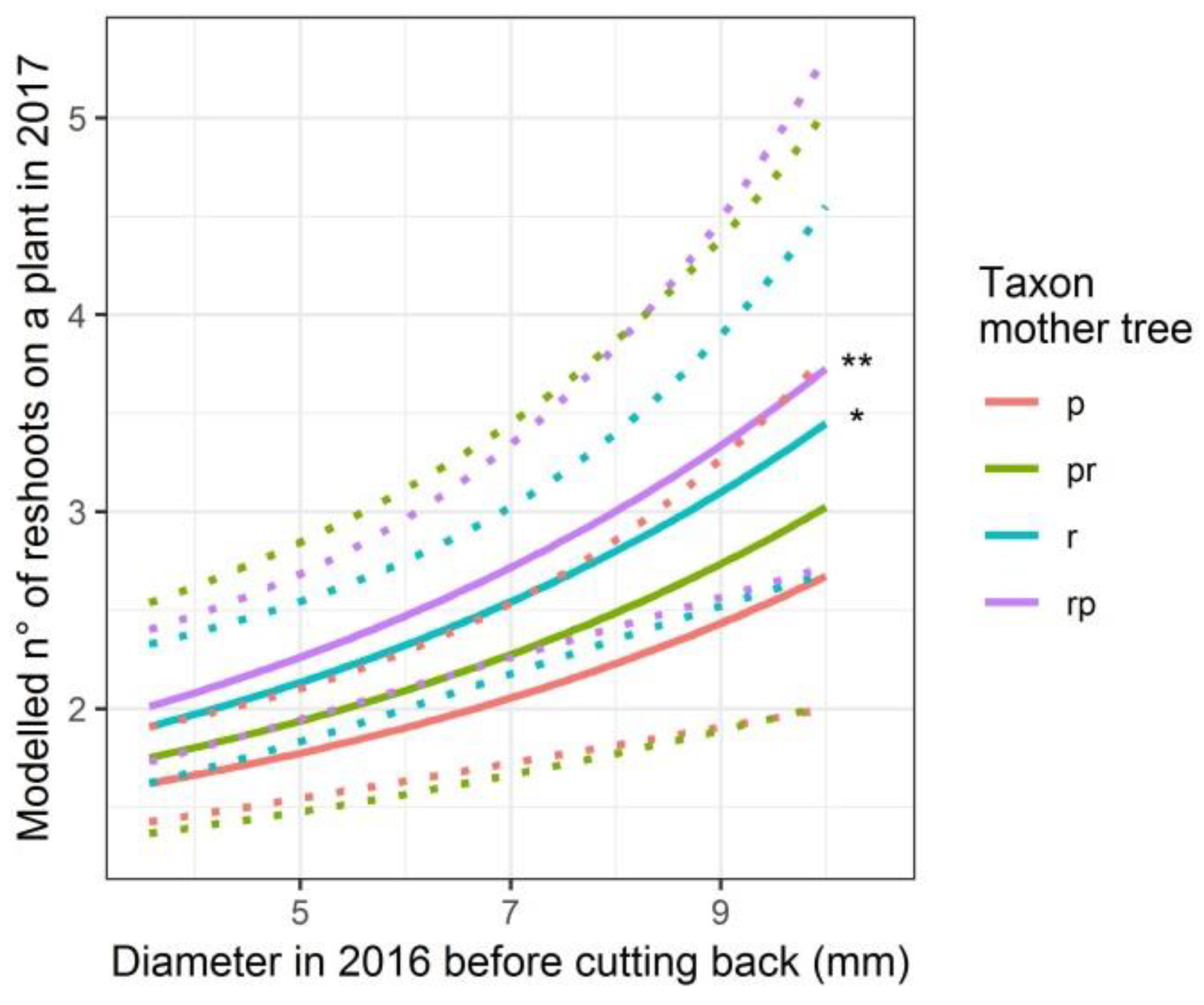
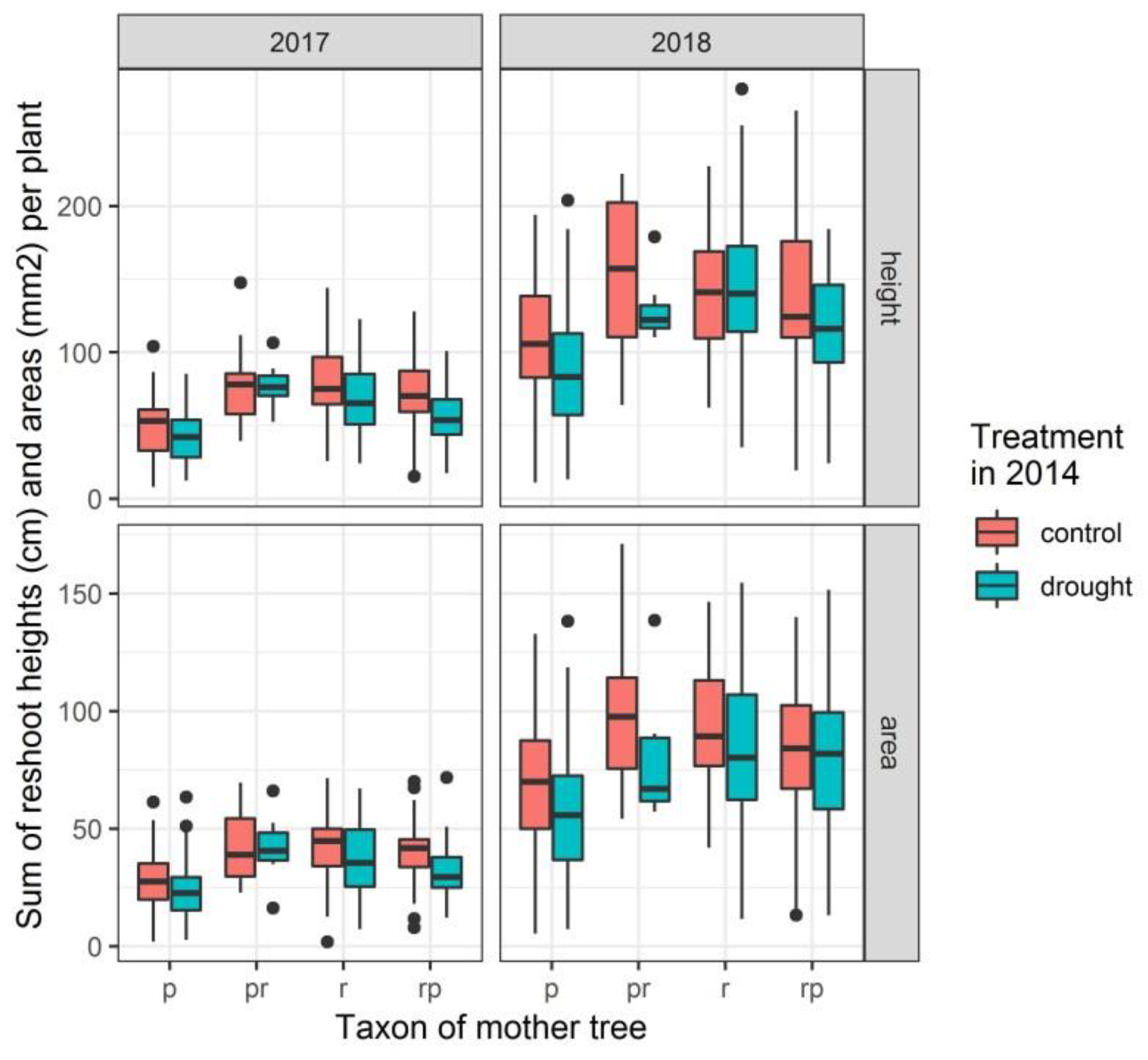
3.3. Height and Radial Growth of the Reshoots
4. Discussion
4.1. Faded Phenological Responses to Drought
4.2. Height and Basal Area
4.3. Plant Architecture
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Lindner, M.; Fitzgerald, J.B.; Zimmermann, N.E.; Reyer, C.; Delzon, S.; van der Maaten, E.; Schelhaas, M.J.; Lasch, P.; Eggers, J.; van der Maaten-Theunissen, M.; et al. Climate change and European forests: What do we know, what are the uncertainties, and what are the implications for forest management? J. Environ. Manag. 2014, 146, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Anderegg, W.R.L.; Berry, J.A.; Field, C.B. Linking definitions, mechanisms, and modeling of drought-induced tree death. Trends Plant. Sci. 2012, 17, 693–700. [Google Scholar] [CrossRef] [PubMed]
- Sheil, D. Forests, atmospheric water and an uncertain future: The new biology of the global water cycle. Ecosyst 2018, 5, 1–22. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Schwalm, C.; Biondi, F.; Camarero, J.J.; Koch, G.; Litvak, M.; Ogle, K.; Shaw, J.D.; Shevliakova, E.; Williams, A.P.; et al. Pervasive drought legacies in forest ecosystems and their implications for carbon cycle models. Science 2015, 349, 528–532. [Google Scholar] [CrossRef] [PubMed]
- Kannenberg, S.A.; Maxwell, J.T.; Pederson, N.; D’Orangeville, L.; Ficklin, D.L.; Phillips, R.P. Drought legacies are dependent on water table depth, wood anatomy and drought timing across the eastern us. Ecol. Lett. 2019, 22, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Hubert, F.; Grimm, G.W.; Jousselin, E.; Berry, V.; Franc, A.; Kremer, A. Multiple nuclear genes stabilize the phylogenetic backbone of the genus Quercus. Syst. Biodivers. 2014, 12, 405–423. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schutze, G.; Uhl, E. Resistance of European tree species to drought stress in mixed versus pure forests: Evidence of stress release by inter-specific facilitation. Plant. Biol. 2013, 15, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Epron, D.; Dreyer, E. Long-term effects of drought on photosynthesis of adult oak trees [Quercus petraea (Matt) Liebl and Quercus robur L] in a natural stand. New Phytol. 1993, 125, 381–389. [Google Scholar] [CrossRef]
