Improving Mountain Pine Beetle Survival Predictions Using Multi-Year Temperatures Across the Western USA

Abstract
1. Introduction
2. Methods
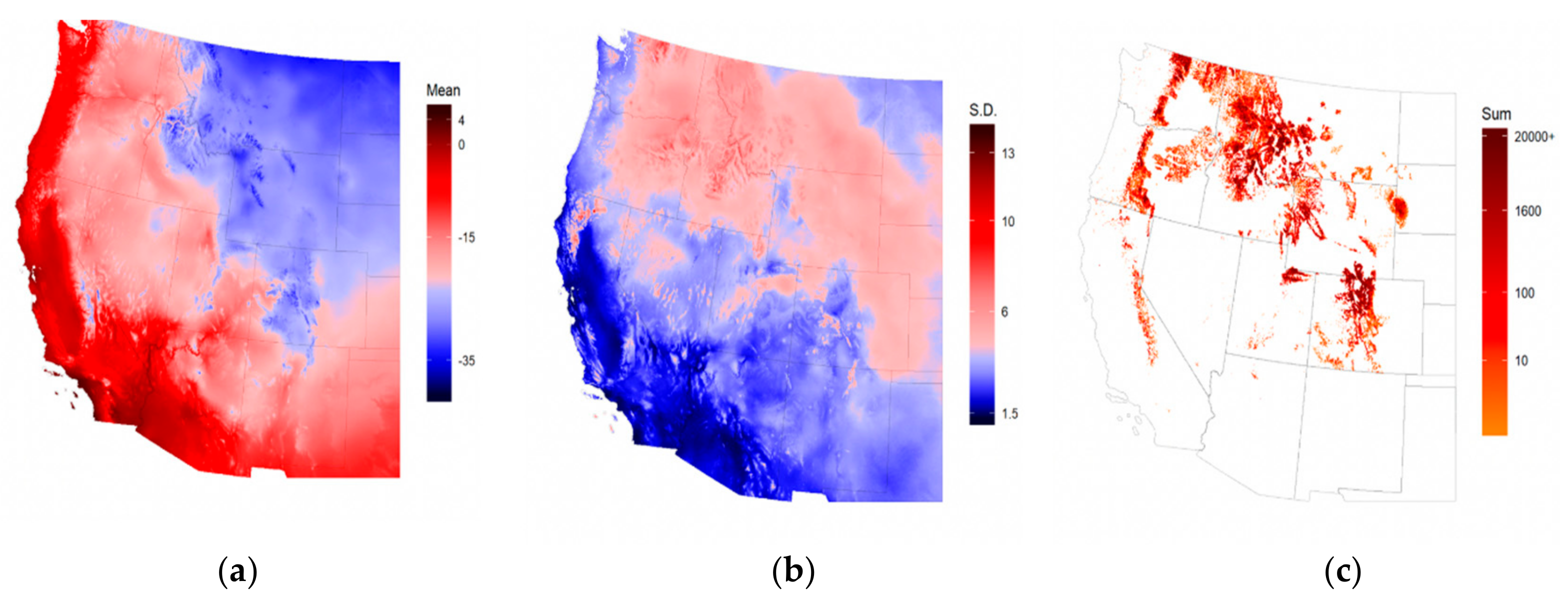
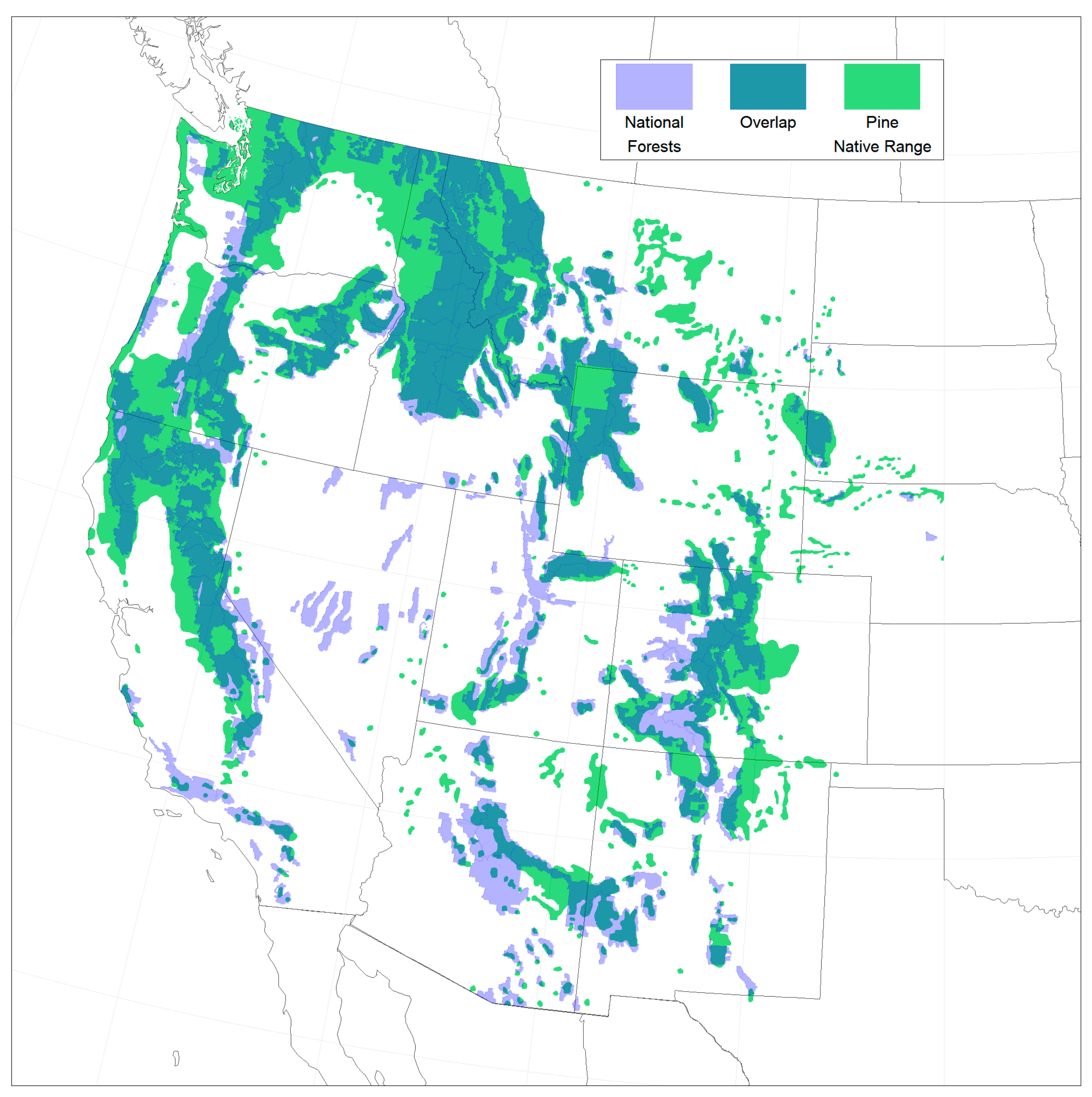
2.1. Study Area and Data
2.2. MPB Overwinter Survival Model
2.3. Analysis
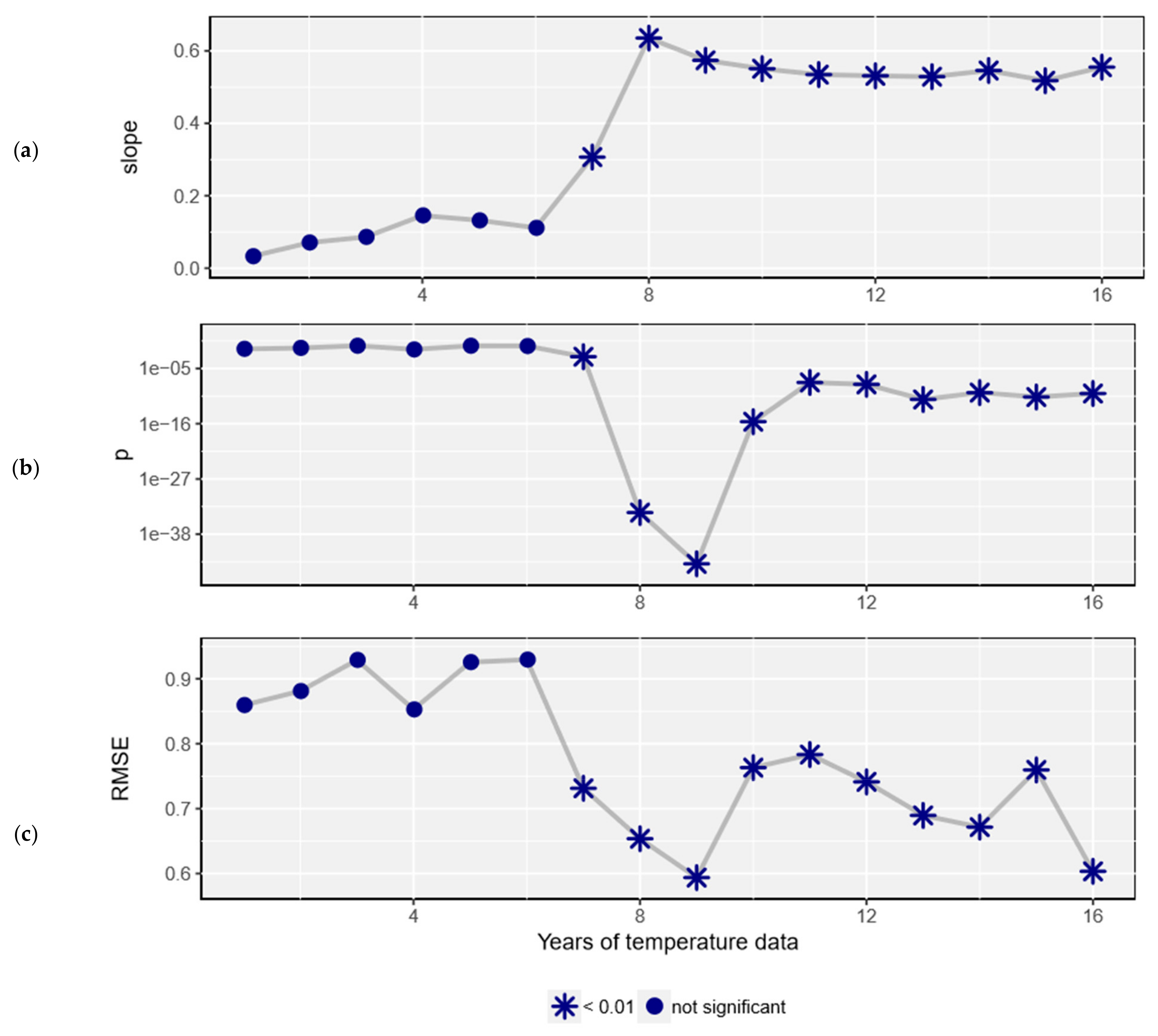
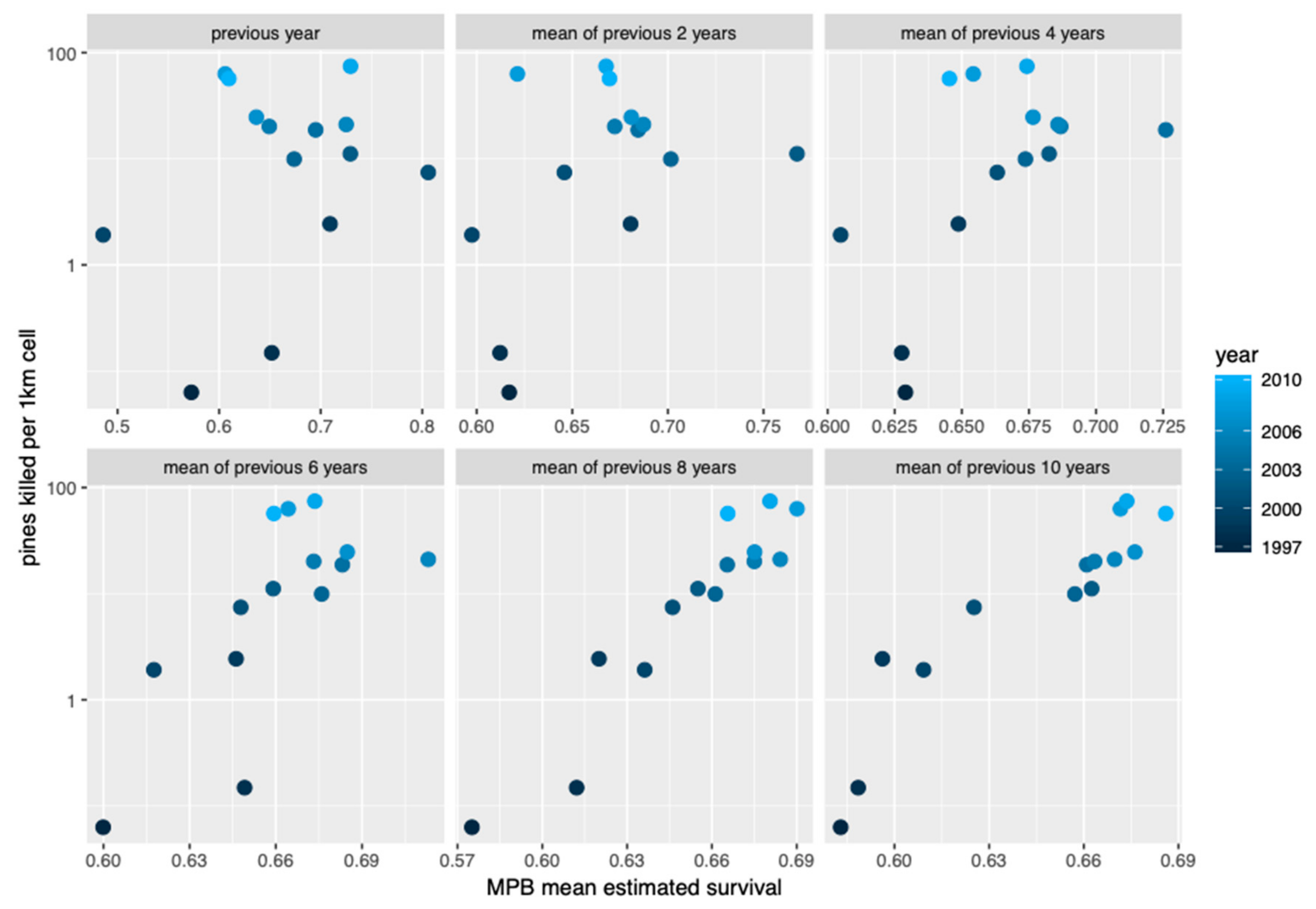
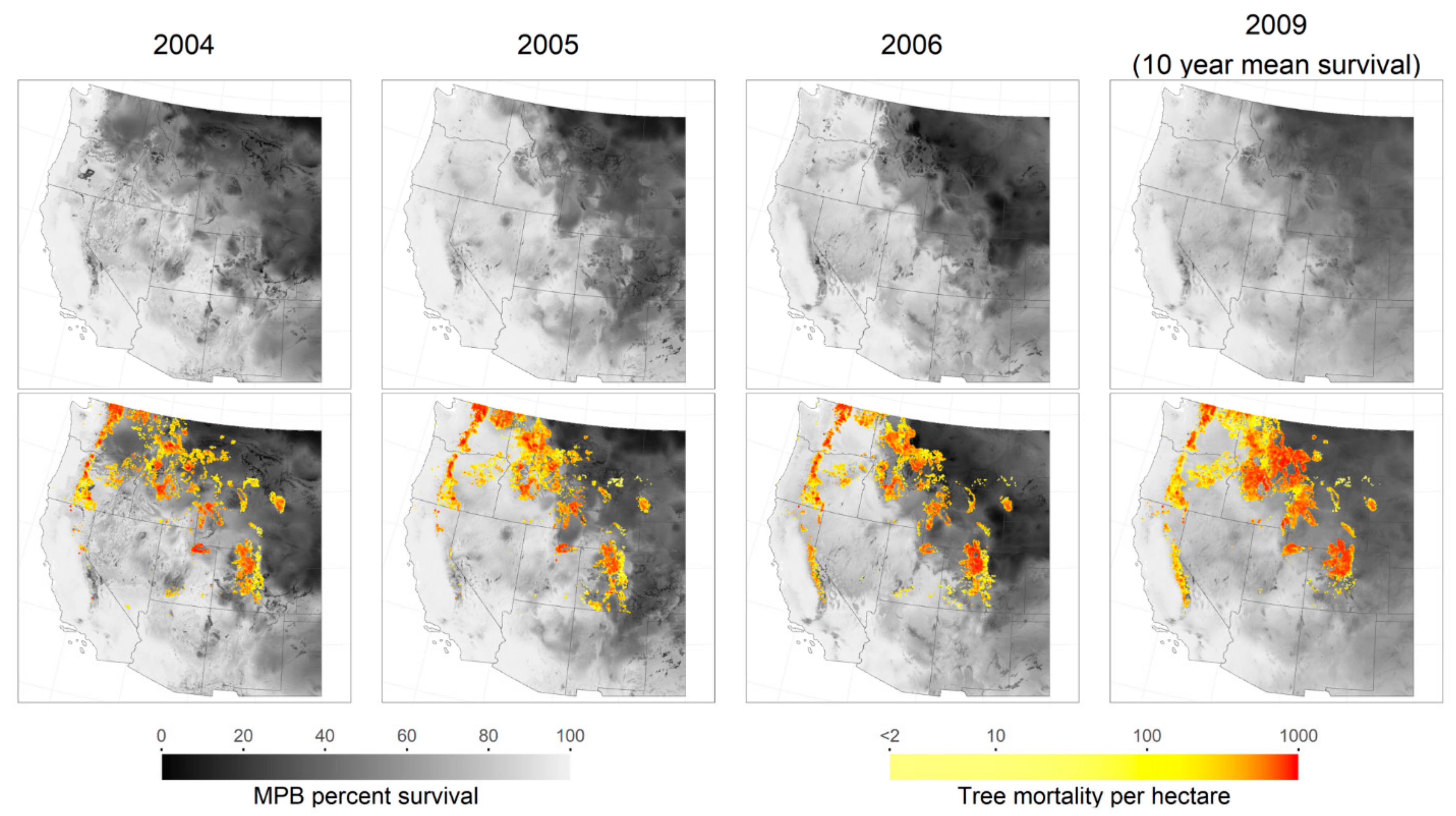
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bentz, B.J.; Régnière, J.; Fettig, C.J.; Hansen, E.M.; Hayes, J.L.; Hicke, J.A.; Kelsey, R.G.; Negrón, J.F.; Seybold, S.J. Climate change and bark beetles of the western United States and Canada: Direct and indirect effects. Bioscience 2010, 60, 602–613. [Google Scholar] [CrossRef]
- Safranyik, L.; Carroll, A.; Régnière, J.; Langor, D.; Riel, W.; Shore, T.; Peter, B.; Cooke, B.; Nealis, V.; Taylor, S. Potential for range expansion of mountain pine beetle into the boreal forest of North America. Can. Entomol. 2010, 142, 415–442. [Google Scholar] [CrossRef]
- MacDonald, G.M. Climate change and water in Southwestern North America special feature: Water, climate change, and sustainability in the southwest. Proc. Natl. Acad. Sci. USA 2010, 107, 21256–21262. [Google Scholar] [CrossRef] [PubMed]
