Scaling Approach for Estimating Stand Sapwood Area from Leaf Area Index in Five Boreal species



Abstract
:1. Introduction
2. Materials and Methods
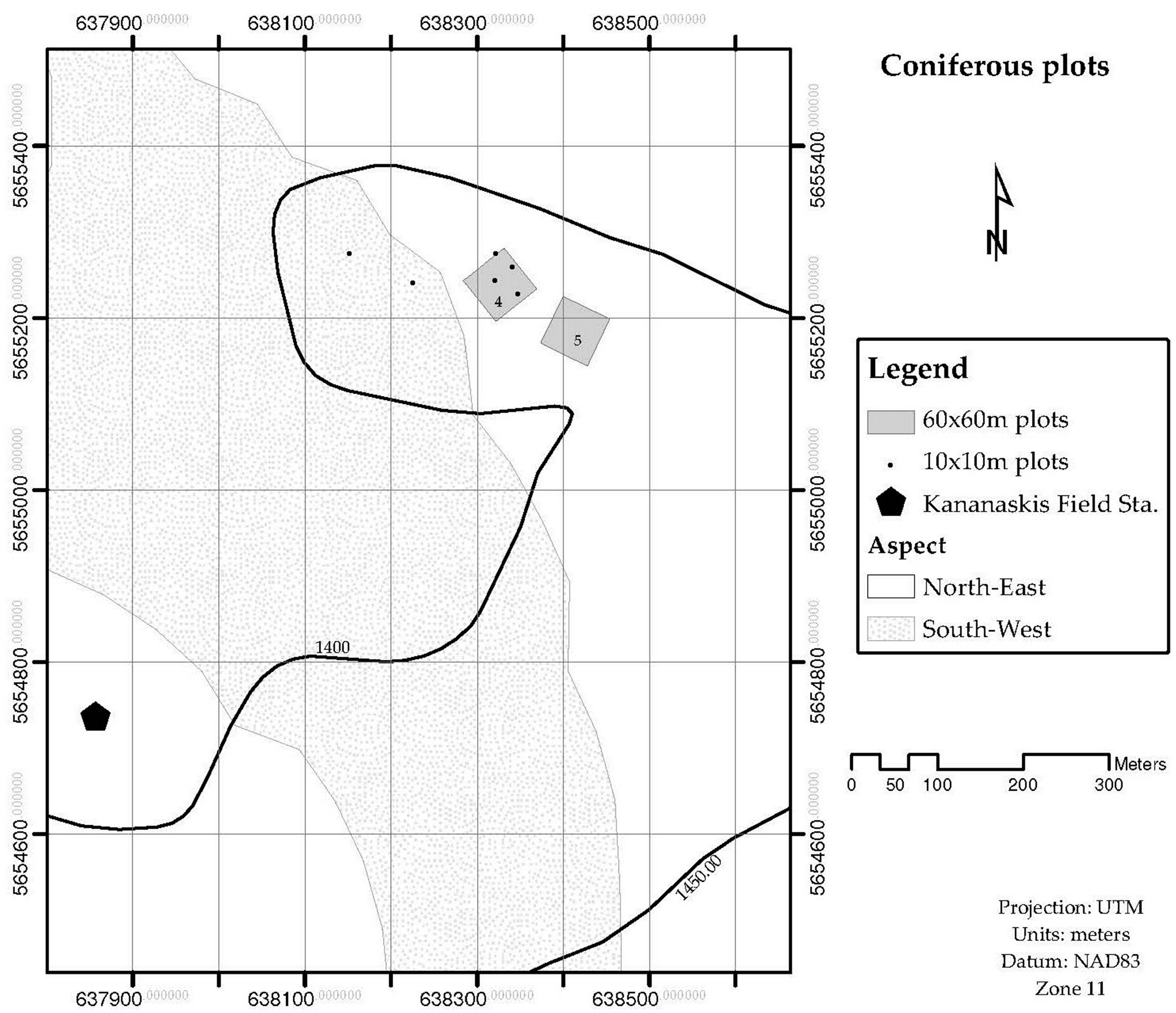
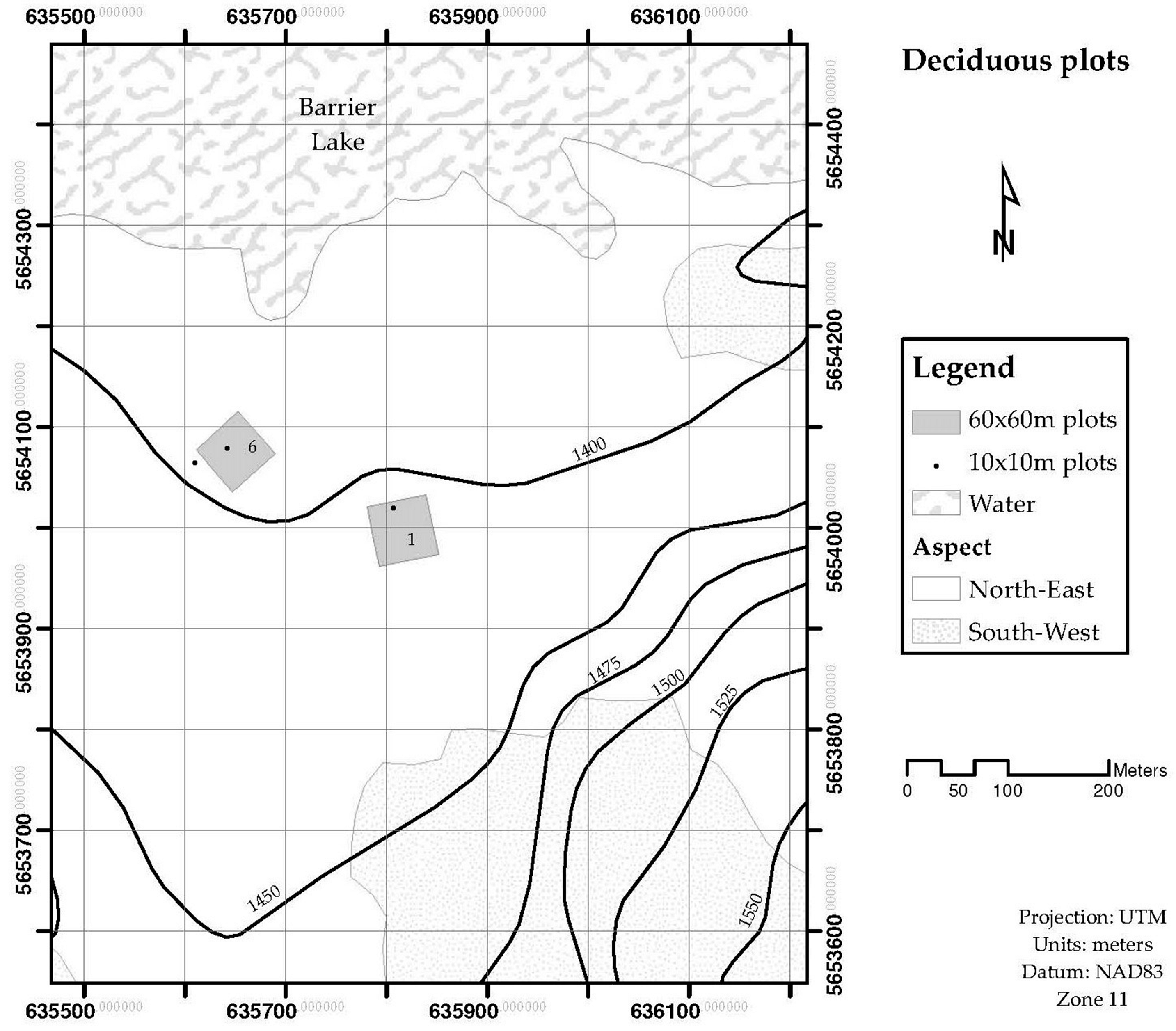
2.1. Model Approach and Sampling Design
2.2. Treatment of Saplings
2.3. Statistical Analysis
3. Model Implementation and Data Results
3.1. Tree-Level Allometric Correlations
3.2. Stand-Level Allometric Correlations
3.2.1. Scaling Up Sapwood Area from the Tree to the Stand Level
3.2.2. Leaf Area Estimates at the Stand-Level
3.2.3. Stand Level Leaf Area Sapwood Area Allometric Correlations
3.2.4. COV Confidence Intervals
3.3. Estimates of Error Propagation
4. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Margolis, H.; Oren, R.; Whitehead, D.; Kaufmann, M.R. Leaf area dynamics of conifer forests. In Ecophysiology of Coniferous Forests, Physical Ecology, 1st ed.; Smith, W.K., Hinckley, T.M., Eds.; Academic Press Inc.: San Diego, CA, USA, 1995; pp. 181–223. ISBN 978-0-08-092593-6. [Google Scholar]
- Whitehead, D. The estimation of foliage area from sapwood basal area in scots pine. Forestry 1978, 51, 137–149. [Google Scholar] [CrossRef]
- Waring, R. Estimating forest growth and efficiency in relation to canopy leaf area. Adv. Ecol. Res. 1983, 13, 327–354. [Google Scholar] [CrossRef]
- Blanche, C.A.; Hodges, J.D.; Nebeker, T.E. A leaf area-sapwood area ratio developed to rate Loblolly Pine tree vigor. Can. J. For. Res. 1985, 15, 1181–1184. [Google Scholar] [CrossRef]
- Borghetti, M.; Vendramin, G.G.; Giannini, R. Specific leaf area and leaf area index distribution in a young Douglas fir plantation. Can. J. For. Res. 1986, 16, 1283–1288. [Google Scholar] [CrossRef]