- Leuschner, C.; Backes, K.; Hertel, D.; Schipka, F.; Schmitt, U.; Terborg, O.; Runge, M. Drought responses at leaf, stem and fine root levels of competitive Fagus sylvatica L. and Quercus petraea (Matt.) Liebl. Trees in dry and wet years. For. Ecol. Manag. 2001, 149, 33–46. [Google Scholar] [CrossRef]
- Levy, G.; Becker, M.; Duhamel, D. A comparison of the ecology of pedunculate and sessile oaks-radial growth in the center and northwest of france. For. Ecol. Manag. 1992, 55, 51–63. [Google Scholar] [CrossRef]
- Cochard, H.; Breda, N.; Granier, A.; Aussenac, G. Vulnerability to air-embolism of 3 European oak species (Quercus petraea (Matt) Liebl, Quercus pubescens Willd, Quercus robur L.). Ann. Sci. For. 1992, 49, 225–233. [Google Scholar] [CrossRef]
- Vivin, P.; Aussenac, G.; Levy, G. Differences in drought resistance among 3 deciduous oak species grown in large boxes. Ann. Sci. For. 1993, 50, 221–233. [Google Scholar] [CrossRef]
- Kleinschmit, J.R.G.; Bacilieri, R.; Kremer, A.; Roloff, A. Comparison of morphological and genetic traits of pedunculate oak (Q. robur L.) and sessile oak (Q. petraea (Matt) Liebl). Silvaec. Genet. 1995, 44, 256–269. [Google Scholar]
- Vanoni, M.; Bugmann, H.; Notzli, M.; Bigler, C. Quantifying the effects of drought on abrupt growth decreases of major tree species in Switzerland. Ecol. Evol. 2016, 6, 3555–3570. [Google Scholar] [CrossRef] [PubMed]
- Thomas, F.M.; Gausling, T. Morphological and physiological responses of oak seedlings (Quercus petraea and Q. robur) to moderate drought. Ann. Sci. 2000, 57, 325–333. [Google Scholar] [CrossRef]
