Modeling Migratory Flight in the Spruce Budworm: Circadian Rhythm


1
Natural Resources Canada, Canadian Forest Service, 1055 PEPS street, Quebec, QC G1V 4C7, Canada
2
Department of Forest and Wildlife Ecology, University of Wisconsin-Madison, Madison, WI 53706, USA
*
Author to whom correspondence should be addressed.
Forests 2019, 10(10), 877; https://doi.org/10.3390/f10100877
Submission received: 30 July 2019
/
Revised: 26 September 2019
/
Accepted: 30 September 2019
/
Published: 5 October 2019
(This article belongs to the Special Issue Protection Strategy against Spruce Budworm)
Abstract
:The crepuscular (evening) circadian rhythm of adult spruce budworm (Choristoneura fumiferana (Clem.)) flight activity under the influence of changing evening temperatures is described using a mathematical model. This description is intended for inclusion in a comprehensive model of spruce budworm flight activity leading to the simulation of mass migration events. The model for the temporal likelihood of moth emigration flight is calibrated using numerous observations of flight activity in the moth’s natural environment. Results indicate an accurate description of moth evening flight activity using a temporal function covering the period around sunset and modified by evening temperature conditions. The moth’s crepuscular flight activity is typically coincident with the evening transition of the atmospheric boundary layer from turbulent daytime to stable nocturnal conditions. The possible interactions between moth flight activity and the evening boundary layer transition, with favorable wind and temperature conditions leading to massive and potentially successful migration events, as well as the potential impact of climate change on this process, are discussed.
1. Introduction
Migration is a biological imperative observed in many animals and generally well understood in ecological and evolutionary terms [1]. This paper focuses on observations of circadian rhythms of migratory flight activity in the adult spruce budworm (Choristoneura fumiferana (Clem.)) (Lepidoptera: Tortricidae) with consideration of near-surface and boundary layer meteorological processes. In a companion paper, we discussed the basic physics of spruce budworm (SBW) flight and developed a mathematical framework with which we can model adult moth migration [2]. The ultimate objective of this work is the development of a simulation model to allow both historical analysis and real-time prediction of SBW population dispersal though moth migration and oviposition. We aim for a model that accounts for source population distribution and density, seasonal phenology, circadian rhythms of activity, and interactions with meteorological conditions at the surface (in the host forest stands) and within the atmospheric boundary layer.
Numerous activities in moths and butterflies have been related to daily cycles of behavior and are often classified into diurnal (daytime), nocturnal (nighttime), and crepuscular (twilight, at dawn and/or dusk) rhythms [3,4]. These rhythms may be truly circadian, driven by an internal (endogenous) biological clock, or diel rhythms that are driven by exogenous cues such as light and temperature cycles [5]. The primary difference between these two types is that circadian rhythms continue, though not necessarily on a 24-h cycle, regardless of external cues, while diel rhythms may shift with a change in exogenous conditions or cease entirely in the absence of such cues. Circadian rhythms may be entrained by exogenous cues, such as the diurnal cycle of light intensity and temperature, and might otherwise drift, but these cycles remain active even in the absence of external cues [5]. In insects, the entraining cues may include the availability of food resources [6], light intensity [7,8,9,10], temperature [11], humidity [5], sex-related differences such as pheromone release and its own dependence on temperature [12,13,14], and predation risk [15,16,17].
The SBW moth’s daily pattern of flight activity is a typical case of crepuscular circadian rhythm [18,19,20]. The adult SBW is poikilothermic [21] with its activity dependent on an optimal range of temperatures in the host forest. Temperature affects the timing of diel periodicities in the behavior of many lepidoptera [16], though a tendency toward purely diurnal or nocturnal activity may be suppressed by high or low temperatures during those times of day, respectively. In the adult SBW, male and female emigration flight is centered around sunset, with peak activity for males about 1.5 h later than for females, depending often on weather-related influences from temperature, wind speed, humidity, cloudiness, rainfall, and barometric pressure [20]. It is important here to distinguish SBW migratory flight behavior from activities such as mating. Environmental conditions that may not be conducive to flight above the forest canopy may still promote levels of flight activity sufficient for mating within the forest canopy, and typically on the same crepuscular cycle [18] but also displaying a minor peak in the morning hours [20]. However, successful migration flights depend on an alignment of both the timing of SBW moth activity and favorable wind and temperature conditions in the atmospheric boundary layer above the forest canopy. Because mating occurs soon after adult emergence [10] and mated and fully gravid females are usually too heavy to fly until at least their first cycle of oviposition [10,22], we may infer two things when we witness migration events. First, conditions in both the forest canopy and the atmospheric boundary layer favor flight activity at that time. Second, conditions on prior evenings may not have been favorable for flight above the forest canopy but at least supported mating activity and initial oviposition that also depend on temperature, so that lighter, less-gravid females can undertake migration.
The time of day at which SBW moths depart on their migratory flights therefore depends on crepuscular circadian rhythms and determines, to a large extent, the atmospheric conditions under which this movement is attempted and the distance that the insects travel [10]. We should perhaps differentiate here between migration on clear nights, which the mathematical model described below addresses, and migration flights that may be triggered by cues such as the passage of a cold front [23,24] or thunderstorm surface outflow [8,25]. Both of these potential triggers are commonly observed over land areas in the late afternoon and evening hours in the summer, when the crepuscular activity of SBW moths is at its peak. Historical observation of a SBW migration flight triggered by a solar eclipse suggests that cues other than time of day can influence the timing of migratory flight in this species [11]. Observations using both entomological and meteorological radar [26,27,28,29,30,31,32,33] have revealed the behavior of many migrating insects; such observations may enable the detection of moth dispersal events triggered under particular weather conditions (e.g. storm winds and the accompanying precipitation). However, it is not clear whether the high turbulence associated with fronts and storm outflows is a trigger for moth migration or simply facilitates the liftoff of a larger proportion of moths in strong updrafts. In many cases, the moths in flight can be drawn into frontal circulations and convective cells and subjected to low temperatures or wetting in rain events. Such conditions cause the SBW moth to fold its wings and drop to the surface [8], ending the migration flight and potentially leading to high mortality. Massive and successful migration events have been observed on generally calm evenings following a hot afternoon, progressing late into the night, with favorable (i.e., mild-to-strong) and coherent winds above the forest canopy [32].
For our effort at flight modeling, we relate the more general crepuscular circadian rhythm of the SBW in its adult stage, including behaviors such as mating within the forest canopy, to its eventual evening liftoff, flight above the canopy, and potentially successful nocturnal migration. In this paper we develop a mathematical model to relate SBW crepuscular activity to time of day and temperature, and we calibrate this model with observations of SBW moth flights made from scaffolds in North American boreal forest stands under natural conditions. The applications of these findings to our model of SBW migratory flight are then discussed.
2. Materials and Methods
2.1. Mathematical Model
We consider two factors as determinants of migratory flight of the SBW in this model: Time of day and temperature. Temperature directly affects the flight activity of SBW moths through wingbeat frequency [2]. The literature also suggests that migratory flight may be inhibited when temperature remains above a certain threshold well into the night, and that migratory flight does not occur much after midnight [10].
