Application of Least-Cost Movement Modeling in Planning Wildlife Mitigation Measures along Transport Corridors: Case Study of Forests and Moose in Lithuania




Abstract
1. Introduction
2. Material and Methods
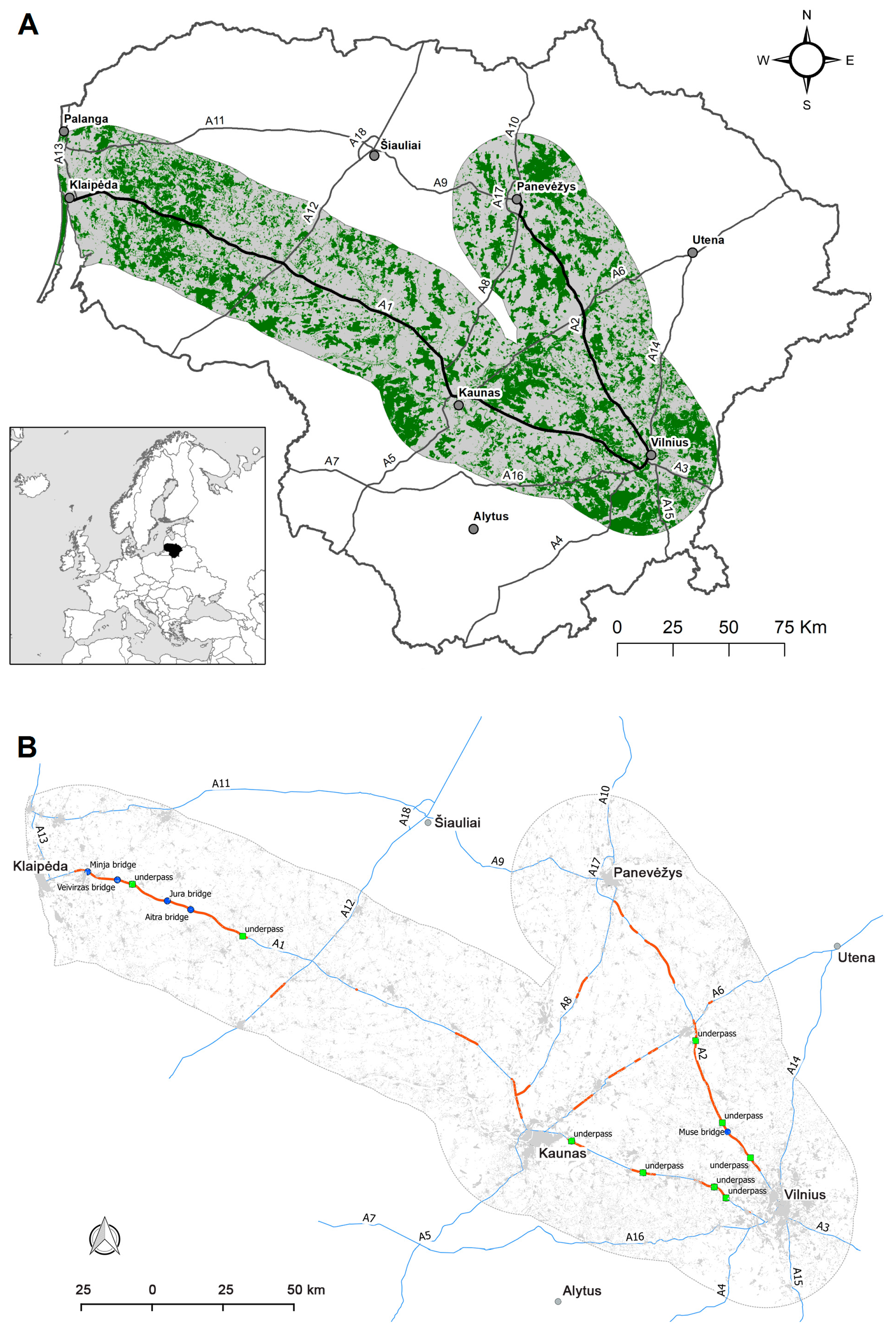
2.1. Study Area
2.2. Moose-Vehicle Collision Data
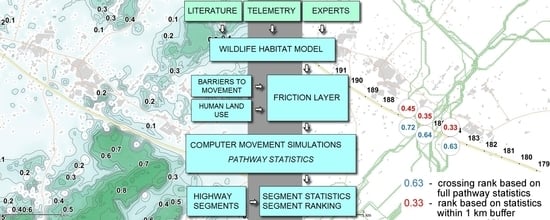
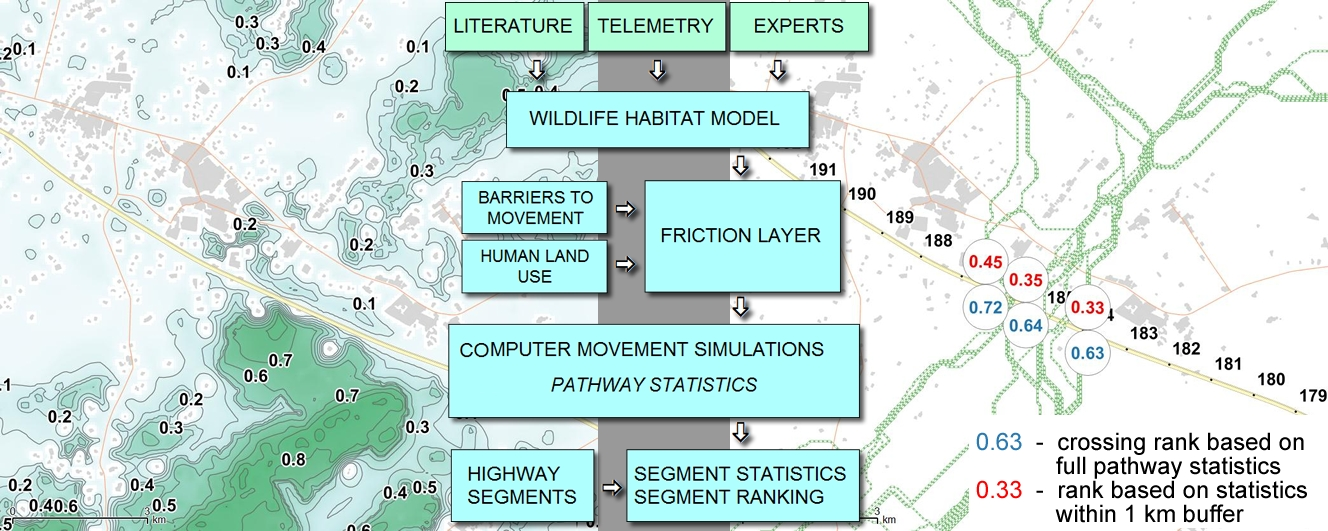
2.3. Workflow
- Selection of the most suitable model and creation of the moose habitat map;
- Adaptation of the selected model to the study area if required;
- Checking if habitats other than forests should be included on the map;
- Creation of the “potential” moose habitat map based on the habitat suitability index (HSI) values, and a “realized” habitat map, imposing the negative impact of human activity;
- Validation of the created moose habitat map (comparing actual moose densities to habitat suitability according to the model at the municipality level);
- Simulation of moose movements according to the habitat (using least cost principle and values of habitat suitability). The simulation was run twice, the first time based only on the realized habitat, the second one including wildlife fences as impermeable barriers to moose movement. As a result, moose movement pathways were obtained based on suitable habitats, and locations of the most probable moose crossing localities were defined;
- Validation/testing of the moose movement model (comparing the locations of actual MVC with the predicted zones of moose road crossing).