- McDowell, N.; Barnard, H.; Bond, B.J.; Hinckley, T.; Hubbard, R.M.; Ishii, H.; K¨ostner, B.; Magnani, F.; Marshall, J.D.; Meinzer, F.C.; et al. The relationships between tree height and leaf area: Sapwood area ratio. Oecologia 2002, 132, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Dean, T.J.; Long, J.N.; Smith, F.W. Bias in leaf area-sapwood area ratios and its impact on growth analysis in Pinus contorta. Trees 1988, 2, 104–109. [Google Scholar] [CrossRef]
- Hungerford, R.D. Estimation of foliage area in dense Montana Lodgepole Pine stands. Can. J. For. Res. 1987, 17, 320–324. [Google Scholar] [CrossRef]
- Keane, M.G.; Weetman, G.F. Leaf area-sapwood cross-sectional area relationships in repressed stands of Lodgepole pine. Can. J. For. Res. 1987, 17, 205–209. [Google Scholar] [CrossRef]
- Kaufman, M.R.; Troendle, C. The relationship of leaf area and foliage biomass to sapwood conducting area in four subalpine forest tree species. For. Sci. 1981, 27, 477–482. [Google Scholar]
- Schuler, T.M.; Smith, F.W. Effect of species mix on size/density and leaf-area relations in southwest pinyon/juniper woodlands. For. Ecol. Manag. 1988, 25, 211–220. [Google Scholar] [CrossRef]
- Coyea, M.R.; Margolis, H.A. Factors affecting the relationship between sapwood area and leaf area of Balsam fir. Can. J. For. Res. 1992, 22, 1684–1693. [Google Scholar] [CrossRef]
- Marchand, J.P. Sapwood area as an estimator of foliage biomass and projected leaf area for Abies balsamea and Picea rubens. Can. J. For. Res. 1984, 14, 85–87. [Google Scholar] [CrossRef]
- Vertessy, R.A.; Benyon, R.G.; O’Sullivan, S.K.; Gribben, P.R. Relationships between stem diameter, sapwood area, leaf area and transpiration in a young mountain ash forest. Tree Physiol. 1995, 15, 559–567. [Google Scholar] [CrossRef]
- Meadows, S.J.; Hodges, D.J. Sapwood area as an estimator of leaf area and foliar weight in Cherrybark oak and green ash. For. Sci. 2002, 48, 69–76. [Google Scholar]