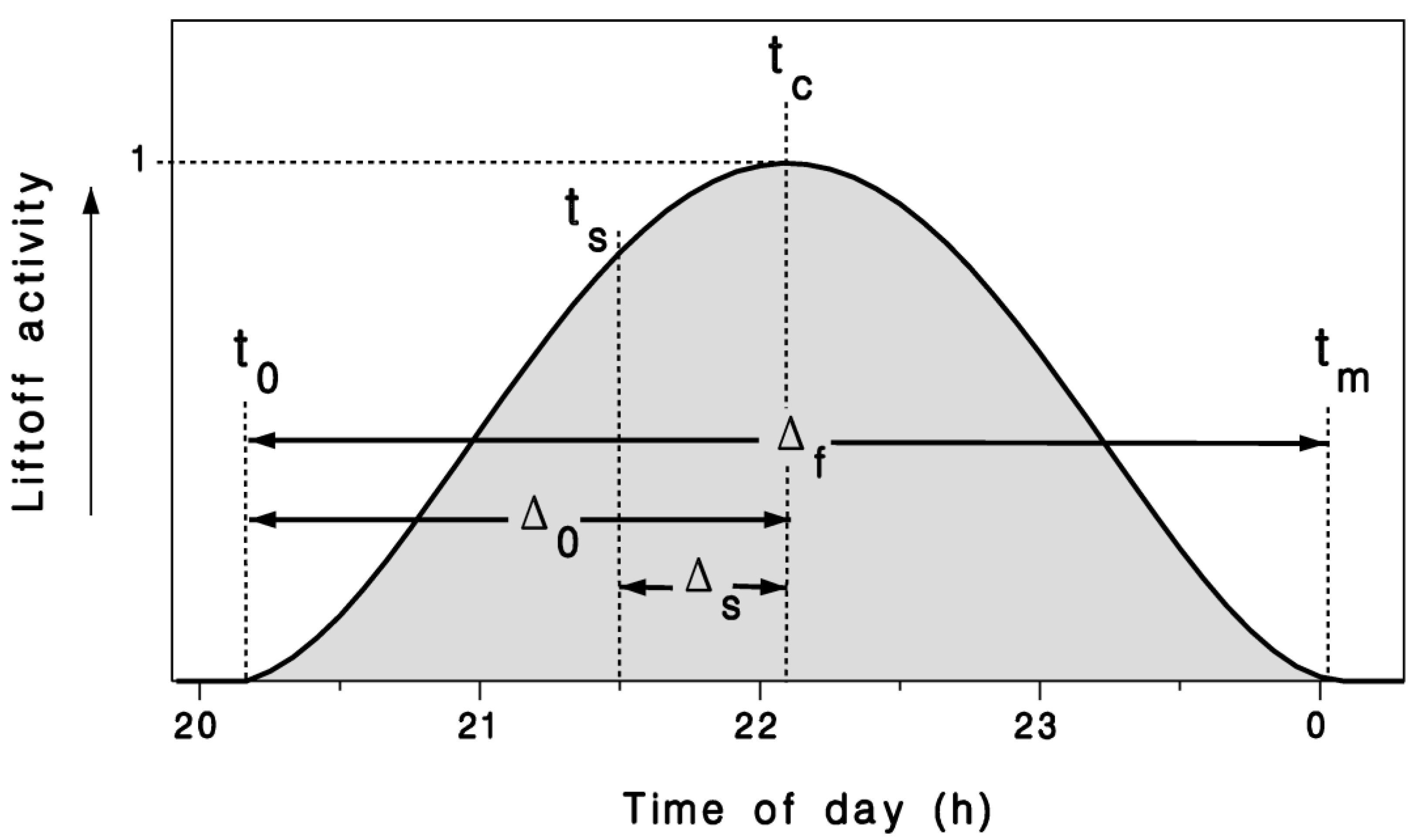
We express the circadian rhythm of moth liftoff activity as a quasi-Gaussian pattern centered near (though not necessarily at) the time of sunset ts. This pattern can be shifted by ambient temperature conditions. We thus develop a time transformation τ around the central time tc
with the start and end of the liftoff period denoted t0 and tm, respectively, and the full likelihood distribution of moth liftoff described by
where ft = 1 is the maximum at t = tc or τ = 0 (Figure 1).
The time difference between the onset of the moth liftoff period t0 and the time of peak likelihood tc is used to define Δ0 (in hours):
The end of the liftoff period tm can be used with the onset time t0 to define the full duration of liftoff activity Δf (in hours):
The function in Figure 1 is not necessarily symmetric about the peak time tc, by virtue of Equation (1). Expecting the peak of the moth liftoff period to occur after the time of local sunset, we can define the time difference Δs (in hours):
where the sunset time ts depends on latitude and time of year.
Equation (2) can be viewed as a probability density function for moth liftoff depending on time-of-day and subject to the offsets and durations defined in Equations (3)–(5). To use this in a discrete time simulation context, we need to express Equation (2) as a cumulative distribution function ranging from 0 at t0 to 1 at tm so that we can calculate the probability of moth liftoff in a short interval. This definite integral is given by:
where C = 1 − 2/3 + 1/5 = 0.533333 is a constant of integration ensuring that Ft remains strictly in the range (0,1) for τ (defined by Equation (1)) in the range (−1, 1). The probability of a moth taking off in an interval Δτ is therefore:
To completely specify this model, we must estimate the values of the three duration and offset parameters: Δ0 for Equation (3), Δf for Equation (4), and Δs for Equation (5).
The circadian likelihood of a moth attempting liftoff in a time interval Δτ is given by Equation (7), but the actual physical capability for flight is determined by temperature with respect to the moth’s weight and forewing area. It was established in [2] that wingbeat frequency is a sigmoid function of ambient temperature T (°C),
and that the wingbeat frequency required for moth liftoff is a function of its weight M and (single) forewing surface area A,
The actual diel pattern of migration on any given evening is therefore a potentially complex interaction between the circadian rhythm of activity (given by Equation (7)) and the effect of decreasing evening temperature on flight capability (Equations (8) and (9)). Because we are concerned with the deposition of eggs at the end of a migratory flight, leading to the next generation of SBW larvae in potentially new locations, we also calculate the proportion of initial fecundity carried away by emigrating females (G) based on their mass and forewing area (see [2]):
2.2. Observations and Model Calibration
The time of local sunset and the ambient temperature are external inputs to this system, but several terms defining Equations (3)–(5) (Δ0, Δf, and Δs, respectively) must be determined from observations. For this purpose, we assembled 22 historical datasets of diel SBW moth flight observations. Two of these datasets were published previously by Greenbank et al. [10]. The other 20 datasets were obtained from similar observations made in the vicinity of Sault Ste. Marie and Black Sturgeon Lake, ON, by the late C.J. Sanders and his team in 1976 (two datasets), 1987 (eight datasets), and 1989 (10 datasets). In all cases, the number of moths ascending into emigration flight was observed over short periods of time (typically 5 min) from tall scaffolds above the canopy and recorded at intervals of 15 min from early evening (prior to sunset) to total darkness. On some nights, night-vision (infrared) devices were used after total darkness to observe late flying moths, primarily to ensure that those were not numerous. Air temperature was recorded at the scaffold location at the same frequency. Sunset times for each location and observation date were obtained from www.timeanddate.com/sun/canada/ (last accessed 1 October 2019). Because these canopy-level observations did not distinguish between males and females, and because Greenbank et al. [10] reported that the sex ratio of moths caught during emigration liftoff did not vary much with time of night, we made no attempt to model explicitly sex differences in the diel periodicity of SBW flight.
We used all of these datasets to estimate Δ0 and Δs from the time at which the first (t0) and the median (50%; tc) migrating moths were observed on each observation night. The value of Δf (duration of the diel liftoff period) was determined from the time of first (t0) and last (tm) emigration. The values of Δs were related by regression analysis to T19h30 the ambient air temperature (°C) at 19h30 local DST, prior to the earliest observed emigration flights:
Observed values of Δ0 were related by regression analysis to the observed values of Δs:
The observed duration of the flight period (Δf in hours from first to last moth seen emigrating) was related to the observed values of Δ0. Given pronounced heteroscedasticity in these observations, we expressed this relationship as
However, because cooling evening temperatures have as a consequence the apparent reduction of the flight period, we used an optimization algorithm to estimate the value of a sixth parameter kf that, by multiplying with the value of p5, maximized the R2 between observed and simulated proportions of moths taking flight (with a precision of ±5%). The potential duration of the flight period is then given by
2.3. Simulations
To obtain the simulated proportions of moths taking flight we simulated 10,000 individual SBW moths (50% female) using the method described in [2], with mean and variances of wing surface areas and weights corresponding to the moths caught in canopy traps above a forest stand near Lac des Huit-Milles near Amqui, QC, during the 1989 and 1990 SBW flight seasons. Equation (7) was used to determine the proportion of individuals that could potentially lift off at a given time, and then Equations (8) and (9) were used to determine which of those moths could actually lift off given the ambient temperature and their individual mass and wing area. The proportion and sex ratio of moths taking off and the proportion of total fecundity carried away by emigrating females (Equation (10)) were compiled at intervals of 15 min.
