Landscape-Scale Mixtures of Tree Species are More Effective than Stand-Scale Mixtures for Biodiversity of Vascular Plants, Bryophytes and Lichens

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
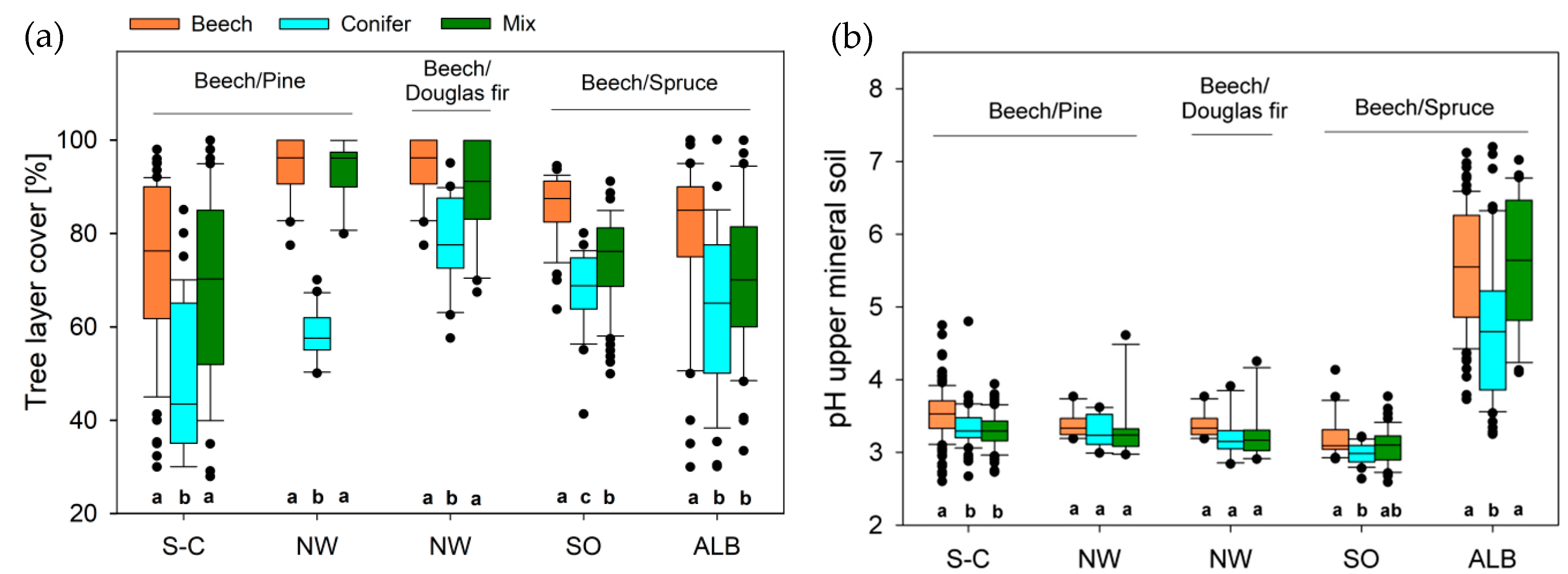
2.1. Study Regions and Data Sampling
2.2. Data Analysis