- Weed, A.S.; Ayres, M.P.; Hicke, J.A. Consequences of climate change for biotic disturbances in North American forests. Ecol. Monogr. 2013, 83, 441–470. [Google Scholar] [CrossRef]
- Schlyter, P.; Stjernquist, I.; Bärring, L.; Jönsson, A.; Nilsson, C. Assessment of the impacts of climate change and weather extremes on boreal forests in northern Europe, focusing on Norway spruce. Clim. Res. 2006, 31, 75–84. [Google Scholar] [CrossRef]
- Bolte, A.; Ammer, C.; Löf, M.; Madsen, P.; Nabuurs, G.-J.; Schall, P.; Spathelf, P.; Rock, J. Adaptive forest management in central Europe: Climate change impacts, strategies and integrative concept. Scand. J. For. Res. 2009, 24, 473–482. [Google Scholar] [CrossRef]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.-J.; Rammer, W.; Verkerk, P.J. Increasing forest disturbances in Europe and their impact on carbon storage. Nat. Clim. Chang. 2014, 4, 806–810. [Google Scholar] [CrossRef]
- Morris, J.L.; Cottrell, S.; Fettig, C.J.; DeRose, R.J.; Mattor, K.M.; Carter, V.A.; Clear, J.; Clement, J.; Hansen, W.D.; Hicke, J.A.; et al. Bark beetles as agents of change in social-ecological systems. Front Ecol. Environ. 2018, 16, S34–S43. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Mooney, H.A.; Agard, J.; Capistrano, D.; Defries, R.S.; Díaz, S.; Dietz, T.; Duraiappah, A.K.; Oteng-Yeboah, A.; Pereira, H.M.; et al. Science for managing ecosystem services: Beyond the millennium ecosystem assessment. Proc. Natl. Acad. Sci. USA 2009, 106, 1305–1312. [Google Scholar] [CrossRef]
- Corbett, L.J.; Withey, P.; Lantz, V.A.; Ochuodho, T.O. The economic impact of the mountain pine beetle infestation in British Columbia: Provincial estimates from a CGE analysis. Forestry 2016, 89, 100–105. [Google Scholar] [CrossRef]
- Klutsch, J.G.; Negron, J.F.; Costello, S.L.; Rhoades, C.C.; West, D.R.; Popp, J.; Caissie, R. Stand characteristics and downed woody debris accumulations associated with a mountain pine beetle (Dendroctonus ponderosae Hopkins) outbreak in Colorado. For. Ecol. Manag. 2009, 258, 641–649. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Allen, C.D.; Franklin, J.F.; Frelich, L.E.; Harvey, B.J.; Higuera, P.E.; Mack, M.C.; Meentemeyer, R.K.; Metz, M.R.; Perry, G.L.; et al. Changing disturbance regimes, ecological memory, and forest resilience. Front. Ecol. Environ. 2016, 14, 369–378. [Google Scholar] [CrossRef]