- Scheidegger, Y.; Saurer, M.; Bahn, M.; Siegwolf, R. Linking stable oxygen and carbon isotopes with stomatal conductance and photosynthetic capacity: A conceptual model. Oecologia 2000, 125, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Perkins, D.; Uhl, E.; Biber, P.; du Toit, B.; Carraro, V.; Rotzer, T.; Pretzsch, H. Impact of climate trends and drought events on the growth of oaks (Quercus robur L. and Quercus petraea (Matt.) Liebl.) within and beyond their natural range. Forests 2018, 9, 108. [Google Scholar] [CrossRef]
- Spiess, N.; Oufir, M.; Matusikova, I.; Stierschneider, M.; Kopecky, D.; Homolka, A.; Burg, K.; Fluch, S.; Hausman, J.F.; Wilhelm, E. Ecophysiological and transcriptomic responses of oak (Quercus robur) to long-term drought exposure and rewatering. Environ. Exp. Bot. 2012, 77, 117–126. [Google Scholar] [CrossRef]
- Jensen, J.S.; Hansen, J.K. Genetic variation in responses to different soil water treatments in Quercus robur L. Scand. J. For. Res. 2010, 25, 400–411. [Google Scholar] [CrossRef]
- Pflug, E.E.; Siegwolf, R.; Buchmann, N.; Dobbertin, M.; Kuster, T.M.; Gunthardt-Goerg, M.S.; Arend, M. Growth cessation uncouples isotopic signals in leaves and tree rings of drought-exposed oak trees. Tree Physiol. 2015, 35, 1095–1105. [Google Scholar] [CrossRef] [PubMed]
- Kuster, T.M.; Dobbertin, M.; Gunthardt-Goerg, M.S.; Schaub, M.; Arend, M. A phenological timetable of oak growth under experimental drought and air warming. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Vander Mijnsbrugge, K.; Turcsan, A.; Maes, J.; Duchene, N.; Meeus, S.; Steppe, K.; Steenackers, M. Repeated summer drought and re-watering during the first growing year of oak (Quercus petraea) delay autumn senescence and bud burst in the following spring. Front. Plant. Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Niinemets, U. Responses of forest trees to single and multiple environmental stresses from seedlings to mature plants: Past stress history, stress interactions, tolerance and acclimation. For. Ecol. Manag. 2010, 260, 1623–1639. [Google Scholar] [CrossRef]
- Psidova, E.; Ditmarova, L.; Jamnicka, G.; Kurjak, D.; Majerova, J.; Czajkowski, T.; Bolte, A. Photosynthetic response of beech seedlings of different origin to water deficit. Photosynthetica 2015, 53, 187–194. [Google Scholar] [CrossRef]