- Binkley, D.; Reid, P. Long-term responses of stem growth and leaf area to thinning and fertilization in a Douglas fir plantation. Can. J. For. Res. 1984, 14, 656–660. [Google Scholar] [CrossRef]
- Shelburne, V.B.; Hedden, R.L.; Allen, R.M. The effects of site, stand density and sapwood permeability on the relationship between leaf area and sapwood area in loblolly pine (Pinus taeda L.). For. Ecol. Manag. 1993, 58, 193–209. [Google Scholar] [CrossRef]
- White, D.; Beadle, C.; Worledge, D.; Honeysett, J.; Cherry, M. The influence of drought on the relationship between leaf and conducting sapwood area in Eucalyptus globulus and Eucalyptus nitens. Trees 1998, 12, 406–414. [Google Scholar] [CrossRef]
- Mencuccini, M.; Grace, J. Climate influences the leaf area/sapwood area ratio in scots pine. Tree Physiol. 1994, 15, 1–10. [Google Scholar] [CrossRef]
- Hillis, W.E. Heartwood and Tree Exudates; Series in Wood Science; Springer: Berlin, Germany, 1987; ISBN 3-540-17593-8. [Google Scholar]
- Dean, T.J.; Long, J.N. Variation in sapwood area-leaf area relations within two stands of Lodgepole pine. For. Sci. 1986, 32, 749–758. [Google Scholar]
- Lefsky, M.A.; Cohen, W.B.; Acker, S.A.; Parker, G.G.; Spies, T.A.; Harding, D. Lidar remote sensing of the canopy structure and biophysical properties of Douglas fir western hemlock forests. Remote Sens. Environ. 1999, 70, 339–361. [Google Scholar] [CrossRef]
- Means, J.E.; Acker, S.A.; Harding, D.J.; Blair, J.B.; Lefsky, M.A.; Cohen, W.B.; Harmon, M.E.; McKee, W.A. Use of large-footprint scanning airborne LIDAR to estimate forest stand characteristics in the western cascades of Oregon. Remote Sens. Environ. 1999, 67, 298–308. [Google Scholar] [CrossRef]
- Lassen, L.E.; Okkonen, E.A. Sapwood Thickness of Douglas Fir and Five Other Western Softwoods; Research Paper FPL-124; US Dept. of Agriculture, Forest Service, Forest Products Laboratory: Madison, WI, USA, 1969; p. 16.