- Pelz, K.A.; Rhoades, C.C.; Hubbard, R.M.; Smith, F.W. Severity of overstory mortality influences conifer recruitment and growth in mountain pine beetle-affected forests. Forests 2018, 9, 536. [Google Scholar] [CrossRef]
- Logan, J.A.; Powell, J.A. Ghost forests, global warming, and the mountain pine beetle (Coleoptera: Scolytidae). Am. Entomol. 2001, 47, 160–173. [Google Scholar] [CrossRef]
- Buotte, P.C.; Hicke, J.A.; Preisler, H.K.; Abatzoglou, J.T.; Raffa, K.F.; Logan, J.A. Climate influences on whitebark pine mortality from mountain pine beetle in the Greater Yellowstone Ecosystem. Ecol. Appl. 2016, 26, 2507–2524. [Google Scholar] [CrossRef]
- Sambaraju, K.R.; Carroll, A.L.; Aukema, B.H. Multiyear weather anomalies associated with range shifts by the mountain pine beetle preceding large epidemics. For. Ecol. Manag. 2019, 438, 86–95. [Google Scholar] [CrossRef]
- Safranyik, L.; Carroll, A. The biology and epidemiology of the mountain pine beelte in lodgepole pine forests. In The Mountain Pine Beetle: A Synthesis of Biology, Management, and Impacts on Lodgepole Pine; Safranyik, L., Wilson, W., Eds.; Canadian Forest Service Publications: Victoria, BC, Canada, 2006; pp. 3–66. [Google Scholar]
- Stahl, K.; Moore, R.; McKendry, I. Climatology of winter cold spells in relation to mountain pine beetle mortality in British Columbia, Canada. Clim. Res. 2006, 32, 13–23. [Google Scholar] [CrossRef]
- Preisler, H.K.; Hicke, J.A.; Ager, A.A.; Hayes, J.L. Climate and weather influences on spatial temporal patterns of mountain pine beetle populations in Washington and Oregon. Ecology 2012, 93, 2421–2434. [Google Scholar] [CrossRef]
- Bentz, B.J.; Mullins, D.E. Ecology of mountain pine beetle (Coleoptera: Scolytidae) cold hardening in the intermountain west. Environ. Entomol. 1999, 28, 577–587. [Google Scholar] [CrossRef]
- Carroll, A.L.; Raffa, K.F.; Aukema, B.H.; Bentz, B.J.; Hicke, J.A.; Turner, M.G.; Romme, W.H. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: The dynamics of bark beetle eruptions. Bioscience 2008, 58, 501–517. [Google Scholar]
- Logan, J.; Amman, G. A distribution model for egg development in mountain pine beetle. Can. Entomol. 1986, 118, 361–372. [Google Scholar] [CrossRef]