2.2.1. Analyzing Differences among Stand Types
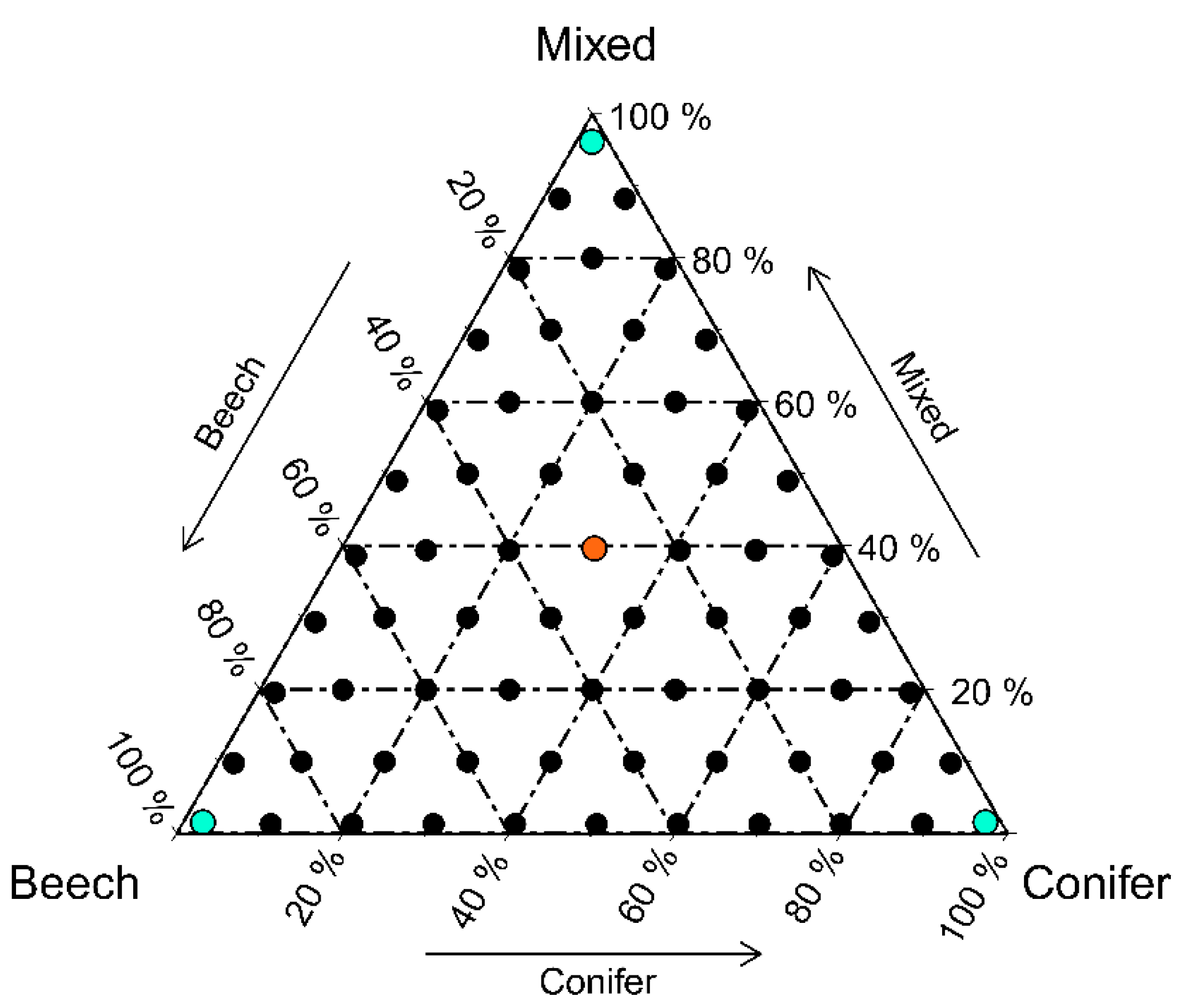
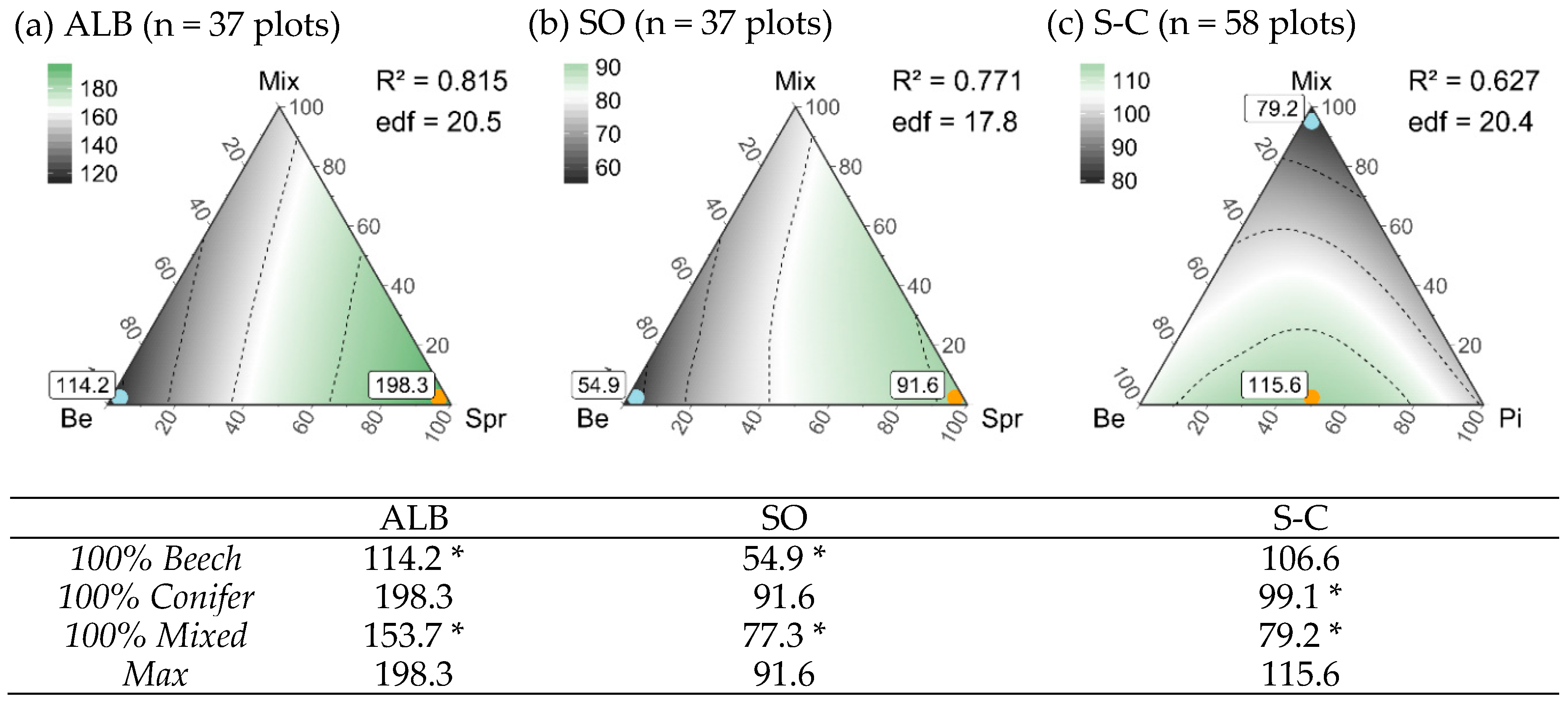
2.2.2. Finding an Optimized Composition of