2.4. Model selection
2.5. Adaptation of Allen’s Model II for Lithuania
- 40% to 50% of the evaluation area is covered by sites with at least 50% of shrubs or forest <20 years old (S1); these early succession stages are assumed to provide abundant, preferred forage for moose;
- 5% to 15% of the evaluation area is dominated by spruce/fir over 20 years old (S2), providing optimal availability of winter cover for moose;
- 35% to 55% of the evaluation area is dominated by upland deciduous or mixed forest ≥20 years old (S3). These forest cover types are assumed to provide food as well as cover;
- 5% to 10% of the evaluation area is wetlands dominated by open water, emergent vegetation or submerged/floating-leaved hydrophytes (S4) [41].
2.6. Validation of the Moose Habitat Model
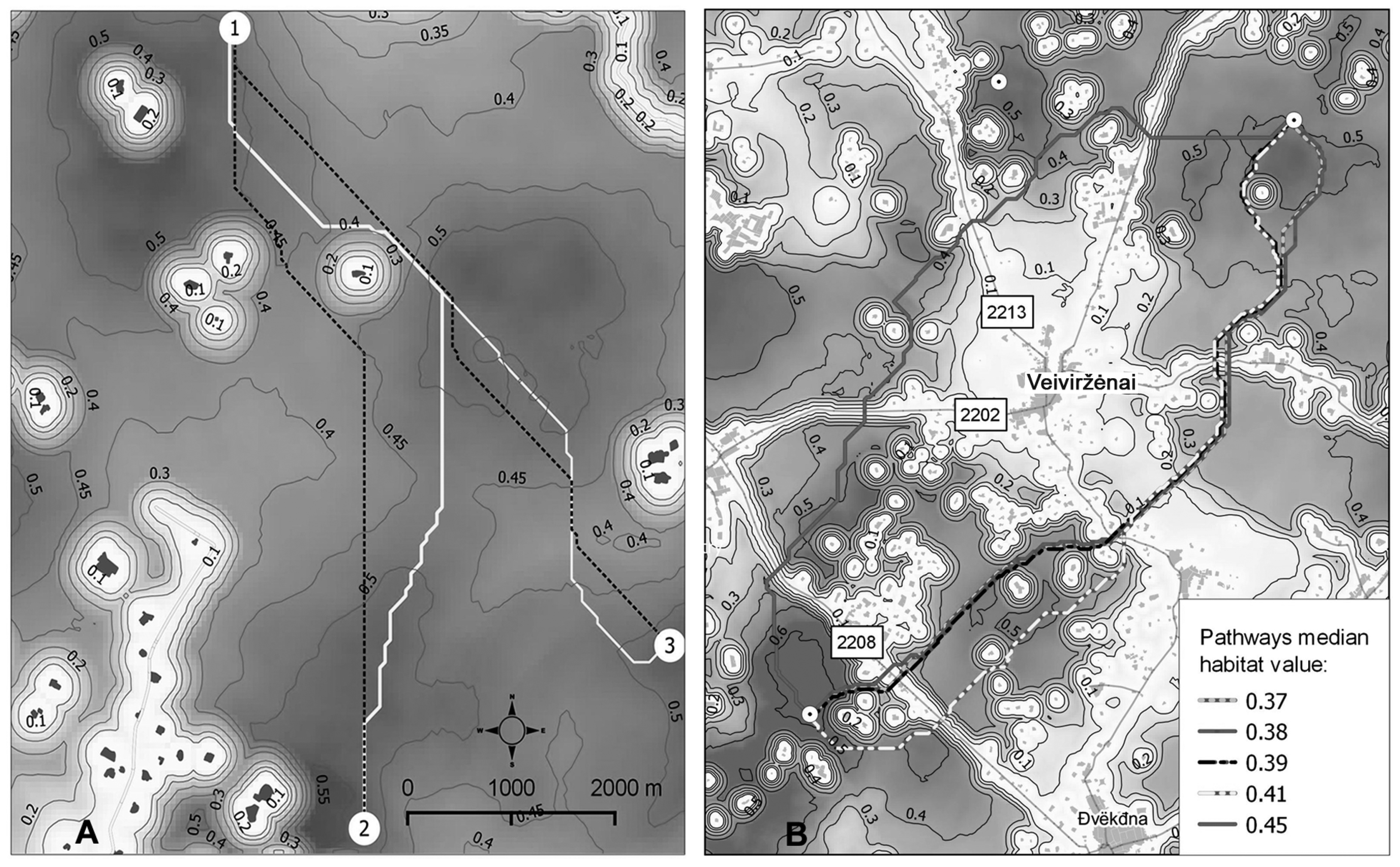
2.7. Moose Movement Simulation Model
2.8. Moose Movement Model Testing
2.9. Prognostic Value of the Moose Movement Model
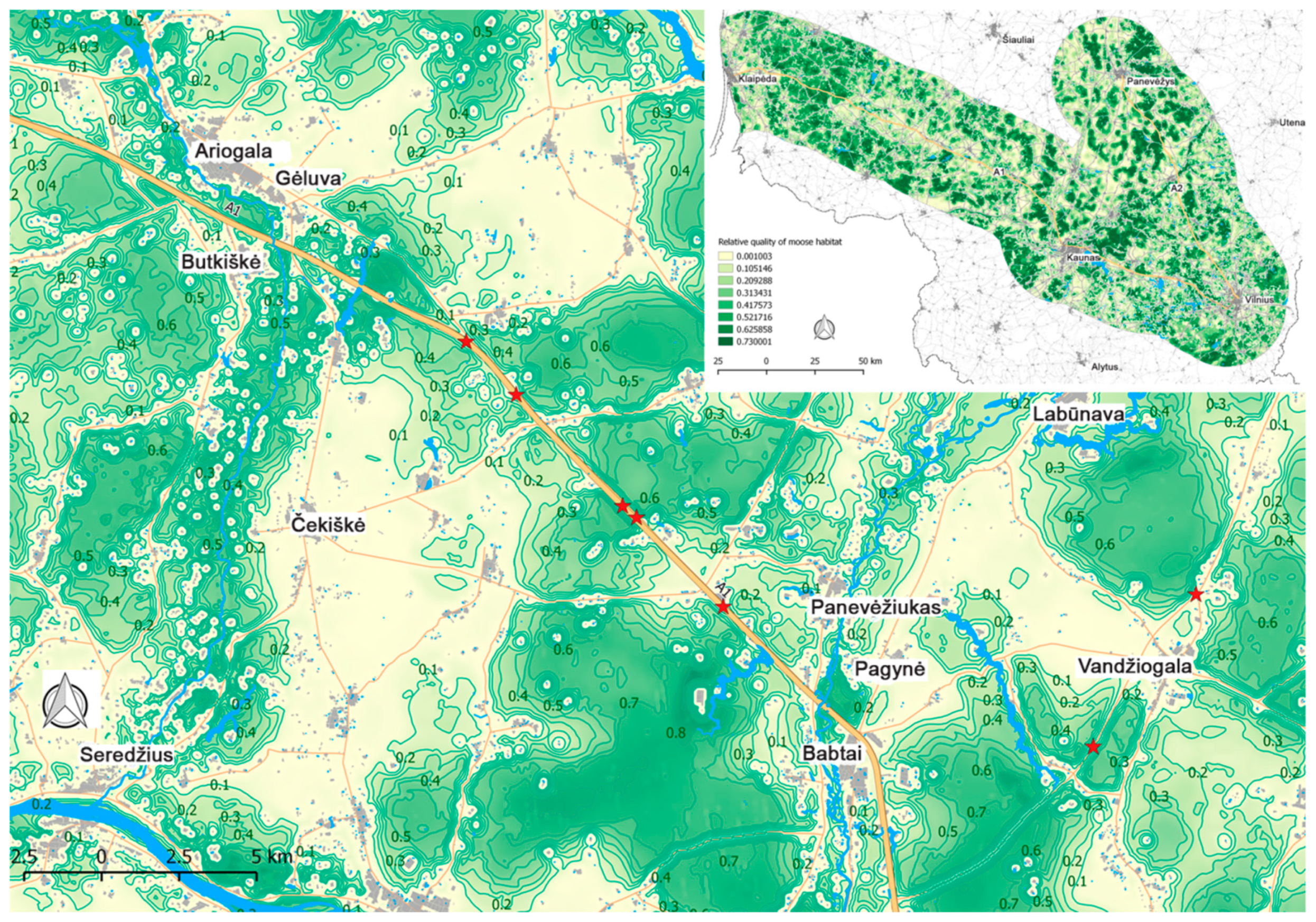
3. Results
3.1. Validity of the Moose Habitat Model
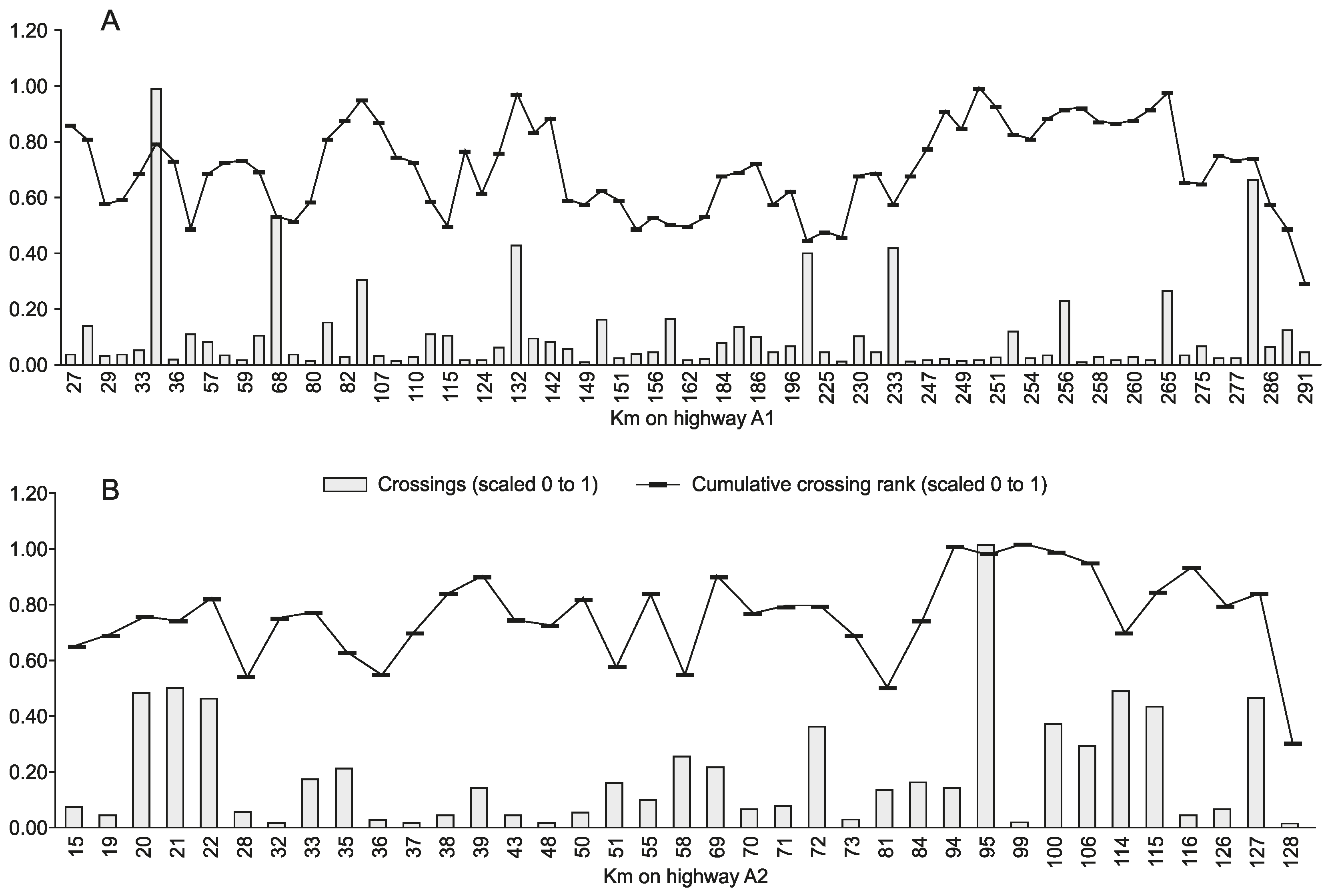
3.2. Moose Movement Model Results
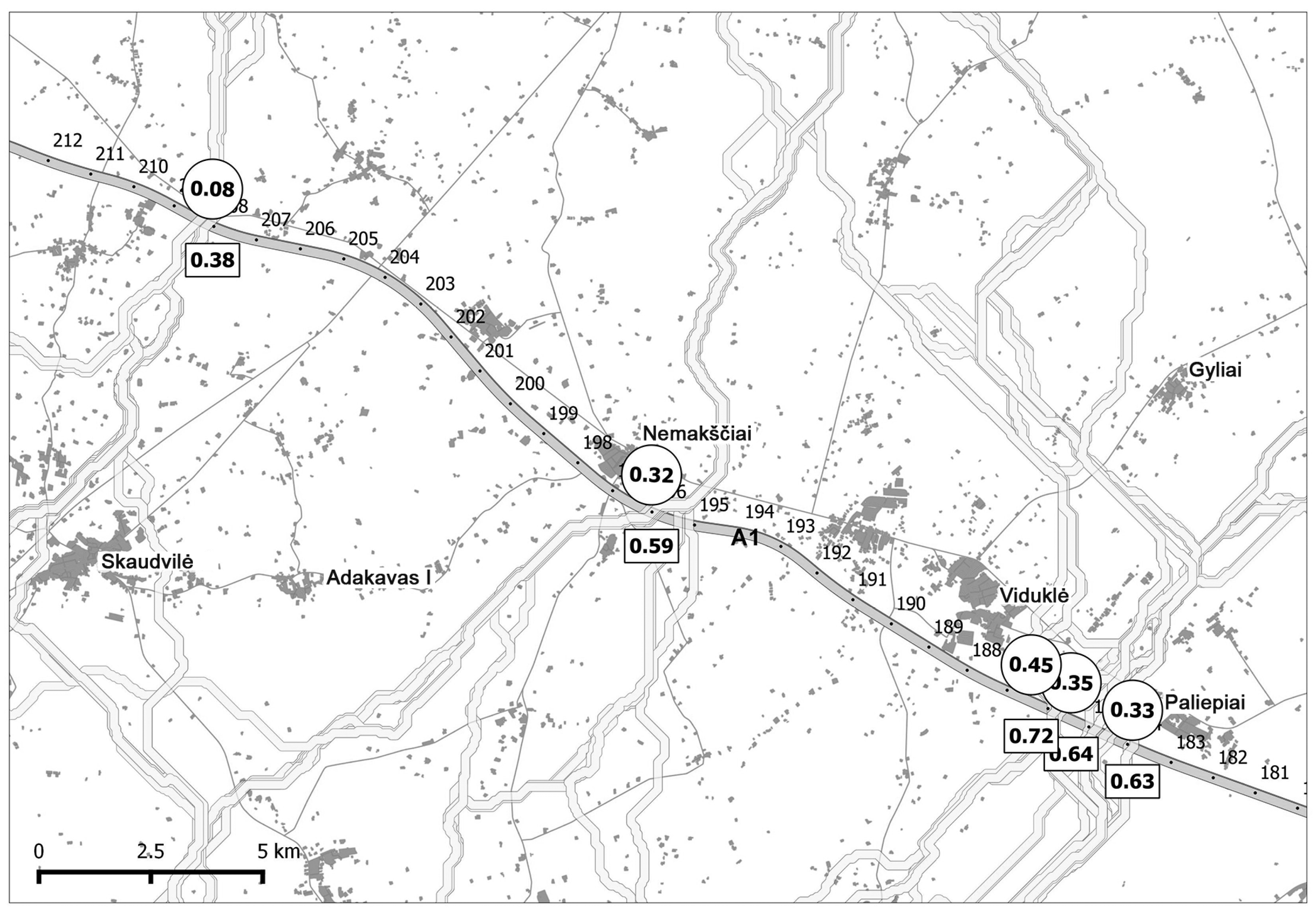
3.3. Moose Movement Model Testing Results
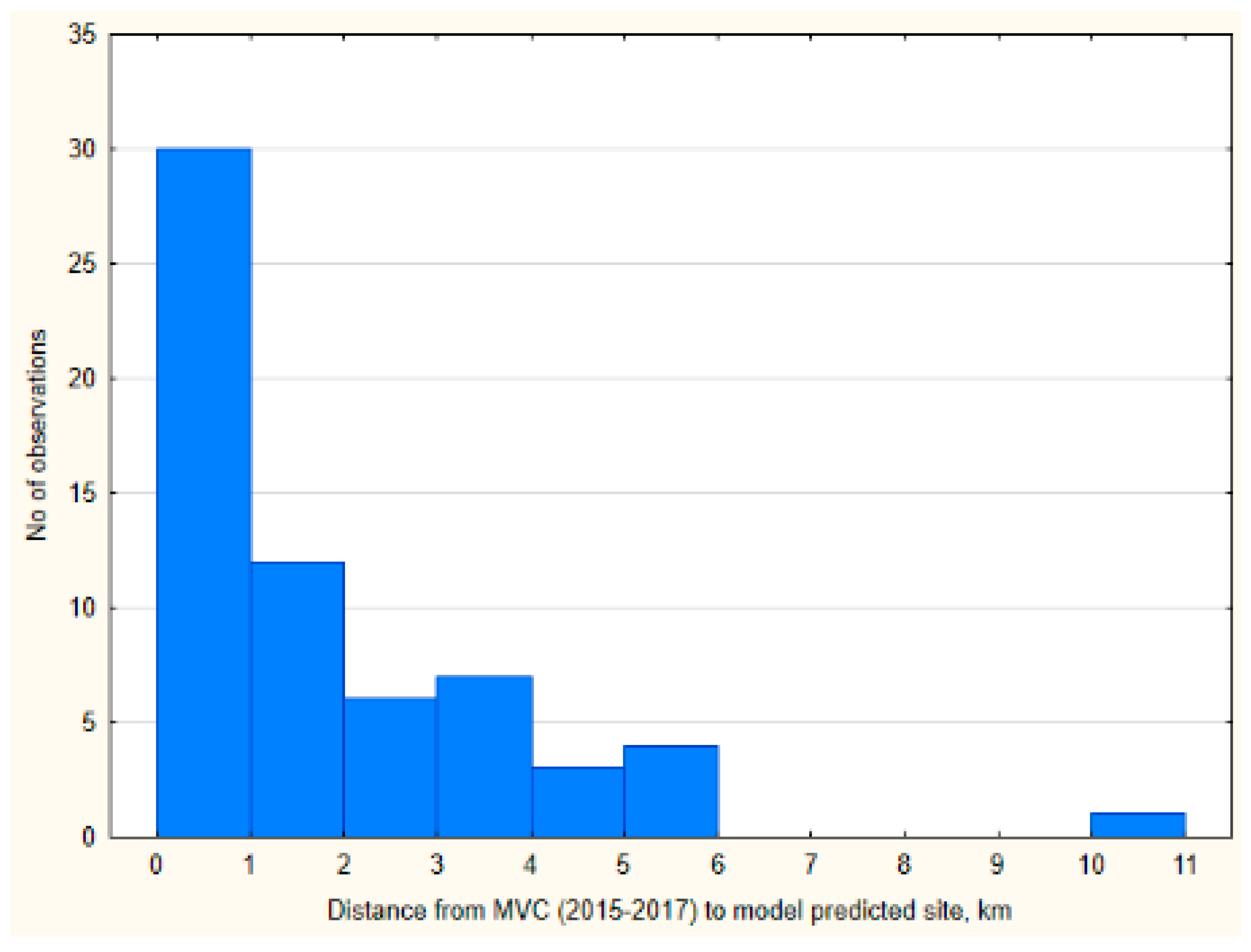