- Domec, J.C.; Lachenbruch, B.; Pruyn, M.; Spicer, R. Effects of age-related increases on sapwood area, leaf area, and xylem conductivity on height-related hydraulic costs in two contrasting coniferous species. Ann. For. Sci. 2012, 69, 17–27. [Google Scholar] [CrossRef]
- Jaskierniak, D.; Kuczera, G.; Benyon, R.G.; Lucieer, A. Estimating tree and stand sapwood area in spatially heterogeneous southeastern Australian forests. J. Plant Ecol. 2016, 9, 272–284. [Google Scholar] [CrossRef]
- Jaskierniak, D.; Benyon, R.; Kuczera, G.; Robinson, A. A new method for measuring stand sapwood area in forests. Ecohydrology 2015. [Google Scholar] [CrossRef]
- Jaskierniak, D.; Kuczera, G.; Benyon, R.; Wallace, L. Using tree detection algorithms to predict sapwood area, basal area and stocking density in Eucalyptus regnans forest. Remote Sens. 2015, 7, 7298–7323. [Google Scholar] [CrossRef]
- Mitchell, P.J.; Lane, P.N.J.; Benyon, R.G. Capturing within catchment variation in evapotranspiration from montane forests using LIDAR canopy profiles with measured and modelled fluxes of water. Ecohydrology 2012, 5, 708–720. [Google Scholar] [CrossRef]
- Cˇerm’ak, J.; Nadezhdina, N. Sapwood as the scaling parameter defining according to xylem water content or radial pattern of sap flow? Ann. Sci. For. 1998, 55, 509–521. [Google Scholar]
- Quiñonez-Piñón, M.R.; Valeo, C. Allometry of sapwood depth in five boreal trees. Forests 2017, 8, 457. [Google Scholar] [CrossRef]
- Jonckheere, I.; Fleck, S.; Nackaerts, K.; Muysa, B.; Coppin, P.; Weiss, M.; Baret, F. Review of methods for in situ leaf area index determination. Part I. Theories, sensors and hemispherical photography. Agric. For. Meteorol. 2004, 121, 19–35. [Google Scholar] [CrossRef]
- Peet, R.K. Forests and Meadows of the Rocky Mountains. In North American Terrestrial Vegetation, 1st ed.; Barbour, M.G., Billings, W.D., Eds.; Cambridge University Press: New York, NY, USA, 1988; ISBN 13-978-0-52-126198-2. [Google Scholar]
- Rowe, J.S. Forest Regions of Canada; Department of the Environment Canadian Forestry Service: Ottawa, ON, Canada, 1972; p. 177.
- Strong, W.L.; Leggat, K.R. Ecoregions of Alberta. In Forestry, Lands, and Wildlife (1986–1993); The Provincial Archives of Alberta: Edmonton, AB, Canada, 1992; p. 59. [Google Scholar]
- Elliot-Frisk, D.L. The taiga and boreal forest. In North American Terrestrial Vegetation, 1st ed.; Barbour, M.G., Billings, W.D., Eds.; Cambridge University Press: New York, NY, USA, 1988; ISBN 13-978-0-52-126198-2. [Google Scholar]
- Quiñonez-Piñón, M.R.; Valeo, C. Assessing the translucence and color-change methods for estimating sapwood depth in three boreal species. Forests 2018, 9, 686. [Google Scholar] [CrossRef]
- Kramer, P.J.; Kozlowski, T.T. Physiology of Woody Plants, 1st ed.; Academic Press Inc.: New York, NY, USA, 1979. [Google Scholar]
- Payton, M.E.; Miller, A.E.; Raun, W.R. Testing statistical hypothesis using standard error bars and confidence intervals. Commun. Soil Sci. Plant Anal. 2000, 31, 547–551. [Google Scholar] [CrossRef]
- Vangel, M.G. Confidence intervals for a normal coefficient of variation. Am. Stat. 1996, 50, 21–26. [Google Scholar]
- Verrill, S. Confidence Bounds for Normal and Log Normal Distribution Coefficients of Variation; Research Paper FPL-RP-609; The Forest Products Laboratory: Madison, WI, USA, 2003; pp. 1–10.
- Payton, M.E. Confidence intervals for the coefficient of variation. In Refereed Proceedings for the 1996 Kansas State University Conference for Applied Statistics in Agriculture; Kansas State University: Manhattan, KS, USA, 1997; pp. 82–87. [Google Scholar]
- Chen, J.M.; Cihlar, J. Retrieving leaf area index of boreal conifer forests using Landsat-TM Images. Remote Sens. Environ. 1996, 55, 153–162. [Google Scholar] [CrossRef]
- Leblanc, S.G.; Chen, J.M.; Kwong, M. Tracing Radiation and Architecture of Canopies. TRAC Manual Version 2.1.3., 1st ed.; Natural Resources Canada, Canada Centre for Remote Sensing: Ottawa, ON, Canada, 2002.