3. Results
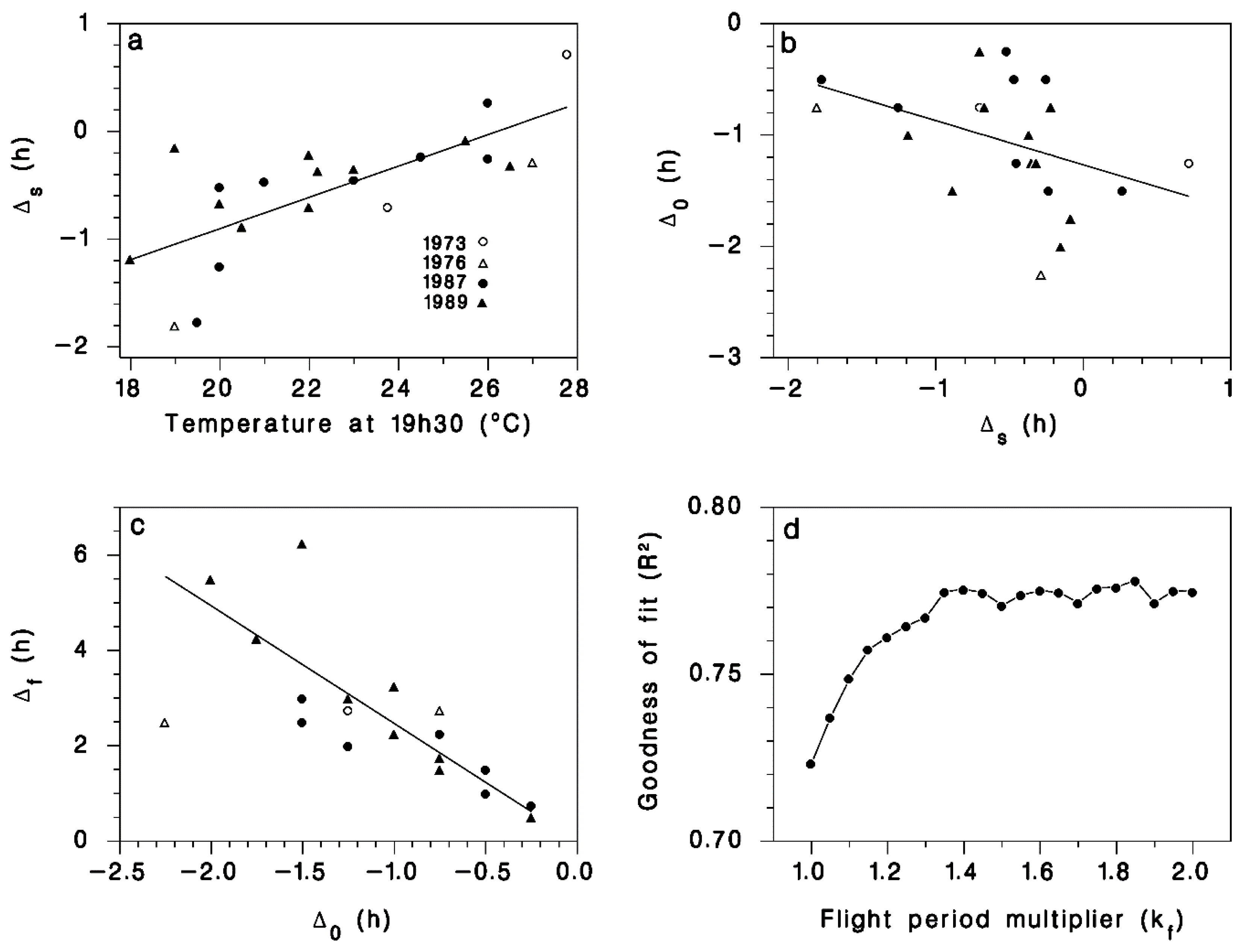
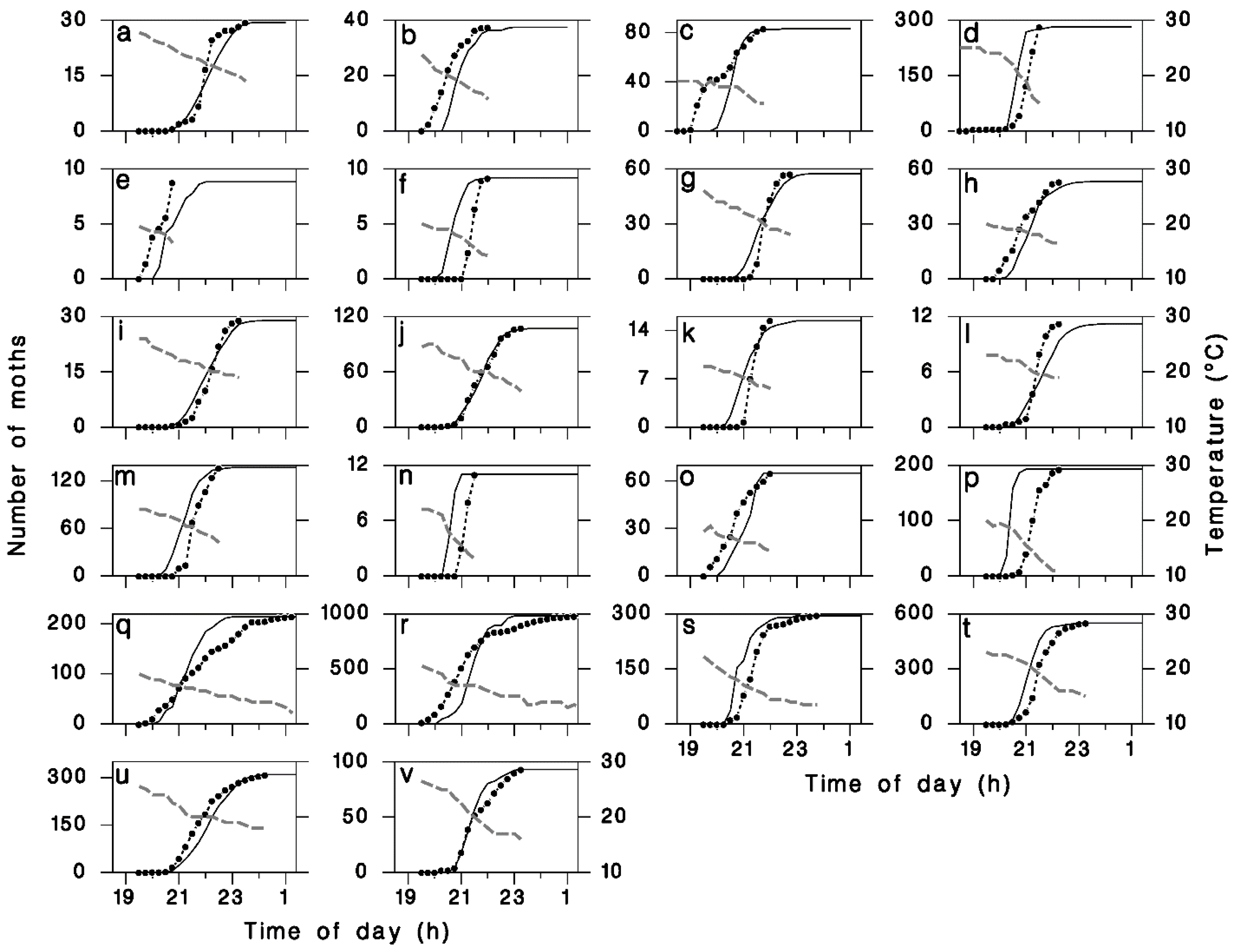
We found a significant linear relationship between T19h30 and the median timing of moth emigration Δs (Figure 2a) as described by Equation (11). There was also a significant negative relationship between the timing of the onset of emigration Δ0 and the value of Δs (Figure 2b), as described by Equation (12). The average value of parameter p5 in Equation (13) provided a good description of the relationship between observed flight duration Δf and the time Δ0 between first flight and peak flight (Figure 2c). Using the calibrated values for model parameters p1 through p5 (Table 1), the goodness-of-fit (R2) between observed and simulated cumulative liftoff patterns improved from 0.72 to an asymptotic maximum of 0.775 using kf = 1.35 (Figure 2d) in Equation (14). The resulting simulated patterns of liftoff timing corresponded well with field observations (Figure 3).