- Brouwers, J.; Peeters, B.; Van Steertegem, M.; van Lipzig, N.; Wouters, H.; Beullens, J.; Demuzere, M.; Willems, P.; De Ridder, K.; Maiheu, B.; et al. Mira Climate Report 2015: About Observed and Future Climate Changes in Flanders and Belgium; Flanders Environment Agency: Aalst, Belgium, 2015. [Google Scholar]
- Vander Mijnsbrugge, K.; Turcsan, A.; Maes, J.; Duchene, N.; Meeus, S.; Van der Aa, B.; Steppe, K.; Steenackers, M. Taxon-independent and taxon-dependent responses to drought in seedlings from Quercus robur L., Q. -petraea (Matt.) Liebl. and their morphological intermediates. Forests 2017, 8, 407. [Google Scholar]
- Büsgen, M.; Münch, E.; Thomson, T. Structure Life Forest Trees; Chapman and Hall: London, UK, 1929. [Google Scholar]
- Tainter, F.H.; Retzlaff, W.A.; Starkey, D.A.; Oak, S.W. Decline of radial growth in red oaks is associated with short-term changes in climate. Eur. J. Pathol. 1990, 20, 95–105. [Google Scholar] [CrossRef]
- Bacilieri, R.; Ducousso, A.; Petit, R.J.; Kremer, A. Mating system and asymmetric hybridization in a mixed stand of European oaks. Evolution 1996, 50, 900–908. [Google Scholar] [CrossRef] [PubMed]
- Vander Mijnsbrugge, K.; Cox, K.; Van Slycken, J. Conservation approaches for autochthonous woody plants in Flanders. Silvaec. Genet. 2005, 54, 197–206. [Google Scholar] [CrossRef]
- Vander Mijnsbrugge, K.; Coart, E.; Beeckman, H.; Van Slycken, J. Conservation measures for autochthonous oaks in Flanders. For. Genet. 2003, 10, 207–217. [Google Scholar]
- Vander Mijnsbrugge, K.; De Cleene, L.; Beeckman, H. A combination of fruit and leaf morphology enables taxonomic classification of the complex Q. robur L.-Q. × rosacea Bechst.-Q. petraea (Matt.) Liebl. in autochthonous stands in flanders. Silvaec. Genet. 2011, 60, 139–148. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing. R Foundation for Statistical Computing; Vienna, Austria. 2014. Available online: http://www.R-project.org/ (accessed on 5 July 2019).