- Chen, J.M.; Rich, P.M.; Gower, S.T.; Norman, J.M.; Plummer, S. Leaf area index of boreal forests: Theory, techniques, and measurements. J. Geophys. Res. 1997, 102, 429–443. [Google Scholar] [CrossRef]
- Robinson, D.E.; Wagner, R.G.; Swanton, C.J. Effects of nitrogen on the growth of Jack pine competing with Canada blue-joint grass and large leaved aster. For. Ecol. Manag. 2002, 160, 233–242. [Google Scholar] [CrossRef]
- Chapra, S.C.; Canale, R.P. Numerical Methods for Engineers, 2nd ed.; McGraw Hill, Inc.: New York, NY, USA, 1988; p. 833. ISBN 13-9780079099440. [Google Scholar]
- Sperry, J.; Tyree, M.T. Water-stress-induced xylem embolism in three species of conifers. Plant Cell Environ. 1989, 13, 427–436. [Google Scholar] [CrossRef]
- Xiao, C.W.; Janssens, J.; Curiel, Y.; Ceulemans, R. Variation of specific leaf area and upscaling to leaf area index in mature Scots pine. Trees 2006, 20, 304. [Google Scholar] [CrossRef]
- Scheinder, R.; Berninger, F.; Ung, C.; Bernier, P.Y.; Swift, D.E.; Zhang, S.Y. Calibration of jack pine allometric relationships with simultaneous regressions. Can. J. For. Res. 2008, 38, 2566. [Google Scholar] [CrossRef]
- Thurner, M.; Beer, C.; Crowther, T.; Falster, D.; Manzoni, S.; Prokushkin, A.; Schulze, E.D. Supporting Information for Sapwood Biomass Carbon in Northern Boreal and Temperate Forests. 2019. Available online: https://pure.mpg.de › item_3065977_1 › component › file_3065979 › content (accessed on 16 August 2019).
- Shinohara, Y.; Komatsu, H.; Otsuki, K. Sapwood and intermediate wood variations in Japanese cedar: Impacts on sapwood area estimates. Hydrol. Res. Lett. 2015, 9, 35. [Google Scholar] [CrossRef]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists, 1st ed.; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Hernandez-Santana, V.; Hernandez-Hernandez, A.; Vadenboncoeur, M.A.; Asbjornsen, H. Scaling from single-point sap velocity measurements to stand transpiration in a multispecies deciduous forest: Uncertainty sources, stand structure effect, and future scenarios. Can. J. For. Res. 2015, 45, 1489. [Google Scholar] [CrossRef]