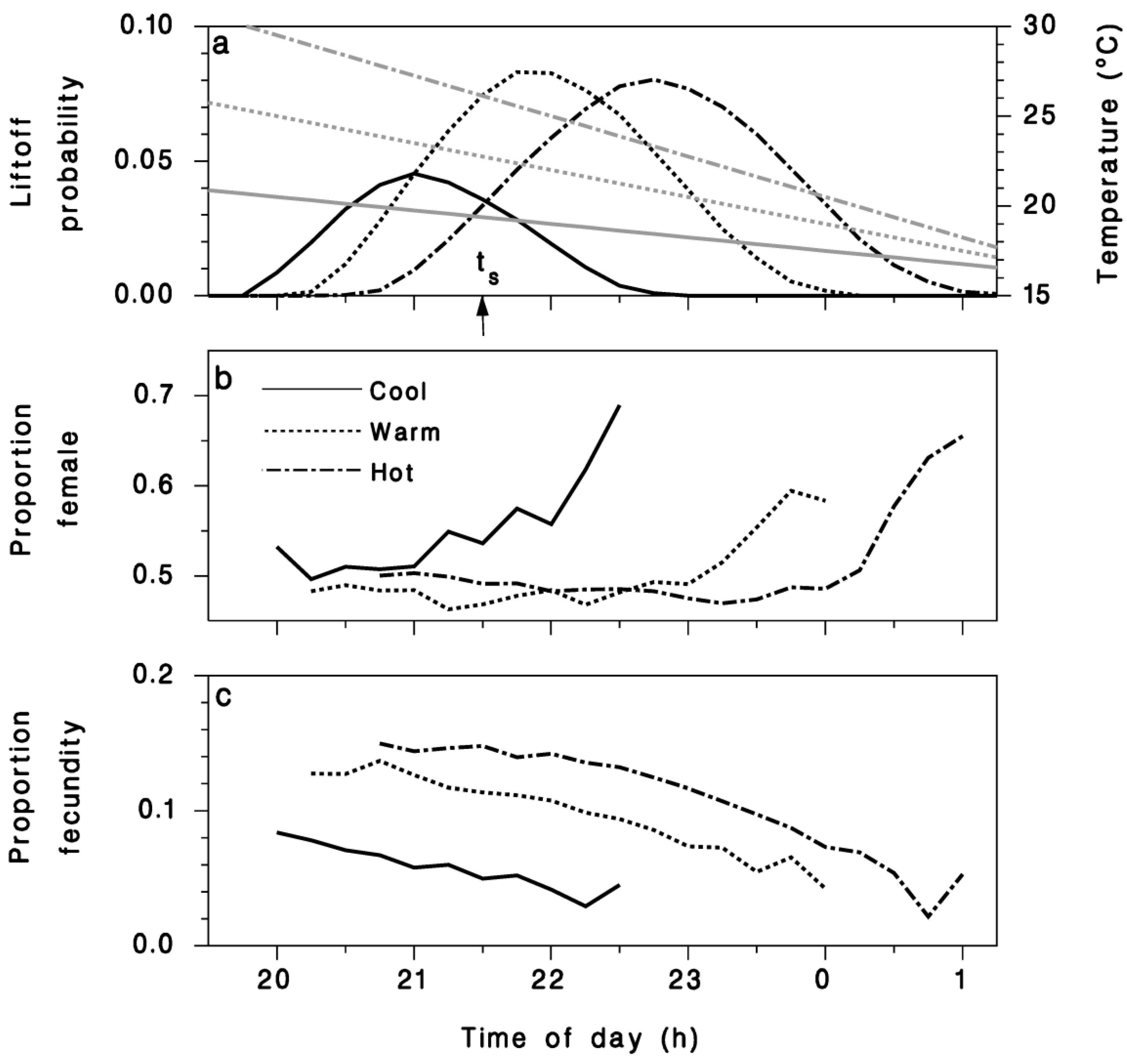
The circadian pattern of migration flight is strongly affected by temperature in two ways. As ambient temperature increases, the entire curve illustrated in Figure 1 shifts later relative to sunset and increases in overall duration (Figure 4a). Based on this model, the likelihood of crepuscular flight should peak nearly 0.5 h before sunset for a relatively cool evening (e.g., T19h30 = 20 °C), nearly 0.5 h after sunset for a warmer evening (e.g., T19h30 = 25 °C), and even later, nearly 1.5 h after sunset, for a relatively hot evening (e.g., T19h30 = 30 °C). However, the relationships between the wingbeat frequency and temperature, body weight, and wing area also determine how many moths can lift off at any given time. The resulting migration pattern, and its amplitude, are the composite of these effects (Figure 4a). Interestingly, as a result of the liftoff wingbeat relationship (Equations (8) and (9)), the sex ratio (proportion females) of emigrating moths varies with temperature and time during the evening (Figure 4b). Females make up the larger portion of migrants on cooler evenings, and later in the evening, contradicting the observation reported by Greenbank et al. [10] that the sex ratio of moths caught during migration liftoff did not vary much with time of night. The fecundity carried away by emigrating females also varies as a function of temperature and time through the evening (Figure 4c). Heavier females (i.e., those with a greater proportion of their remaining fecundity) can only lift off at warmer temperatures and thus typically earlier in the evening, while lighter females emigrating in cooler air at the end of the evening tend to have lower gravidity.
4. Discussion
The interaction of a crepuscular circadian rhythm of activity around sunset and the temporal variation of ambient temperature determines the diel pattern of emigration flight in adult SBW moths. These moths are believed capable of flight at temperatures between 14 and 30 °C [2,10,34]. Below that range, the moths are lethargic and cannot fan their wings fast enough to lift their body weight in flight. Above that range, the moths probably enter a state of torpor that protects them against heat damage [34]. As with circadian rhythms in many other species, temperature also affects the onset and duration of diel activity [13,14]. On cool evenings, the diel pattern of emigration starts and peaks early relative to sunset, and on warm evenings the pattern is shifted to later hours. This dependence of activity on temperature is likely adaptive, allowing migration to occur under a wider variety of atmospheric conditions. It may also ensure that emigration is synchronous for the moths in a given area, serving to keep the migrants together. Such mass flights are important to the population dynamics of this insect species: immigrants, when numerous enough, can produce progeny in numbers that overwhelm the various natural enemies keeping populations at endemic levels during the subsequent generation [35]. In addition, the migrating moths that begin their flights later on warm nights may still have sufficient time to travel long distances to new host areas, especially with favorable wind support, without running the risk of overly long flight periods and exhausted energy reserves. It is generally believed that migratory flights of SBW do not continue much past sunrise [10], although this assumption needs empirical support.
The total number of moths that emigrate from a given location, and the number of eggs that females can carry away by migration, are both strongly affected by evening temperatures on nights favorable to flight activity. Coherent wind patterns in the atmospheric boundary layer can develop from various phenomena under both stormy and clear conditions, but to our knowledge the meteorological support for insect migration events on otherwise clear nights has not been examined in prior studies. The evening transition from the turbulent daytime atmospheric boundary layer to the often-stable nighttime boundary layer over land, starting generally 1–2 h before sunset, is a subject of particular interest to both scientific and engineering communities [36,37]. The structure of the stable, stratified boundary layer has been examined in extensive detail [38]. However, describing the evening transition near sunset to that stable state remains a complex task, primarily because both the spatially heterogeneous land surface and the free atmosphere 1–2 km above the surface can exert complex dynamic influences on the lowest several hundred meters of the atmosphere. Detailed work [39,40,41,42,43] has described a three-stage evening transition of the boundary layer from convective (daytime) to stratified (nighttime) conditions, generally through the development of an inversion layer (where temperature increases with height) that grows upward from the surface after sunset. As the growth of the stable boundary layer eventually ceases in the hours after sunset, a coherent layer of high wind speeds known as the low-level jet often develops at the top of the temperature inversion in association with both the large-scale synoptic pattern and surface physiography (e.g., coastlines, mountain barriers, land cover heterogeneity) [37,44].
As the surface cools quickly in post-sunset hours, the migrating SBW moths ascend above the forest canopy and eventually concentrate [2,9,10] at the top of the inversion layer with higher temperatures (favorable to higher moth wingbeat frequencies) and stronger winds. Shear-based turbulence beneath the low-level jet may intermittently disrupt the upper levels of the stable layer well into the night [45,46,47]. Researchers have identified that turbulence associated with areas and layers of strong winds can serve as an indicator that helps orient flying insects in a common downwind direction [48]. In the absence of additional disruption by an overnight storm or frontal passage, the stable nocturnal boundary layer generally persists until its turbulent erosion shortly after sunrise the next morning. Insect crepuscular activity has evolved to take advantage of these dynamic conditions in the sunset transition of the atmospheric boundary layer for nocturnal migration flights. Similar patterns of behavior have been observed and studied in various species, and under similar weather conditions, on six continents around the world: Africa [49], Australia [26,27,50], East Asia [51,52], Europe [28,29,53,54,55,56], South America [57], and North America [8,10,31,32,33].
5. Conclusions
Our analysis of evening crepuscular flight activity in SBW moths is an essential component of our larger model of migration activity, deepening our understanding of spatiotemporal patterns and processes in the course of SBW population fluctuations and outbreak events. It is in the evening development and overnight persistence of the stable nocturnal boundary layer, amid otherwise calm synoptic conditions, that we find strong support for coherent, long-distance SBW migration events resulting from the temporal alignment of crepuscular circadian activity and favorable boundary layer temperature and wind conditions above the forest canopy. Where a nocturnal low-level jet develops above the boundary layer, greater shear turbulence is produced just below that jet in the warm inversion, with those combined conditions favorable to coherent long-distance migratory flight for large numbers of insects. We are thus interested in continuing our examination of the evening crepuscular conditions favorable to liftoff from the forest canopy that can result in mass migration events. We also intend to focus on the dynamic conditions in the evening transition and nocturnal boundary layer above the forest canopy that lead to the coherent layering [26,28,29,50,53,54,55,58], orientation [9,10,58,59,60], and long-distance transport [27,61] of migrating insects.