- Bentz, B.J.; Logan, J.A.; Amman, G.D. Temperature-dependent development of the mountain pine beetle (Coleoptera: Scolytidae) and simulation of its phenology. Can. Entomol. 1991, 123, 1083–1094. [Google Scholar] [CrossRef]
- Bolstad, P.V.; Bentz, B.J.; Logan, J.A. Modelling micro-habitat temperature for dendroctonus ponderosae (Coleoptera: Scolytidae). Ecol. Model. 1997, 94, 287–297. [Google Scholar] [CrossRef]
- Logan, J.A.; Bentz, B.J. Model analysis of mountain pine beetle (Coleoptera: Scolytidae) seasonality. Environ. Entomol. 1999, 28, 924–934. [Google Scholar] [CrossRef]
- Powell, J.; Jenkins, J.L.; Logan, J.A.; Bentz, B.J. Seasonal temperature alone can synchronize life cycles. Bull. Math. Boil. 2000, 62, 977–998. [Google Scholar] [CrossRef]
- Perkins, D.L.; Roberts, D.W. Predictive models of whitebark pine mortality from mountain pine beetle. For. Ecol. Manag. 2003, 174, 495–510. [Google Scholar] [CrossRef]
- Aukema, B.H.; Carroll, A.L.; Zheng, Y.; Zhu, J.; Raffa, K.F.; Moore, R.D.; Stahl, K.; Taylor, S.W. Movement of outbreak populations of mountain pine beetle: Influences of spatiotemporal patterns and climate. Ecography 2008, 31, 348–358. [Google Scholar] [CrossRef]
- Mitchell, R.G.; Waring, R.H.; Pitman, G.B. Thinning lodgepole pine increases tree vigor and resistance to mountain pine beetle. For. Sci. 1983, 29, 204–211. [Google Scholar]
- Régnière, J.; Bentz, B. Modeling cold tolerance in the mountain pine beetle, Dendroctonus ponderosae. J. Insect Physiol. 2007, 53, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Thornton, P.E.; Thornton, M.; Mayer, B.W.; Wei, Y.; Devarakonda, R.; Vose, R.S.; Cook, R.B. Daymet: Daily surface weather data on a 1-km grid for North America, Version 2. In Proceedings of the American Geophysical Union Fall Meeting, San Francisco, CA, USA, 15–19 December 2014. [Google Scholar]
- Meddens, A.J.; Hicke, J.A. Spatial and temporal patterns of Landsat-based detection of tree mortality caused by a mountain pine beetle outbreak in Colorado, USA. For. Ecol. Manag. 2014, 322, 78–88. [Google Scholar] [CrossRef]
- USDA Forest Service Pest Portal. Available online: https://www.fs.fed.us/foresthealth/applied-sciences/mapping-reporting/data-app-development.shtml (accessed on 5 January 2019).