- Bell, D.M.; Ward, E.J.; Oishi, C.; Oren, R.; Flikkema, P.G.; Clark, J.S. A state-space modeling approach to estimating the canopy conductance and associated uncertainties from sap flux density data. Tree Physiol. 2015, 35, 792–802. [Google Scholar] [CrossRef]
- Mencuccini, M.; Bonosi, L. Leaf/sapwood area ratios in Scots pine show acclimation across Europe. Can. J. For. Res. 2001, 31, 442. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species (k) | (cm2) | |
|---|---|---|
| Pinus banksiana and Pinus contorta | 33 | 176.47 |
| Picea mariana | 21 | 213.57 |
| Populus tremuloides | 26 | 277.13 |
| Picea glauca | 17 | 277.52 |
| Plot | Tree species | No. of Trees | Max. DOB | Min. DOB | Avg. DOB | SAsp | SAplot | ∆SAplot |
|---|---|---|---|---|---|---|---|---|
| Conifer-4 1 | Picea glauca | 434 | 43.3 | 2.2 | 13.14 | 2.92 | ||
| Pinus contorta | 276 | 33.42 | 5.73 | 20.15 | 4.87 | 7.79 | ± 0.03 | |
| Conifer-5 1 | Picea glauca | 164 | 48.38 | 2.86 | 18.31 | 2.29 | ||
| Pinus contorta | 48 | 26.1 | 1.59 | 14.09 | 0.85 | |||
| Populus tremuloides | 114 | 19.42 | 2.86 | 7.23 | 0.14 | 3.28 | ± 0.03 | |
| Conifer-11 2 | Pinus banksiana | 129 | 25.15 | 6.37 | 16.57 | 2.28 | 2.28 | ± 0.03 |
| Conifer-12 2 | Picea mariana | 60 | 23.55 | 5.09 | 15.69 | 1.28 | 1.28 | ± 0.01 |
| Deciduous-1 1 | Populus tremuloides | 83 | 25.46 | 9.55 | 18.57 | 1.45 | 1.45 | ± 0.01 |
| Deciduous-6 1 | Populus tremuloides | 498 | 31.19 | 8.91 | 19.1 | 9.37 | ||
| Picea glauca | 14 | 48.38 | 7 | 23.4 | 0.18 | 9.55 | ± 0.08 |
| Plot | Tree Species | No. of Trees | Max. DOB | Min. DOB | Avg. DOB | SAsp | SAplot | ∆SAplot |