Climate change may affect the circadian rhythm of SBW emigration. However, the coarse representations of the atmospheric boundary layer provided by current climate models [62] do not allow us to predict those effects with any accuracy using the emigration model described here. The most obvious evidence of changing climate has been found in an increase of daily minimum temperatures [63], with less pronounced increases of daily maximum temperatures. In our model, SBW moths attempt to optimize their liftoff time with respect to evening temperature to remain in the warmer air that supports the wingbeat of heavier moths before the cooler nocturnal boundary layer is established and flight activity is curtailed. All else remaining constant, increasing nighttime minimum temperature allows for slower evening cooling and a corollary lengthening of the migration flight period. Additional data of the type used in the present analysis may show such a change over the past several decades and in the future. Warmer nighttime temperatures may also destabilize the nocturnal boundary layer to some extent [64]. An overall warmer and well-mixed nocturnal boundary layer would allow SBW moths to fly at lower altitudes, which would otherwise remain too cold for flight activity. A less stable nocturnal boundary layer implies greater turbulence, which can bolster some moths flying in updrafts while driving others toward the surface in downdrafts. Finally, diminished nocturnal boundary layer stability would likely result in a more diffuse upper boundary of the near-surface inversion layer, which may not provide sufficient thermal gradients for the formation of the low-level jet. Without the aid of high wind speeds in and near that nocturnal low-level jet, far shorter SBW flight distances could be expected in mass migration events. We look forward to the opportunity to examine such changes in boundary layer dynamics using detailed numerical models (e.g., [65]) and the potential effects of changing evening transition and nocturnal boundary-layer conditions on our insect flight modeling efforts.
Author Contributions
J.R. developed the model and parameter estimates; M.G. helped with parameter estimation, literature review and writing; R.S.-A. helped with programming and writing. Conceptualization, J.R. and R.S.-A.; Methodology, J.R.; Software, R.S.-A.; Validation, J.R., M.G. and R.S.-A.; Formal Analysis, J.R.; Investigation, J.R.; Resources, J.R.; Data Curation, J.R.; Writing—Original Draft Preparation, J.R. and M.G.; Writing—Review & Editing, J.R., M.G. and R.S.-A.; Visualization, J.R.; Funding Acquisition, J.R.
Funding
This research was funded by the natural resources and forest departments of Newfoundland, Nova Scotia, New Brunswick, Quebec, Ontario, Manitoba, Saskatchewan, Alberta and British Columbia. Funds were also provided by the Atlantic Canada Opportunities Agency, and under the US/Canada Forest Health & Innovation Summit Initiative sponsored by the USDA Forest Service, Canadian Forest Service of Natural Resources Canada, and the USA Endowment for Forestry and Communities, with cooperation by SERG-International.
Acknowledgments
The authors acknowledge Brian R. Sturtevant for his helpful comments on the manuscript. M.G. wishes to acknowledge the support of Philip A. Townsend at the University of Wisconsin–Madison.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dingle, H. Migration: The Biology of Life on the Move, 2nd ed.; Oxford University Press: New York, NY, USA, 2014; 326p. [Google Scholar] [CrossRef]
- Régnière, J.; Delisle, J.; Sturtevant, B.R.; Garcia, M.; St-Amant, R. Modeling migratory flight in the spruce budworm: Temperature constraints. Forests 2019, 10, 802. [Google Scholar] [CrossRef]
- Hu, G.; Lim, K.S.; Reynolds, D.R.; Reynolds, A.M.; Chapman, J.W. Wind-related orientation patterns in diurnal, crepuscular and nocturnal high-altitude insect migrants. Front. Behav. Neurosci. 2016, 10, 32. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, A.Y.; Plotkin, D.; Hamilton, C.A.; Gough, H.; St Laurent, R.; Owens, H.L.; Homziak, N.T.; Barber, J.R. Diel behavior in moths and butterflies: A synthesis of data illuminates the evolution of temporal activity. Org. Divers. Evol. 2018, 18, 13–27. [Google Scholar] [CrossRef]
- Rund, S.; O’Donnell, A.; Gentile, J.; Reece, S. Daily rhythms in mosquitoes and their consequences for malaria transmission. Insects 2016, 7, 14. [Google Scholar] [CrossRef]
- Broadhead, G.T.; Basu, T.; von Arx, M.; Raguso, R.A. Diel rhythms and sex differences in the locomotor activity of hawkmoths. J. Exp. Biol. 2017, 220, 1472–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wellington, W.G. The light reactions of the spruce budworm, Choristoneura fumiferana Clemens (Lepidoptera: Tortricidae). Can. Entomol. 1948, 80, 56–82. [Google Scholar] [CrossRef]
- Henson, W.R. Mass flights of the spruce budworm. Can. Entomol. 1951, 83, 240. [Google Scholar] [CrossRef]
- Schaefer, G.W. Radar observations of insect flight. Symp. R. Entomol. Soc. Lond. 1976, 7, 157–197. [Google Scholar]
- Greenbank, D.O.; Schaefer, G.W.; Rainey, R.C. Spruce budworm (Lepidoptera: Tortricidae) moth flight and dispersal: New understanding from canopy observations, radar and aircraft. Mem. Entomol. Soc. Can. 1980, 112, 1–49. [Google Scholar] [CrossRef]
- Kipp, L.R.; Lonergan, G.C.; Bell, W.J. Male periodicity and the timing of mating in the spruce budworm (Lepidoptera: Tortricidae): Influence of population density and temperature. Environ. Entomol. 1995, 24, 1150–1159. [Google Scholar] [CrossRef]
- Cardé, R.T.; Roelofs, W.L. Temperature modification of male sex pheromone response and factors affecting female calling in Holomelina immaculata (Lepidoptera: Arctiidae). Can. Entomol. 1973, 105, 1505–1512. [Google Scholar] [CrossRef]
- Cardé, R.T.; Comeau, A.; Baker, T.C.; Roelofs, W.L. Moth mating periodicity: Temperature regulates the circadian gate. Experientia 1975, 31, 46–48. [Google Scholar] [CrossRef] [PubMed]
- Comeau, A.; Cardé, R.T.; Roelofs, W.L. Relationship of ambient temperatures to diel periodicities of sex attraction in six species of Lepidoptera. Can. Entomol. 1976, 108, 415–418. [Google Scholar] [CrossRef]
- Alerstam, T.; Chapman, J.W.; Bäckman, J.; Smith, A.D.; Karlsson, H.; Nilsson, C.; Reynolds, D.R.; Klaassen, H.G.; Hill, J.K. Convergent patterns of long-distance nocturnal migration in noctuid moths and passerine birds. Proc. R. Soc. B Biol. Sci. 2011, 278, 3074–3080. [Google Scholar] [CrossRef]
- Krauel, J.J.; Westbrook, J.K.; McCracken, G.F. Weather-driven dynamics in a dual-migrant system: Moths and bats. J. Anim. Ecol. 2015, 84, 604–614. [Google Scholar] [CrossRef]
- Krauel, J.J.; Brown, V.A.; Westbrook, J.K.; McCracken, G.F. Predator–prey interaction reveals local effects of high-altitude insect migration. Oecologia 2018, 186, 49–58. [Google Scholar] [CrossRef]
- Sanders, C.J. Daily activity patterns and sex pheromone specificity as sexual isolating mechanisms in two species of Choristoneura (Lepidoptera: Tortricidae). Can. Entomol. 1971, 103, 498–502. [Google Scholar] [CrossRef]
- Sanders, C.J.; Lucuik, G.S. Effects of photoperiod and size on flight activity and oviposition in the eastern spruce budworm (Lepidoptera: Tortricidae). Can. Entomol. 1975, 107, 1289–1299. [Google Scholar] [CrossRef]
- Simmons, G.A.; Chen, C.W. Application of harmonic analysis and polynomial regression to study flight activity of Choristoneura fumiferana (Clem.) (Lepidoptera: Tortricidae) in the field. Abstracts, Forty-Seventh Annual Meeting, Eastern Branch Entomological Society of America. J. N. Y. Entomol. Soc. 1975, 83, 266. [Google Scholar]
- Régnière, J.; Powell, J.; Bentz, B.; Nealis, V. Effects of temperature on development, survival and reproduction of insects: Experimental design, data analysis and modeling. J. Insect Physiol. 2012, 58, 634–647. [Google Scholar] [CrossRef]
- Wellington, W.G.; Henson, W.R. Notes on the effects of physical factors on the spruce budworm, Choristoneura fumiferana (Clem.). Can. Entomol. 1947, 79, 168–170. [Google Scholar] [CrossRef]
- Dickison, R.B.B.; Haggis, M.J.; Rainey, R.C. Spruce budworm moth flight and storms: Case study of a cold front system. J. Clim. Appl. Meteorol. 1983, 22, 278–286. [Google Scholar] [CrossRef]
- Dickison, R.B.B.; Haggis, M.J.; Rainey, R.C.; Burns, L.M.D. Spruce budworm moth flight and storms: Further studies using aircraft and radar. J. Clim. Appl. Meteorol. 1986, 25, 1600–1608. [Google Scholar] [CrossRef]
- Pedgley, D.E.; Reynolds, D.R.; Riley, J.R.; Tucker, M.R. Flying insects reveal small-scale wind systems. Weather 1982, 37, 295–306. [Google Scholar] [CrossRef]