3.4. Prognostic Value of the Moose Movement Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Trombulak, S.C.; Frissell, C.A. Review of ecological effects of roads on terrestrial and aquatic communities. Conserv. Biol. 2000, 14, 18–30. [Google Scholar] [CrossRef]
- Fahrig, L.; Rytwinski, T. Effects of roads on animal abundance: An empirical review and synthesis. Ecol. Soc. 2009, 14, 21. [Google Scholar] [CrossRef]
- Rhodes, J.R.; Lunney, D.; Callaghan, J.; McAlpine, C.A. A few large roads or many small ones? How to accommodate growth in vehicle numbers to minimise impacts on wildlife. PLoS ONE 2014, 9, e91093. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.; Minor, E.S.; Woo, D.; Lee, D.; Park, C.R. Forest mammal roadkills as related to habitat connectivity in protected areas. Biodivers. Conserv. 2016, 25, 2673–2686. [Google Scholar] [CrossRef]
- Roger, E.; Laffan, S.W.; Ramp, D. Road impacts a tipping point for wildlife populations in threatened landscapes. Popul. Ecol. 2011, 53, 215–227. [Google Scholar] [CrossRef]
- World Road Statistics by International Road Federation. 2018. Available online: https://www.gihub.org/resources/data/world-road-statistics/ (accessed on 20 December 2018).