|---|---|---|---|---|---|---|---|---|
| Conifer-1’ | Picea glauca | 29 | 24.51 | 5.091 | 6.53 | 0.16 | ||
| Pinus contorta | 12 | 26.74 | 14.01 | 19.89 | 0.21 | 0.37 | ± 0.001 | |
| Conifer-2’ | Pice glauca | 15 | 27.06 | 7.641 | 14.88 | 0.21 | ||
| Pinus contorta | 9 | 27.37 | 16.23 | 20.23 | 0.16 | 0.37 | ± 0.001 | |
| Conifer-3’ | Picea glauca | 19 | 25.78 | 4.461 | 12.48 | 0.19 | ||
| Pinus contorta | 13 | 29.6 | 10.82 | 19.98 | 0.23 | 0.42 | ± 0.0007 | |
| Conifer-4’ | Picea glauca | 9 | 21.33 | 3.501 | 12.59 | 0.04 | ||
| Pinus contorta | 14 | 27.06 | 11.78 | 20.35 | 0.25 | 0.29 | ± 0.0003 | |
| Conifer-5’ | Picea glauca | 4 | 28.33 | 12.41 | 18.94 | 0.05 | ||
| Pinus contorta | 13 | 32.79 | 15.92 | 22.33 | 0.23 | 0.28 | ± 0.0002 | |
| Conifer-10’ | Picea glauca | 5 | 33.74 | 5.091 | 16.87 | 0.19 | ||
| Pinus contorta | 14 | 34.7 | 12.73 | 21.99 | 0.11 | |||
| Populus tremuloides | 1 | 21.01 | 0.02 | 0.32 | ± 0.0005 | |||
| Deciduous-7’ | Populus tremuloides | 31 | 32.79 | 9.871 | 17.15 | 0.34 | 0.34 | ± 0.004 |
| Deciduous-8’ | Populus tremuloides | 28 | 28.01 | 14.01 | 20.29 | 0.59 | 0.59 | ± 0.005 |
| Deciduous-9’ | Populus tremuloides | 22 | 23.24 | 14.64 | 19.92 | 0.43 | 0.43 | ± 0.004 |
| Site | LAI | Plot Size (m2) | LAplot (m2) | ∆LAplot (m2) |
|---|---|---|---|---|
| Conifer-1’ | 6.90 | 150 | 1035.00 | ± 19.90 |
| Conifer-2’ | 6.04 | 150 | 906 | ± 19.04 |
| Conifer-3’ | 6.57 | 150 | 985 | ± 19.57 |
| Conifer-4’ | 5.34 | 150 | 801 | ± 18.34 |
| Conifer-5’ | 4.51 | 150 | 676.5 | ± 17.51 |
| Conifer-10’ | 6.12 | 150 | 918 | ± 19.12 |
| Conifer-4 | 5.71 | 3600.00 | 20,556.00 | ± 1338.20 |
| Conifer-5 | 2.54 | 3600.00 | 9144.00 | ± 855.10 |
| Conifer-11 | 3.76 | 400 | 1502.52 | ± 259.32 |
| Conifer-12 | 4.96 | 300 | 1486.97 | ± 242.82 |
| Deciduous-1 | 2.64 | 415 | 1093.53 | ± 405.85 |
| Deciduous-6 | 3.14 | 3600,00 | 11,304.00 | ± 1144.27 |