- Drake, V.A. The vertical distribution of macro-insects migrating in the nocturnal boundary layer: A radar study. Bound. Layer Meteorol. 1984, 28, 353–374. [Google Scholar] [CrossRef]
- Drake, V.A. Radar observations of moths migrating in a nocturnal low-level jet. Ecol. Entomol. 1985, 10, 259–265. [Google Scholar] [CrossRef]
- Reynolds, D.R.; Chapman, J.W.; Edwards, A.S.; Smith, A.D.; Wood, C.R.; Barlow, J.F.; Woiwod, I.P. Radar studies of the vertical distribution of insects migrating over southern Britain: The influence of temperature inversions on nocturnal layer concentrations. Bull. Entomol. Res. 2005, 95, 259–274. [Google Scholar] [CrossRef]
- Reynolds, D.R.; Smith, A.D.; Chapman, J.W. A radar study of emigratory flight and layer formation by insects at dawn over southern Britain. Bull. Entomol. Res. 2008, 98, 35–52. [Google Scholar] [CrossRef] [PubMed]
- Chapman, J.W.; Drake, V.A.; Reynolds, D.R. Recent insights from radar studies of insect flight. Ann. Rev. Entomol. 2011, 56, 337–356. [Google Scholar] [CrossRef]
- Westbrook, J.K.; Eyster, R.S.; Wolf, W.W. WSR-88D doppler radar detection of corn earworm moth migration. Int. J. Biometeorol. 2014, 58, 931–940. [Google Scholar] [CrossRef]
- Boulanger, Y.; Fabry, F.; Kilambi, A.; Pureswaran, D.S.; Sturtevant, B.R.; Saint-Amant, R. The use of weather surveillance radar and high-resolution three-dimensional weather data to monitor a spruce budworm mass exodus flight. Agric. For. Meteorol. 2017, 234, 127–135. [Google Scholar] [CrossRef]
- Westbrook, J.K.; Eyster, R.S. Doppler weather radar detects emigratory flights of noctuids during a major pest outbreak. Remote Sens. Appl. Soc. Environ. 2017, 8, 64–70. [Google Scholar] [CrossRef]
- Sanders, J.C.; Wallace, D.R.; Luicuik, G.S. Flight activity of female eastern spruce budworm (Lepidoptera: Tortricidae) at constant temperatures in the laboratory. Can. Entomol. 1978, 110, 627–632. [Google Scholar] [CrossRef]
- Régnière, J.; Cooke, B.; Béchard, A.; Dupont, A.; Therrien, P. Dynamics and management of rising outbreak spruce budworm populations. Forests 2019, 10, 748. [Google Scholar] [CrossRef]
- Baklanov, A.A.; Grisogono, B.; Bornstein, R.; Zilitinkevich, S.S.; Taylor, P.; Larsen, S.E.; Rotach, M.W.; Fernando, H.J.S. The nature, theory, and modeling of atmospheric planetary boundary layers. Bull. Am. Meteorol. Soc. 2011, 92, 123–128. [Google Scholar] [CrossRef]
- Angevine, W.M. Transitional, entraining, cloudy, and coastal boundary layers. Acta Geophysica 2008, 56, 2–20. [Google Scholar] [CrossRef]
- Mahrt, L. Stably stratified atmospheric boundary layers. Ann. Rev. Fluid Mech. 2014, 46, 23–45. [Google Scholar] [CrossRef]
- Mahrt, L. The early evening boundary layer transition. Quart. J. R. Meteorol. Soc. 1981, 107, 329–343. [Google Scholar] [CrossRef]
- Mahrt, L. Nocturnal boundary-layer regimes. Bound. Layer Meteorol. 1998, 88, 255–278. [Google Scholar] [CrossRef]
- Mahrt, L. The near-surface evening transition. Quart. J. R. Meteorol. Soc. 2017, 143, 2940–2948. [Google Scholar] [CrossRef]
- Acevedo, O.C.; Fitzjarrald, D.R. The early evening surface-layer transition: Temporal and spatial variability. J. Atmos. Sci. 2001, 58, 2650–2667. [Google Scholar] [CrossRef]
- Sastre, M.; Yagüe, C.; Román-Cascón, C.; Maqueda, G. Atmospheric boundary-layer evening transitions: A comparison between two different experimental sites. Bound. Layer Meteorol. 2015, 157, 375–399. [Google Scholar] [CrossRef]
- Angevine, W.M.; Tjernström, M.; Žagar, M. Modeling of the coastal boundary layer and pollutant transport in New England. J. Appl. Meteorol. Climatol. 2006, 45, 137–154. [Google Scholar] [CrossRef]
- Nieuwstadt, F.T.M. The turbulent structure of the stable, nocturnal boundary layer. J. Atmos. Sci. 1984, 41, 2202–2216. [Google Scholar] [CrossRef]
- Acevedo, O.C.; Mahrt, L.; Puhales, F.S.; Costa, F.D.; Medeiros, L.E.; Degrazia, G.A. Contrasting structures between the decoupled and coupled states of the stable boundary layer. Quart. J. R. Meteorol. Soc. 2016, 142, 693–702. [Google Scholar] [CrossRef]
- Mahrt, L. Microfronts in the nocturnal boundary layer. Quart. J. R. Meteorol. Soc. 2019, 145, 546–562. [Google Scholar] [CrossRef]
- Reynolds, A.M.; Reynolds, D.R.; Smith, A.D.; Chapman, J.W. Orientation cues for high-flying nocturnal insect migrants: Do turbulence-induced temperature and velocity fluctuations indicate the mean wind flow? PLoS ONE 2010, 5, e15758. [Google Scholar] [CrossRef]
- Riley, J.R.; Reynolds, D.R.; Rainey, R.C. Radar-based studies of the migratory flight of grasshoppers in the middle Niger area of Mali. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1979, 204, 67–82. [Google Scholar] [CrossRef]
- Rennie, S.J. Common orientation and layering of migrating insects in southeastern Australia observed with a Doppler weather radar. Meteorol. Appl. 2014, 21, 218–229. [Google Scholar] [CrossRef]
- Feng, H.; Wu, X.; Wu, B.; Wu, K. Seasonal migration of Helicoverpa armigera (Lepidoptera: Noctuidae) over the Bohai Sea. J. Econ. Entomol. 2009, 102, 95–104. [Google Scholar] [CrossRef]
- Fu, X.; Zhao, X.; Xie, B.; Ali, A.; Wu, K. Seasonal pattern of Spodoptera litura (Lepidoptera: Noctuidae) migration across the Bohai Strait in northern China. J. Econ. Entomol. 2015, 108, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Wood, C.R.; Chapman, J.W.; Reynolds, D.R.; Barlow, J.F.; Smith, A.D.; Woiwod, I.P. The influence of the atmospheric boundary layer on nocturnal layers of noctuids and other moths migrating over southern Britain. Int. J. Biometeorol. 2006, 50, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Wood, C.R.; Reynolds, D.R.; Wells, P.M.; Barlow, J.F.; Woiwod, I.P.; Chapman, J.W. Flight periodicity and the vertical distribution of high-altitude moth migration over southern Britain. Bull. Entomol. Res. 2009, 99, 525–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, C.R.; Clark, S.J.; Barlow, J.F.; Chapman, J.W. Layers of nocturnal insect migrants at high-altitude: The influence of atmospheric conditions on their formation. Agric. For. Entomol. 2010, 12, 113–121. [Google Scholar] [CrossRef]
- Dreyer, D.; El Jundi, B.; Kishkinev, D.; Suchentrunk, C.; Campostrini, L.; Frost, B.J.; Zechmeister, T.; Warrant, E.J. Evidence for a southward autumn migration of nocturnal noctuid moths in central Europe. J. Exp. Biol. 2018, 221, 179218. [Google Scholar] [CrossRef]
- Wang, H.-H.; Grant, W.E.; Elliott, N.C.; Brewer, M.J.; Koralewski, T.E.; Westbrook, J.K.; Alves, T.M.; Sword, G.A. Integrated modelling of the life cycle and aeroecology of wind-borne pests in temporally-variable spatially-heterogeneous environment. Ecol. Model. 2019, 399, 23–38. [Google Scholar] [CrossRef]
- Reynolds, A.M.; Reynolds, D.R.; Smith, A.D.; Chapman, J.W. A single wind-mediated mechanism explains high-altitude “non-goal oriented” headings and layering of nocturnally migrating insects. Proc. R. Soc. Lond. B Biol. Sci. 2010, 277, 765–772. [Google Scholar] [CrossRef]
- Aralimarad, P.; Reynolds, A.M.; Lim, K.S.; Reynolds, D.R.; Chapman, J.W. Flight altitude selection increases orientation performance in high-flying nocturnal insect migrants. Anim. Behav. 2011, 82, 1221–1225. [Google Scholar] [CrossRef]
- Riley, J.R. Collective orientation in night-flying insects. Nature 1975, 253, 113. [Google Scholar] [CrossRef]
- Reynolds, A.M.; Reynolds, D.R.; Riley, J.R. Does a “turbophoretic” effect account for layer concentrations of insects migrating in the stable night-time atmosphere? J. R. Soc. Interface 2009, 6, 87–95. [Google Scholar] [CrossRef]
- Holtslag, A.A.M.; Svensson, G.; Baas, P.; Basu, S.; Beare, B.; Beljaars, A.C.M.; Bosveld, F.C.; Cuxart, J.; Lindvall, J.; Steeneveld, G.J.; et al. Stable atmospheric boundary layers and diurnal cycles: Challenges for weather and climate models. Bull. Am. Meteorol. Soc. 2013, 94, 1691–1706. [Google Scholar] [CrossRef]
- Davy, R. The climatology of the atmospheric boundary layer in contemporary global climate models. J. Clim. 2018, 31, 9151–9173. [Google Scholar] [CrossRef]
- McNider, R.T.; Steeneveld, G.J.; Holtslag, A.A.M.; Pielke, R.A.; Mackaro, S.; Pour-Biazar, A.; Walters, J.; Nair, U.; Christy, J. Response and sensitivity of the nocturnal boundary layer over land to added longwave radiative forcing. J. Geophys. Res. Atmos. 2012, 117, D14106. [Google Scholar] [CrossRef]
- Horvath, K.; Koracin, D.; Vellore, R.; Jiang, J.; Belu, R. Sub-kilometer dynamical downscaling of near-surface winds in complex terrain using WRF and MM5 mesoscale models. J. Geophys. Res. Atmos. 2012, 117, D11111. [Google Scholar] [CrossRef]
Figure 1.