- Spellerberg, I.F. Ecological Effects of Roads; Taylor & Francis Inc.: Enfield, CT, USA, 2002; pp. 1–18. [Google Scholar]
- Seiler, A. The Toll of the Automobile: Wildlife and Roads in Sweden; Swedish University of Agricultural Sciences: Uppsala, Sweden, 2003; pp. 11–13. [Google Scholar]
- Polak, T.; Rhodes, J.R.; Jones, D.; Possingham, H.P. Optimal planning for mitigating the impacts of roads on wildlife. J. Appl. Ecol. 2014, 51, 726–734. [Google Scholar] [CrossRef]
- Forman, R.T.T.; Alexander, L.E. Roads and their major ecological effects. Annu. Rev. Ecol. Syst. 1998, 29, 207–231. [Google Scholar] [CrossRef]
- Damarad, T.; Bekker, G.J. COST 341—Habitat Fragmentation Due to Transportation Infrastructure: Findings of the COST Action 341; Office for Official Publications of the European Communities: Luxembourg, 2003. [Google Scholar]
- Tillmann, J.E. Habitat fragmentation and ecological networks in Europe. GAIA 2005, 14, 119–123. [Google Scholar] [CrossRef]
- Najafi, A.; Mohammadi Samani, K. Planning road network in mountain forests using GIS and Analytic Hierarchical Process (AHP). Casp. J. Environ. Sci. 2010, 8, 151–162. [Google Scholar]
- McRae, B.H.; Hall, S.A.; Beier, P.; Theobald, D.M. Where to restore ecological connectivity? Detecting barriers and quantifying restoration benefits. PLoS ONE 2012, 7, e52604. [Google Scholar] [CrossRef]
- Vilt-og-traffikk. 2016. Available online: http://hjortevilt.no/kategori/vilt-og-trafikk/ (accessed on 4 March 2016).
- Seiler, A. Predicting locations of moose-vehicle collisions in Sweden. J. Appl. Ecol. 2005, 42, 371–382. [Google Scholar] [CrossRef]
- Danks, Z.D.; Porter, W.F. Temporal, spatial, and landscape habitat characteristics of moose-vehicle collisions in Western Maine. J. Wildl. Manag. 2010, 74, 1229–1241. [Google Scholar] [CrossRef]
- Sielecki, L.E. WARS 1988–2007: Wildlife Accident Reporting and Mitigation in British Columbia, Special Report. Ministry of Transportation and Infrastructure, 2010. Available online: https://www2.gov.bc.ca/gov/content/transportation/transportation-infrastructure/engineering-standards-guidelines/environmental-management/wildlife-management/wildlife-accident-reporting-system/wars-1988-2007 (accessed on 10 July 2019).
- Rea, R.V.; Johnson, C.J.; Emmons, S. Characterizing moose–vehicle collision hotspots in northern British Columbia. J. Fish Wildl. Manag. 2014, 5, 46–58. [Google Scholar] [CrossRef]
- Kučas, A.; Balčiauskas, L. Temporal patterns of ungulate-vehicle collisions in Lithuania. J. Wildl. Manag. (re-submitted after reviews).
- Wilson, R.E.; Farley, S.D.; McDonough, T.J.; Talbot, S.L.; Barboza, P.S. A genetic discontinuity in moose (Alces alces) in Alaska corresponds with fenced transportation infrastructure. Conserv. Genet. 2015, 16, 791–800. [Google Scholar] [CrossRef]
- Wattles, D.W.; Zeller, K.A.; DeStefano, S. Response of moose to a high-density road network. J. Wildl. Manag. 2018, 82, 929–939. [Google Scholar] [CrossRef]
- Fryxell, J.M.; Hazell, M.; Börger, L.; Dalziel, B.D.; Haydon, D.T.; Morales, J.M.; McIntosh, T.; Rosatte, R.C. Multiple movement modes by large herbivores at multiple spatiotemporal scales. Proc. Natl. Acad. Sci. USA 2008, 105, 19114–19119. [Google Scholar] [CrossRef] [PubMed]
- Beier, P.; Majka, D.R.; Spencer, W.D. Forks in the road: Choices in procedures for designing wildland link. Conserv. Biol. 2008, 22, 836–851. [Google Scholar] [CrossRef]
- Yackulic, C.B.; Blake, S.; Deem, S.; Kock, M.; Uriart, M. One size does not fit all: Flexible models are required to understand animal movement across scales. J. Anim. Ecol. 2011, 80, 1088–1096. [Google Scholar] [CrossRef]
- Clevenger, A.P.; Wierzchowski, J.; Chruszcz, B.; Gunson, K. GIS-Generated, expert-based models for identifying wildlife habitat linkages and planning mitigation passages. Conserv. Biol. 2002, 16, 503–514. [Google Scholar] [CrossRef]
- Adriaensen, F.; Chardon, J.P.; De Blust, G.; Swinnen, E.; Villalba, S.; Gulinck, H.; Matthysen, E. The application of “least-cost” modeling as a functional landscape model. Landsc. Urban Plan. 2003, 64, 233–247. [Google Scholar] [CrossRef]
- Larkin, J.L.; Maehr, D.S.; Hoctor, T.S.; Orlando, M.A.; Whitney, K. Landscape linkages and conservation planning for the black bear in west-central Florida. Anim. Conserv. 2004, 7, 23–34. [Google Scholar] [CrossRef]
- Clevenger, A.P.; Wierzchowski, J. Maintaining and restoring connectivity in landscapes fragmented by roads. In Maintaining Connections for Nature; Crooks, K., Sanjayan, M., Eds.; Cambridge University Press: Cambridge, UK, 2006; pp. 502–535. [Google Scholar] [CrossRef]
- Theobald, D.M.; Reed, S.E.; Fields, K.; Soulé, M. Connecting natural landscapes using a landscape permeability model to prioritize conservation activities in the United States. Conserv. Lett. 2012, 5, 123–133. [Google Scholar] [CrossRef]
- Watts, K.; Eycott, A.E.; Handley, P.; Ray, D.; Humphrey, J.W.; Quine, C.P. Targeting and evaluating biodiversity conservation action within fragmented landscapes: An approach based on generic focal species and least-cost networks. Landsc. Ecol. 2010, 25, 1305–1318. [Google Scholar] [CrossRef]