| Deciduous-7’ | 2.30 | 100 | 230 | ± 12.30 |
| Deciduous-8’ | 3.57 | 100 | 357 | ± 13.57 |
| Deciduous-9’ | 3.22 | 100 | 322 | ± 13.22 |
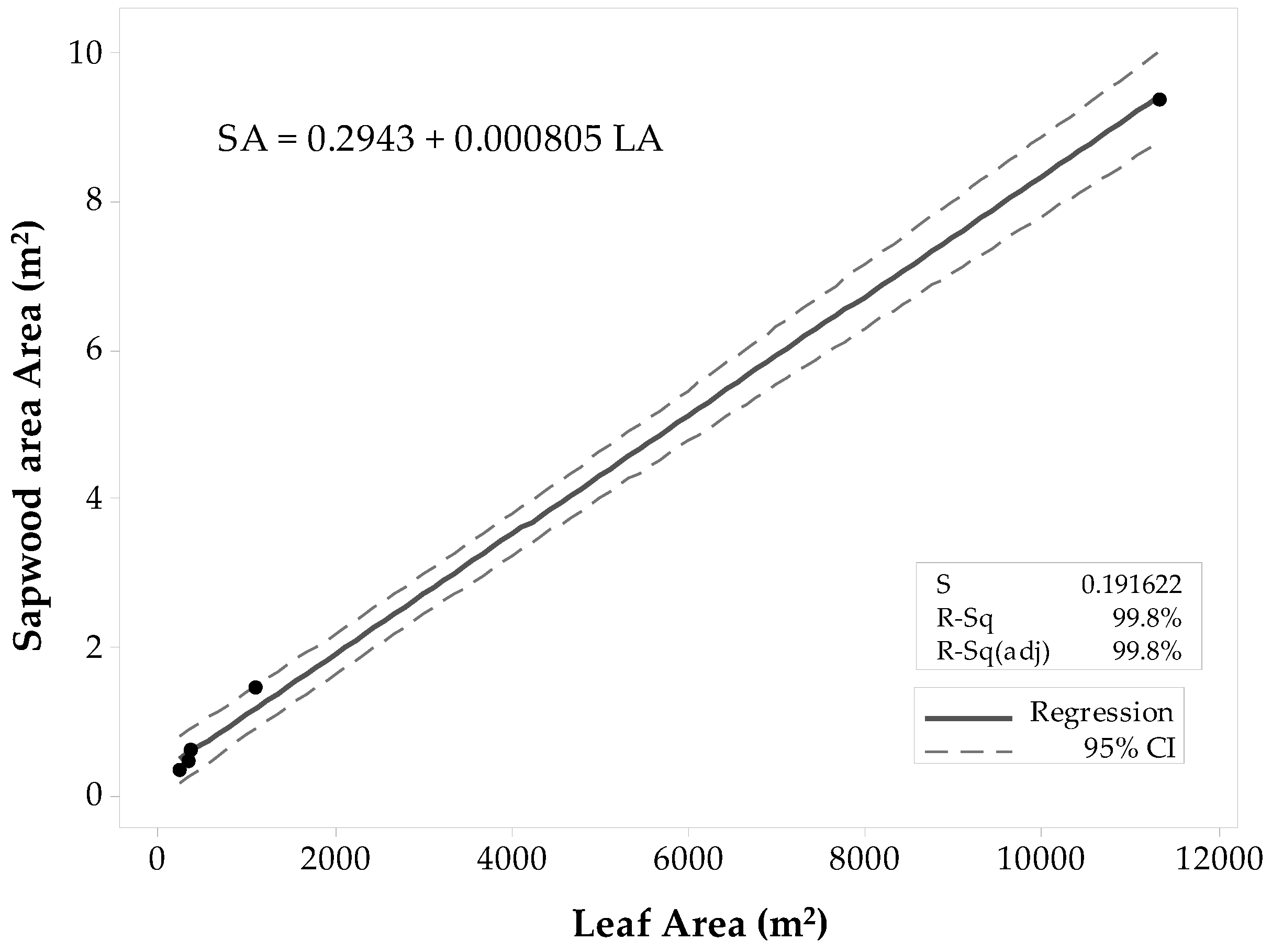
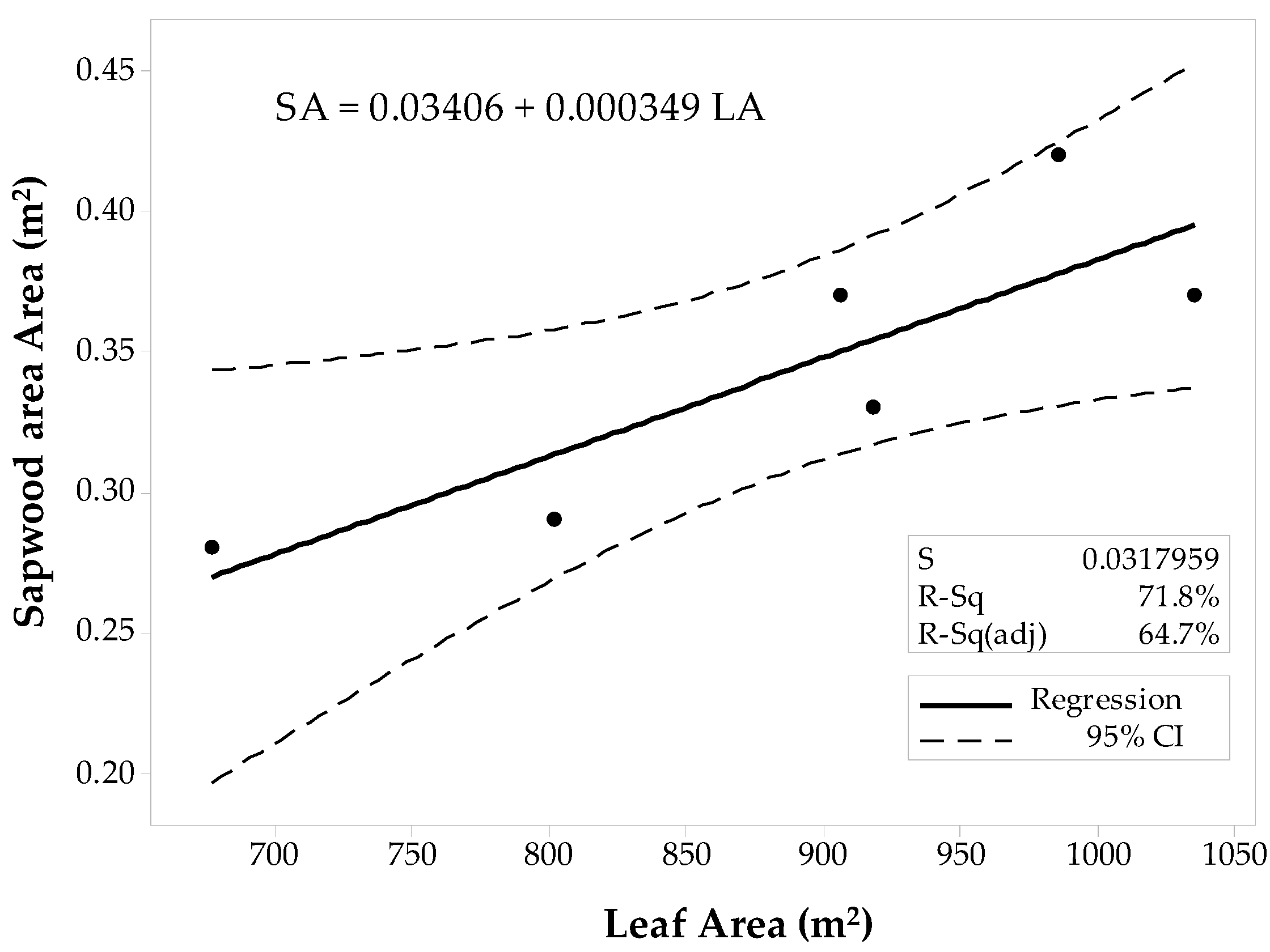
| Site | SAplot (m2) | LAplot (m2) | Pearson’s Correlation Coefficient, (p-Value) |
|---|---|---|---|
| Conifer-1’ | 0.37 | 1035 | |
| Conifer-2’ | 0.37 | 906 | |
| Conifer-3’ | 0.42 | 985.5 | |
| Conifer-4’ | 0.29 | 801 | |
| Conifer-5’ | 0.28 | 676.5 | |
| Conifer-10’ | 0.33 | 918 | |
| 0.85 1, (0.033) | |||
| Conifer-4 | 7.79 | 20,556.00 | |
| Conifer-5 | 3.28 | 9144.00 | |
| Conifer-11 | 2.28 | 1502.52 | |
| Conifer-12 | 1.28 | 1486.97 | |
| 0.98 2, (0.022) | |||
| 0.97 3, (<0.001) |
| Site Type | Variable | COV | 95% C.L. |
|---|---|---|---|
| Coniferous | SAplot | 0.72 | 0.4963–1.6222 |
| LAplot | 0.79 | 0.5943–1.9426 | |
| LAI | 0.32 | 0.2438–0.7976 | |
| Deciduous | SAplot | 1.46 | 0.6614–1.7885 |
| LAplot | 1.82 | 0.7026–1.8999 | |
| LAI | 0.17 | 0.1344–0.3633 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quiñonez-Piñón, M.R.; Valeo, C. Scaling Approach for Estimating Stand Sapwood Area from Leaf Area Index in Five Boreal species. Forests 2019, 10, 829. https://doi.org/10.3390/f10100829
Quiñonez-Piñón MR, Valeo C. Scaling Approach for Estimating Stand Sapwood Area from Leaf Area Index in Five Boreal species. Forests. 2019; 10(10):829. https://doi.org/10.3390/f10100829
Chicago/Turabian StyleQuiñonez-Piñón, M. Rebeca, and Caterina Valeo. 2019. "Scaling Approach for Estimating Stand Sapwood Area from Leaf Area Index in Five Boreal species" Forests 10, no. 10: 829. https://doi.org/10.3390/f10100829
APA StyleQuiñonez-Piñón, M. R., & Valeo, C. (2019). Scaling Approach for Estimating Stand Sapwood Area from Leaf Area Index in Five Boreal species. Forests, 10(10), 829. https://doi.org/10.3390/f10100829