- Christensen, R.H.B. Ordinal: Regression Models for Ordinal Data. R Package Version. 2013. Available online: http://www.Cran.R-project.Org/package=ordinal/ (accessed on 31 October 2013).
- Bates, D.; Machler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S; Springer: New York, NY, USA, 2002. [Google Scholar]
- Alberto, F.J.; Aitken, S.N.; Alia, R.; Gonzalez-Martinez, S.C.; Hanninen, H.; Kremer, A.; Lefevre, F.; Lenormand, T.; Yeaman, S.; Whetten, R.; et al. Potential for evolutionary responses to climate change - Evidence from tree populations. Glob. Chang. Biol. 2013, 19, 1645–1661. [Google Scholar] [CrossRef] [PubMed]
- Milne, R.I.; Abbott, R.J. Origin and evolution of invasive naturalized material of Rhododendron ponticum L. in the British Isles. Mol. Ecol. 2000, 9, 541–556. [Google Scholar] [CrossRef] [PubMed]
- Gieger, T.; Thomas, F.M. Effects of defoliation and drought stress on biomass partitioning and water relations of Quercus robur and Quercus petraea. Basic Appl. Ecol. 2002, 3, 171–181. [Google Scholar] [CrossRef]
- Umehara, M.; Hanada, A.; Yoshida, S.; Akiyama, K.; Arite, T.; Takeda-Kamiya, N.; Magome, H.; Kamiya, Y.; Shirasu, K.; Yoneyama, K.; et al. Inhibition of shoot branching by new terpenoid plant hormones. Nature 2008, 455, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.J.; Wang, X.P.; Weighill, D.A.; Guo, H.B.; Xie, M.; Yang, Y.G.; Yang, J.; Wang, S.C.; Jacobson, D.A.; Guo, H.; et al. Characterization of dwarf14 genes in Populus. Sci. Rep. UK 2016, 6, 20593. [Google Scholar] [CrossRef] [PubMed]
- Ha, C.V.; Leyva-Gonzalez, M.A.; Osakabe, Y.; Tran, U.T.; Nishiyama, R.; Watanabe, Y.; Tanaka, M.; Seki, M.; Yamaguchi, S.; Dong, N.V.; et al. Positive regulatory role of strigolactone in plant responses to drought and salt stress. Proc. Natl. Acad. Sci. USA 2014, 111, 851–856. [Google Scholar] [CrossRef] [PubMed]
- Ammer, C. Growth and biomass partitioning of fagus sylvatica l. And quercus robur l. Seedlings in response to shading and small changes in the r/fr-ratio of radiation. Ann. Sci. 2003, 60, 163–171. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Taxon Mother Tree * | n2014 | n2015 | n2017 |
|---|---|---|---|---|
| control | p | 69 | 68 | 60 |
| pr | 13 | 13 | 11 | |
| rp | 55 | 55 | 53 | |
| r | 58 | 58 | 51 | |
| drought | p | 74 | 50 | 42 |
| pr | 14 | 11 | 7 | |
| rp | 58 | 38 | 35 | |
| r | 51 | 36 | 36 |
| Phenophase | Score Level | Description |
|---|---|---|
| leaf senescence | 1 | light green leaves |
| 2 | light green leaves with yellow parts | |
| 3 | yellow leaves still having green parts | |