- Stevenson-Holt, C.D.; Watts, K.; Bellamy, C.C.; Nevin, O.T.; Ramsey, A.D. Defining landscape resistance values in least-cost connectivity models for the invasive grey squirrel: A comparison of approaches using expert-opinion and habitat suitability modelling. PLoS ONE 2014, 9, e112119. [Google Scholar] [CrossRef] [PubMed]
- Zeller, K.A.; McGarigal, K.; Whiteley, A.R. Estimating landscape resistance to movement: A review. Landsc. Ecol. 2012, 27, 777–797. [Google Scholar] [CrossRef]
- Nikula, A.; Nivala, V.; Matala, J.; Heliövaara, K.T. Modelling the effect of habitat composition and roads on the occurrence and number of moose damage at multiple scales. Silva Fenn. 2019, 53, 9918. [Google Scholar] [CrossRef]
- Laforge, M.P.; Michel, N.L.; Wheeler, A.L.; Brook, R.K. Habitat selection by female moose in the Canadian prairie ecozone. J. Wildl. Manag. 2016, 80, 1059–1068. [Google Scholar] [CrossRef]
- Laforge, M.P.; Michel, N.L.; Brook, R.K. Spatio-temporal trends in crop damage inform recent climate-mediated expansion of a large boreal herbivore into an agro-ecosystem. Sci. Rep. 2017, 7, 15203. [Google Scholar] [CrossRef]
- Bjorge, R.R.; Anderson, D.; Herdman, E.; Stevens, S. Status and management of moose in the parkland and grassland natural regions of Alberta. Alces 2018, 54, 71–84. [Google Scholar]
- Leoniak, G.; Barnum, S.; Atwood, J.L.; Rinehart, K.; Elbroch, M. Testing GIS-generated least-cost path predictions for Martes pennanti (Fisher) and its application for identifying mammalian road-crossings in northern New Hampshire. Northeast. Nat. 2012, 19, 147–157. [Google Scholar] [CrossRef]
- Cushman, S.A.; Lewis, J.S.; Landguth, E.L. Evaluating the intersection of a regional wildlife connectivity network with highways. Mov. Ecol. 2013, 1, 12. [Google Scholar] [CrossRef] [PubMed]
- Vanthomme, H.P.A.; Nzamba, B.S.; Alonso, A.; Todd, A.F. Empirical selection between least-cost and current flow designs for establishing wildlife corridors in Gabon. Conserv. Biol. 2018, 33, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Allen, A.W.; Peter, A.; Jordan, J.W.T. Habitat Suitability Index Models: Moose, Lake Superior Region. 1987. Available online: http://oai.dtic.mil/oai/oai?verb=getRecord&metadataPrefix=html&identifier=ADA323425 (accessed on 12 August 2015).
- Allen, A.W.; Terrell, J.W.; Mangus, W.L.; Lindquist, E.L. Application and partial validation of a habitat model for moose in the Lake Superior region. Alces 1991, 27, 50–64. [Google Scholar]
- Hickey, L. Assessing re-colonization of moose in New York with HSI models. Alces 2008, 44, 117–126. [Google Scholar]
- Dettki, H.; Löfstrand, R.; Edenius, L. Modeling habitat suitability for moose in coastal northern Sweden: Empirical vs process-oriented approaches. Ambio 2003, 32, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Andersone-Lilley, Ž.; Balčiauskas, L.; Ozoliņš, J.; Randveer, T.; Tönisson, J. Ungulates and their management in the Baltics (Estonia, Latvia and Lithuania). In European Ungulates and Their Management in the 21st Century; Apollonio, M., Andersen, R., Putman, R., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 103–128. [Google Scholar]
- Cederlund, G.; Sandegren, F.; Larsson, K. Summer movements of moose and dispersal of their offspring. J. Wildl. Manag. 1987, 51, 342–352. [Google Scholar] [CrossRef]
- Eastman, J.R. Idrisi Kilimanjaro. Guide to GIS and Image Processing; Clark Labs, Clark University: Worcester, MA, USA, 2003; pp. 1–328. [Google Scholar]
- Spear, S.F.; Balkenhol, N.; Fortin, M.J.; McRae, B.H.; Scribner, K. Use of resistance surfaces for landscape genetic studies: Considerations for parameterization and analysis. Mol. Ecol. 2010, 19, 3576–3591. [Google Scholar] [CrossRef] [PubMed]
- Saaty, T.L. A scaling method for priorities in hierarchical structures. J. Math. Psychol. 1977, 15, 234–281. [Google Scholar] [CrossRef]
- Bíl, M.; Andrášik, R.; Svoboda, T.; Sedoník, J. The KDE+ software: A tool for effective identification and ranking of animal-vehicle collision hotspots along networks. Landsc. Ecol. 2016, 31, 231–237. [Google Scholar] [CrossRef]
- Bíl, M.; Andrášik, R.; Nezval, V.; Bílová, M. Identifying locations along railway networks with the highest tree fall hazard. Appl. Geogr. 2017, 87, 45–53. [Google Scholar] [CrossRef]
- Laurian, C.; Dussault, C.; Ouellet, J.-P.; Courtois, R.; Poulin, M.; Breton, L. Behaviour of moose relative to a road network. J. Wildl. Manag. 2008, 72, 1550–1557. [Google Scholar] [CrossRef]
- Bartzke, G.S.; May, R.; Solberg, E.J.; Rolandsen, C.M.; Røskaft, E. Differential barrier and corridor effects of power lines, roads and rivers on moose (Alces alces) movements. Ecosphere 2015, 6, 1–17. [Google Scholar] [CrossRef]
- Wattles, D.; DeStefano, S. Use and movements of moose in Massachusetts: Implications for conservation of large mammals in a fragmented environment. Alces 2013, 49, 65–81. [Google Scholar]