Circadian model of crepuscular migration liftoff activity (Equation (2)) illustrating terms used to define τ in Equation (1), with ts: Time of sunset; Δs: Time between sunset ts and peak flight tc; Δ0: Time between first flight t0 and peak flight tc; Δf: Duration of the migration liftoff period; and tm: End of flight period.
Figure 1.
Circadian model of crepuscular migration liftoff activity (Equation (2)) illustrating terms used to define τ in Equation (1), with ts: Time of sunset; Δs: Time between sunset ts and peak flight tc; Δ0: Time between first flight t0 and peak flight tc; Δf: Duration of the migration liftoff period; and tm: End of flight period.
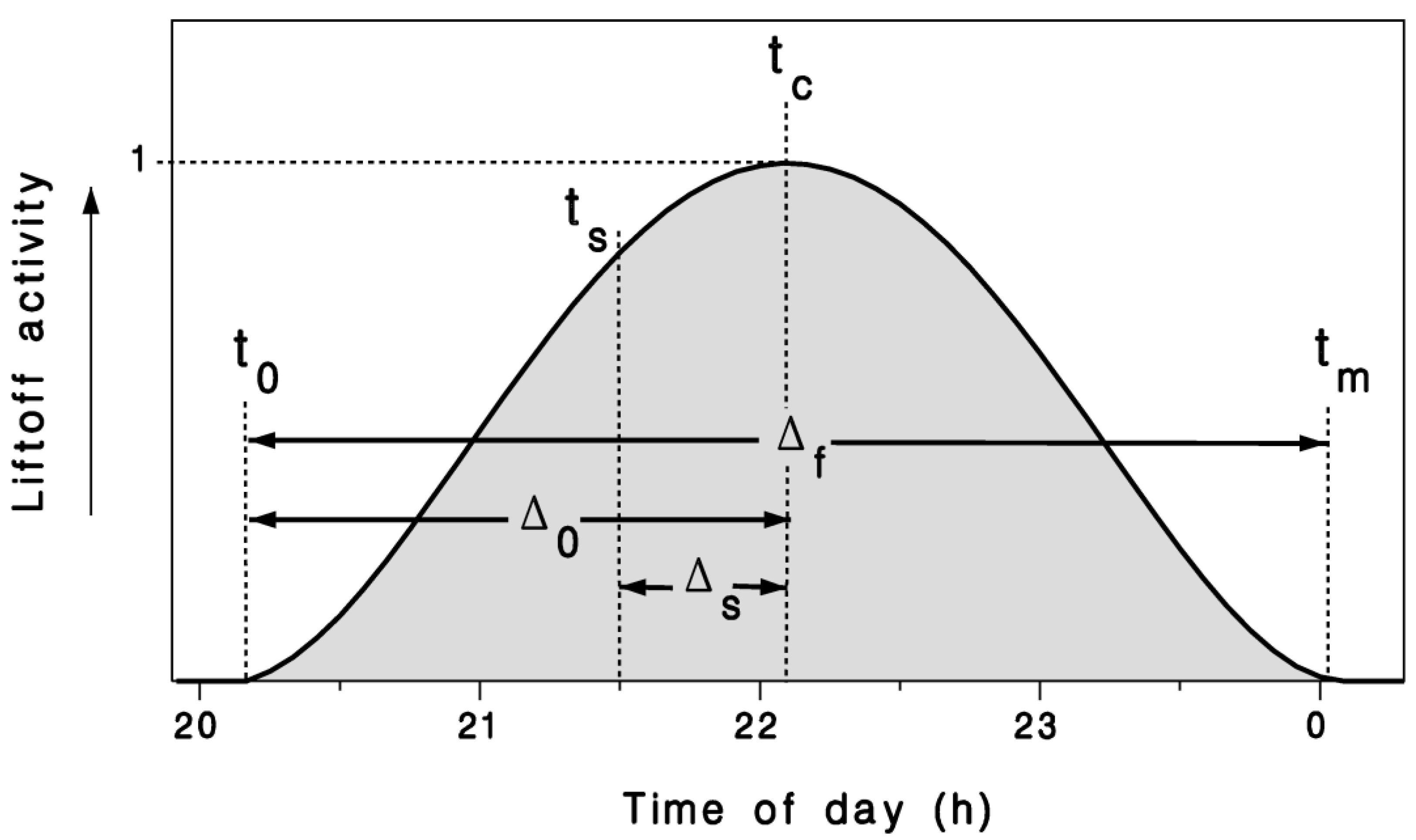
Figure 2.
(a) Relationship between T19h30 the temperature at 19h30 local DST and Δs the shift between the time at which 50% of migrating moths were observed and the time of sunset ts. Line is Equation (11). (b) Relationship between Δs and the shift between the first migrating moth and sunset Δ0. Line is Equation (12). (c) Relationship between Δ0 and the observed duration of the moth migration period Δf. Line is Equation (13). (d) Optimization (maximum R2) of the value of parameter kf in Equation (14). The value kf =1.35 yielded R2 = 0.775 between observed and simulated cumulative proportion of moths emigrating over 22 evenings (see Figure 3).
Figure 2.
(a) Relationship between T19h30 the temperature at 19h30 local DST and Δs the shift between the time at which 50% of migrating moths were observed and the time of sunset ts. Line is Equation (11). (b) Relationship between Δs and the shift between the first migrating moth and sunset Δ0. Line is Equation (12). (c) Relationship between Δ0 and the observed duration of the moth migration period Δf. Line is Equation (13). (d) Optimization (maximum R2) of the value of parameter kf in Equation (14). The value kf =1.35 yielded R2 = 0.775 between observed and simulated cumulative proportion of moths emigrating over 22 evenings (see Figure 3).