| 4 | yellow leaves with brown parts | |
| 5 | brown leaves still having yellow parts | |
| 6 | brown leaves | |
| bud burst | 1 | apical bud in winter rest |
| 2 | apical bud swollen | |
| 3 | apical bud opening, leaves not yet protruding | |
| 4 | leaves protruding but not yet unfolding | |
| 5 | leaves unfolding but not yet fully unfolded | |
| 6 | leaves fully unfolded |
| Leaf Senescence 2017 | Bud Burst 2018 | |||||||
|---|---|---|---|---|---|---|---|---|
| Variable | Estimate | Std. error | z-Value | p-Value | Estimate | Std. Error | z-Value | p-Value |
| Tpr | −2.61 | 0.74 | −3.52 | <0.001 *** | 0.66 | 0.92 | 0.72 | 0.472 |
| Tr | −1.02 | 0.44 | −2.33 | 0.020 * | 0.94 | 0.53 | 1.78 | 0.074. |
| Trp | −0.92 | 0.45 | −2.03 | 0.043 * | 1.50 | 0.54 | 2.79 | 0.005 ** |
| S | −0.27 | 0.36 | −0.74 | 0.461 | −0.31 | 0.40 | −0.79 | 0.432 |
| H | 0.00 | 0.00 | 0.88 | 0.382 | 0.00 | 0.00 | 1.01 | 0.315 |
| S:Tpr | 0.34 | 0.92 | 0.37 | 0.712 | −0.15 | 0.99 | −0.15 | 0.878 |
| S:Tr | −0.06 | 0.56 | −0.11 | 0.912 | 0.17 | 0.58 | 0.30 | 0.766 |
| S:Trp | −0.60 | 0.54 | −1.11 | 0.266 | 0.10 | 0.55 | 0.19 | 0.849 |
| n° of Reshoots per Plant (2017) | n° of Side Shoots on Reshoots per Plant (2017) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Variable | Estimate | Std. Error | z−Value | p−Value | Variable | Estimate | Std. Error | t−Value | p−Value |
| (Intercept) | −1.11 | 0.30 | −3.75 | <0.001 *** | (Intercept) | −0.12 | 0.29 | −0.41 | 0.681 |
| Tpr | 0.33 | 0.29 | 1.15 | 0.252 | Tpr | 0.73 | 0.50 | 1.44 | 0.172 |
| Tr | 0.40 | 0.19 | 2.08 | 0.038 * | Tr | 0.87 | 0.32 | 2.77 | 0.015 * |
| Trp | 0.54 | 0.19 | 2.91 | 0.004 ** | Trp | 0.70 | 0.32 | 2.17 | 0.048 * |
| S | 0.08 | 0.23 | 0.35 | 0.730 | S | −0.99 | 0.47 | −2.11 | 0.036 * |
| D | 0.15 | 0.04 | 3.76 | <0.001 *** | H | 0.00 | 0.00 | 0.54 | 0.590 |
| S:Tpr | −0.14 | 0.47 | −0.30 | 0.764 | S:Tpr | 0.70 | 0.76 | 0.93 | 0.354 |
| S:Tr | −0.02 | 0.29 | −0.05 | 0.958 | S:Tr | 0.65 | 0.53 | 1.22 | 0.222 |
| S:Trp | −0.05 | 0.30 | −0.17 | 0.868 | S:Trp | 0.76 | 0.54 | 1.41 | 0.161 |
| Taxon Mother Tree | Variable | Estimate | St. Error | df | t−Value | p−Value |
|---|---|---|---|---|---|---|
| Q. petraea | (Intercept) | 49.90 | 6.64 | 97 | 7.52 | <0.001 *** |
| S | −13.96 | 5.49 | 95 | −2.54 | 0.013 * | |
| Y | 53.69 | 3.02 | 97 | 17.80 | <0.001 *** | |
| long stalked intermediate | (Intercept) | 79.82 | 10.85 | 16 | 7.35 | <0.001 *** |
| S | −9.24 | 16.03 | 16 | −0.58 | 0.572 | |
| Y | 66.05 | 8.77 | 16 | 7.53 | <0.001 *** | |
| Q. robur | (Intercept) | 76.74 | 6.01 | 72 | 12.77 | <0.001 *** |