- Crum, N.J.; Fuller, A.K.; Sutherland, C.S.; Cooch, E.G.; Hurst, J. Estimating occupancy probability of moose using hunter survey data. J. Wildl. Manag. 2017, 81, 521–534. [Google Scholar] [CrossRef]
- Neumann, W.; Ericsson, G.; Dettki, H.; Radeloff, V.C. Behavioural response to infrastructure of wildlife adapted to natural disturbances. Landsc. Urban Plan. 2013, 114, 9–27. [Google Scholar] [CrossRef]
- Yost, A.C.; Wright, R.G. Moose, caribou, and grizzly bear distribution in relation to road traffic in Denali National Park, Alaska. Arctic 2001, 54, 41–48. [Google Scholar] [CrossRef]
- Ball, J.; Nordengren, C.; Wallin, K. Partial migration by large ungulates: Characteristics of seasonal moose Alces alces ranges in northern Sweden. Wildl. Biol. 2001, 7, 39–47. [Google Scholar] [CrossRef]
- Bjørneraas, K.; Solberg, E.J.; Herfindal, I.; Van Moorter, B.; Rolandsen, C.M.; Tremblay, J.P.; Skarpe, C.; Sæther, B.E.; Eriksen, R.; Astrup, R. Moose Alces alces habitat use at multiple temporal scales in a human-altered landscape. Wildl. Biol. 2011, 17, 44–54. [Google Scholar] [CrossRef]
- Kantar, L. Broccoli and moose, not always best served together: Implementing a controlled moose hunt in Maine. Alces 2011, 47, 83–90. [Google Scholar]
- Hamilton, G.D.; Drysdale, P.D.; Euler, D.L. Moose winter browsing patterns on clear-cuttings in northern Ontario. Can. J. Zool. 1980, 158, 1412–1416. [Google Scholar] [CrossRef]
- Balčiauskas, L. Distribution of species-specific wildlife–vehicle accidents on Lithuanian roads, 2002–2007. Est. J. Ecol. 2009, 58, 157–168. [Google Scholar] [CrossRef]
- Dussault, C.; Courtois, R.; Ouellet, J.-P. A habitat suitability index model to assess moose habitat selection at multiple spatial scales. Can. J. For. Res. 2006, 36, 1097–1107. [Google Scholar] [CrossRef]
- Hurley, M.V.; Rapaport, E.K.; Johnson, C.J. A spatial analysis of moose-vehicle collisions in Mount Revelstoke and Glacier National Parks, Canada. Alces 2007, 43, 79–100. [Google Scholar]
- Balčiauskas, L. The influence of roadkill on protected species and other wildlife in Lithuania. In Proceedings of the 2011 International Conference on Ecology and Transportation, Seattle, WA, USA, 21–25 August 2011; Wagner, P.J., Nelson, D., Murray, E., Eds.; North Carolina State University, Center for Transportation and the Environment: Raleigh, NC, USA, 2012; pp. 647–655. [Google Scholar]
- DeMars, C.A.; Serrouya, R.D.; Mumma, M.A.; Gillingham, M.P.; McNay, R.S.; Boutin, S. Moose, caribou and fire: Have we got it right yet? Can. J. Zool. 2019. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Full Length of Pathways | Area within 500 m Buffer Each Side of Highways | ||
|---|---|---|---|
| Layer | Weight | Layer | Weight |
| Moose habitat | 0.3787 | Moose habitat, side A | 0.2940 |
| Path proportion in hiding cover | 0.3645 | Moose habitat, side B | 0.2940 |
| Distance to human development | 0.1516 | Distance to human development, side A | 0.0588 |
| Number of roads crossed | 0.0771 | Distance to human development, side B | 0.0588 |
| Pathway complexity | 0.0281 | Path proportion in hiding cover | 0.2941 |
| Year | Cluster | Fencing | Rc | Npath | Dist. | ||
|---|---|---|---|---|---|---|---|
| Strength | Start | End | |||||
| 2017 | 0.50 | 44.8 | 45.0 | no | 0.4 | 40 | 0 |
| 2016 | 0.67 | 135.8 | 136.0 | no | 0.82 | 35 | 2 |
| 2016 | 0.50 | 130.3 | 130.5 | no | 0.97 | 132 | < 1 |
| 2015 | 0.50 | 136.4 | 136.6 | no | 0.82 | 35 | 1.5 |
| Road | Cluster | Fencing | Rc | Npath | Dist. | ||
|---|---|---|---|---|---|---|---|
| Strength | Start | End | |||||
| A1 | 0.67 | 135.8 | 136.0 | no | 0.82 | 35 | 2 |
| A1 | 0.50 | 44.8 | 45.0 | no | 0.4 | 40 | 0 |
| A1 | 0.50 | 136.4 | 136.6 | no | 0.82 | 35 | 1.5 |
| A1 | 0.50 | 130. 3 | 130.5 | no | 0.97 | 132 | < 1 |
| A1 | 0.50 | 265.4 | 265.6 | yes | 1.0 | 95 | 0 |
| A2 | 0.44 | 46.7 | 46.9 | yes | 0.92 | 22 | 5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wierzchowski, J.; Kučas, A.; Balčiauskas, L. Application of Least-Cost Movement Modeling in Planning Wildlife Mitigation Measures along Transport Corridors: Case Study of Forests and Moose in Lithuania. Forests 2019, 10, 831. https://doi.org/10.3390/f10100831
Wierzchowski J, Kučas A, Balčiauskas L. Application of Least-Cost Movement Modeling in Planning Wildlife Mitigation Measures along Transport Corridors: Case Study of Forests and Moose in Lithuania. Forests. 2019; 10(10):831. https://doi.org/10.3390/f10100831
Chicago/Turabian StyleWierzchowski, Jack, Andrius Kučas, and Linas Balčiauskas. 2019. "Application of Least-Cost Movement Modeling in Planning Wildlife Mitigation Measures along Transport Corridors: Case Study of Forests and Moose in Lithuania" Forests 10, no. 10: 831. https://doi.org/10.3390/f10100831
APA StyleWierzchowski, J., Kučas, A., & Balčiauskas, L. (2019). Application of Least-Cost Movement Modeling in Planning Wildlife Mitigation Measures along Transport Corridors: Case Study of Forests and Moose in Lithuania. Forests, 10(10), 831. https://doi.org/10.3390/f10100831