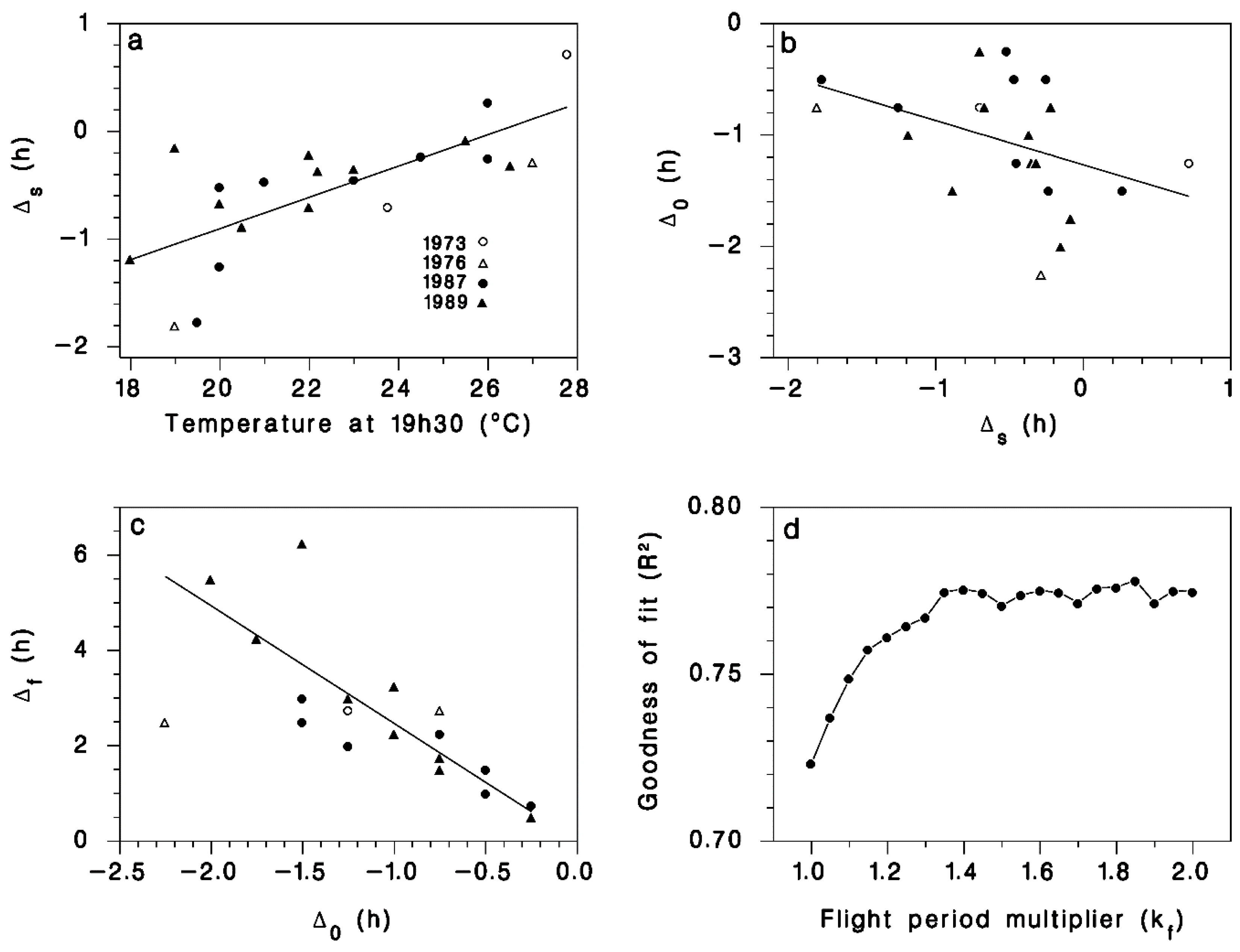
Figure 3.
Observed (closed circles with dotted lines) and simulated (solid lines) cumulative moths flights with observed evening temperatures (dashed grey lines). (a) and (b) 9 and 16 July 1973 using data from [19]. (c) and (d) 2 and 5 July 1976, from observations by C.J. Sanders near Sault Ste. Marie, Ontario. (e–v): 18 individual nights in July 1987 and 1989, from observations by C.J. Sanders near Black Sturgeon Lake, Ontario.
Figure 3.
Observed (closed circles with dotted lines) and simulated (solid lines) cumulative moths flights with observed evening temperatures (dashed grey lines). (a) and (b) 9 and 16 July 1973 using data from [19]. (c) and (d) 2 and 5 July 1976, from observations by C.J. Sanders near Sault Ste. Marie, Ontario. (e–v): 18 individual nights in July 1987 and 1989, from observations by C.J. Sanders near Black Sturgeon Lake, Ontario.
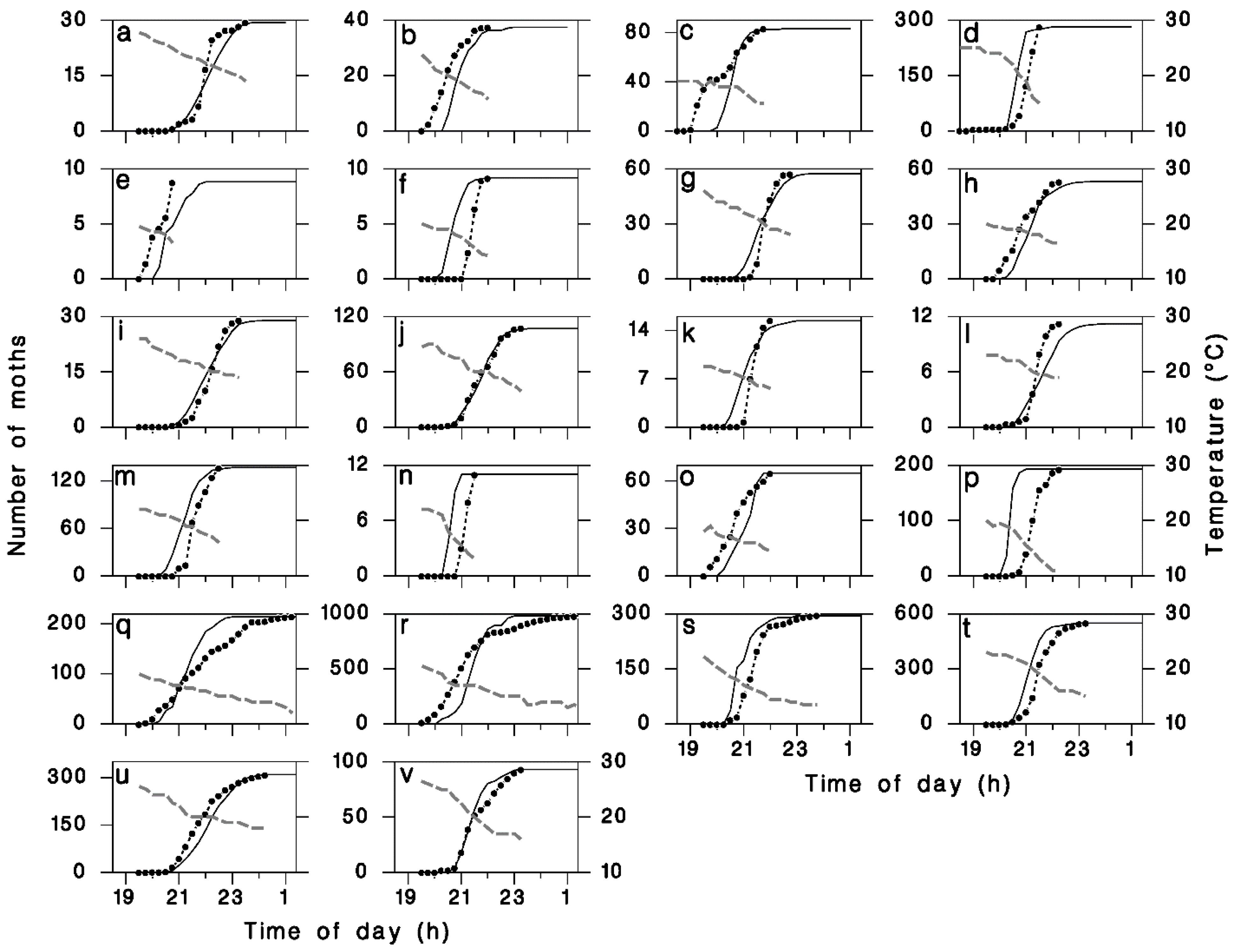
Figure 4.
(a) Proportion of moths (black curves) emigrating under three temperature regimes (gray lines) on cool (solid), warm (dotted), and hot (dash-dotted) nights. (b) Proportion female among the moths emigrating at each time step. (c) Average proportion of initial fecundity carried away by emigrating females (Equation (10)).
Figure 4.
(a) Proportion of moths (black curves) emigrating under three temperature regimes (gray lines) on cool (solid), warm (dotted), and hot (dash-dotted) nights. (b) Proportion female among the moths emigrating at each time step. (c) Average proportion of initial fecundity carried away by emigrating females (Equation (10)).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Model parameter calibration and regression results.
| Parameter | Calibration Value | Equation | Regression Statistics |
|---|---|---|---|
| p1 | −3.8 ± 0.7 h | (11) | F = 22.5; df = 1,20; R2 = 0.529; p < 0.001 |
| p2 | 0.145 ± 0.031 h/°C | ||
| p3 | −1.267 ± 0.146 h | (12) | F = 4.5; df = 1,20; R2 = 0.183; p < 0.047 |
| p4 | −0.397 ± 0.187 | ||
| p5 | −2.465 ± 0.152 | (13) | |
| kf | 1.35 ± 0.025 | (14) | R2 = 0.775 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Régnière, J.; Garcia, M.; Saint-Amant, R. Modeling Migratory Flight in the Spruce Budworm: Circadian Rhythm. Forests 2019, 10, 877. https://doi.org/10.3390/f10100877
AMA Style
Régnière J, Garcia M, Saint-Amant R. Modeling Migratory Flight in the Spruce Budworm: Circadian Rhythm. Forests. 2019; 10(10):877. https://doi.org/10.3390/f10100877
Chicago/Turabian StyleRégnière, Jacques, Matthew Garcia, and Rémi Saint-Amant. 2019. "Modeling Migratory Flight in the Spruce Budworm: Circadian Rhythm" Forests 10, no. 10: 877. https://doi.org/10.3390/f10100877
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.