| S | −5.27 | 8.30 | 69 | −0.63 | 0.528 | |
| Y | 71.07 | 3.81 | 72 | 18.65 | <0.001 *** | |
| short stalked intermediate | (Intercept) | 71.71 | 6.05 | 86 | 11.86 | <0.001 *** |
| S | −14.77 | 6.89 | 81 | −2.14 | 0.035 * | |
| Y | 59.51 | 3.57 | 86 | 16.67 | <0.001 *** |
| Taxon mother tree | Variable | Estimate | St. Error | df | t−Value | p−Value |
|---|---|---|---|---|---|---|
| Q. petraea | (Intercept) | 2.86 | 0.44 | 96 | 6.45 | <0.001 *** |
| S | −0.85 | 0.36 | 95 | −2.34 | 0.021 * | |
| Y | 3.77 | 0.20 | 96 | 18.69 | <0.001 *** | |
| long stalked intermediate | (Intercept) | 4.64 | 0.67 | 32 | 6.89 | <0.001 *** |
| S | −0.99 | 0.86 | 32 | −1.15 | 0.257 | |
| Y | 4.92 | 0.84 | 32 | 5.86 | <0.001 *** | |
| Q. robur | (Intercept) | 4.36 | 0.53 | 71 | 8.16 | <0.001 *** |
| S | −0.89 | 0.46 | 69 | −1.92 | 0.059 | |
| Y | 4.87 | 0.25 | 71 | 19.26 | <0.001 *** | |
| short stalked intermediate | (Intercept) | 3.87 | 0.34 | 86 | 11.42 | <0.001 *** |
| S | −0.60 | 0.44 | 81 | −1.36 | 0.177 | |
| Y | 4.60 | 0.25 | 86 | 18.51 | <0.001 *** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vander Mijnsbrugge, K.; Turcsan, A.; Moreels, S.; Van Goethem, M.; Meeus, S.; Van der Aa, B. Does Drought Stress on Seedlings Have Longer Term Effects on Sapling Phenology, Reshooting, Growth and Plant Architecture in Quercus robur, Q. petraea and Their Morphological Intermediates? Forests 2019, 10, 1012. https://doi.org/10.3390/f10111012
Vander Mijnsbrugge K, Turcsan A, Moreels S, Van Goethem M, Meeus S, Van der Aa B. Does Drought Stress on Seedlings Have Longer Term Effects on Sapling Phenology, Reshooting, Growth and Plant Architecture in Quercus robur, Q. petraea and Their Morphological Intermediates? Forests. 2019; 10(11):1012. https://doi.org/10.3390/f10111012
Chicago/Turabian StyleVander Mijnsbrugge, Kristine, Arion Turcsan, Stefaan Moreels, Michiel Van Goethem, Steven Meeus, and Beatrijs Van der Aa. 2019. "Does Drought Stress on Seedlings Have Longer Term Effects on Sapling Phenology, Reshooting, Growth and Plant Architecture in Quercus robur, Q. petraea and Their Morphological Intermediates?" Forests 10, no. 11: 1012. https://doi.org/10.3390/f10111012
APA StyleVander Mijnsbrugge, K., Turcsan, A., Moreels, S., Van Goethem, M., Meeus, S., & Van der Aa, B. (2019). Does Drought Stress on Seedlings Have Longer Term Effects on Sapling Phenology, Reshooting, Growth and Plant Architecture in Quercus robur, Q. petraea and Their Morphological Intermediates? Forests, 10(11), 1012. https://doi.org/10.3390/f10111012