- Little, E. Atlas of United States Trees, Conifers and Important Hardwoods; US Department of Agriculture Miscellaneous Publication: Washington, DC, USA, 1971. [Google Scholar]
- USDA Forest Service Geodata Clearinghouse. Available online: https://data.fs.usda.gov/geodata/ (accessed on 5 January 2019).
- Creeden, E.P.; Hicke, J.A.; Buotte, P.C. Climate, weather, and recent mountain pine beetle outbreaks in the western United States. For. Ecol. Manag. 2014, 312, 239–251. [Google Scholar] [CrossRef]
- Waring, R.H.; Pitman, G.B. Modifying lodgepole pine stands to change susceptibility to mountain pine beetle attack. Ecology 1985, 66, 889–897. [Google Scholar] [CrossRef]
- Christiansen, E.; Waring, R.H.; Berryman, A.A. Resistance of conifers to bark beetle attack: Searching for general relationships. For. Ecol. Manag. 1987, 22, 89–106. [Google Scholar] [CrossRef]
- Romo, C.; Bader, M.-F.; Pawson, S. Inner log temperatures vary with log direction and forest cover: Implications for predicting the phenology of saproxylic insects. Agric. For. Meteorol. 2019, 275, 329–339. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Model | Reference | Difference |
|---|---|---|---|
| Fairfield Ranger Station | 0.353 | 0.408 | 0.055 |
| Ketchum Ranger Station | 0.556 | 0.55 | −0.006 |
| Stanley | 0.184 | 0.205 | 0.021 |
| Banner Summit | 0.51 | 0.602 | 0.092 |
| Galena | 0.547 | 0.533 | −0.014 |
| Dollarhide Summit | 0.698 | 0.777 | 0.079 |
| Galena Summit | 0.613 | 0.652 | 0.039 |
| Vienna MINE | 0.61 | 0.623 | 0.013 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bone, C.; Nelson, M.F. Improving Mountain Pine Beetle Survival Predictions Using Multi-Year Temperatures Across the Western USA. Forests 2019, 10, 866. https://doi.org/10.3390/f10100866
Bone C, Nelson MF. Improving Mountain Pine Beetle Survival Predictions Using Multi-Year Temperatures Across the Western USA. Forests. 2019; 10(10):866. https://doi.org/10.3390/f10100866
Chicago/Turabian StyleBone, Christopher, and Michael France Nelson. 2019. "Improving Mountain Pine Beetle Survival Predictions Using Multi-Year Temperatures Across the Western USA" Forests 10, no. 10: 866. https://doi.org/10.3390/f10100866
APA StyleBone, C., & Nelson, M. F. (2019). Improving Mountain Pine Beetle Survival Predictions Using Multi-Year Temperatures Across the Western USA. Forests, 10(10), 866. https://doi.org/10.3390/f10100866