Stand Types for Gamma Diversity of Vascular Plants, Bryophytes and Lichens
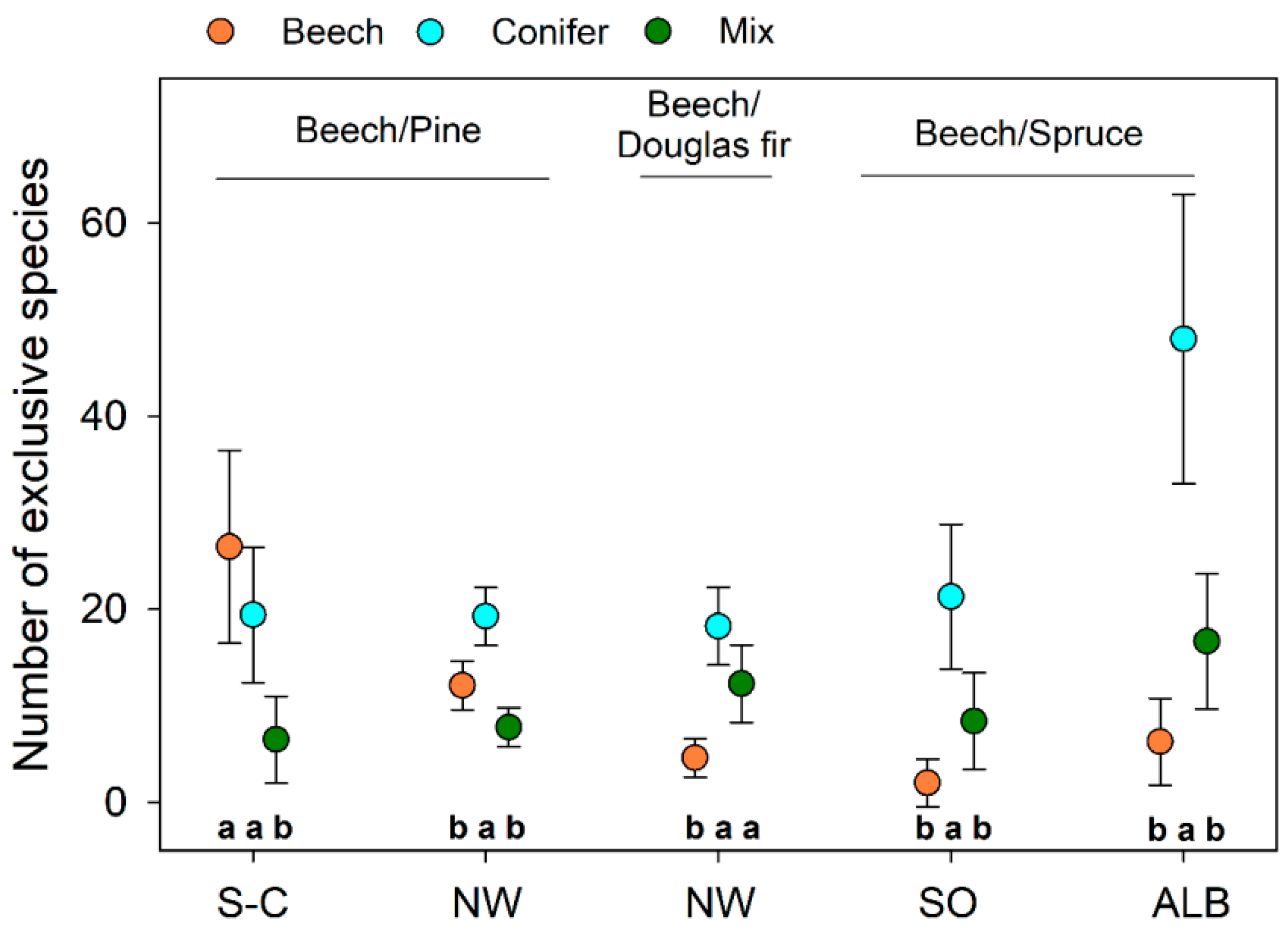
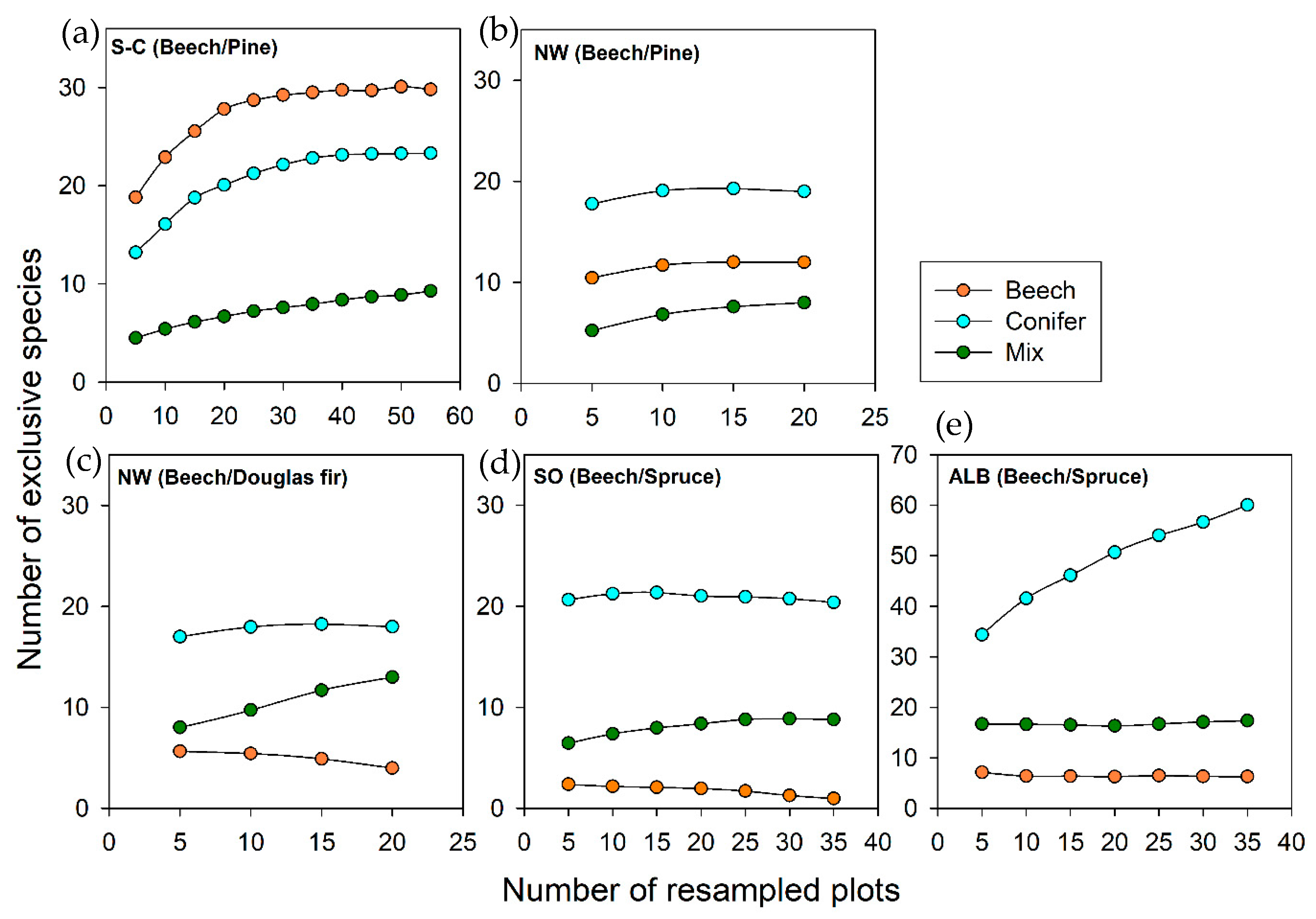
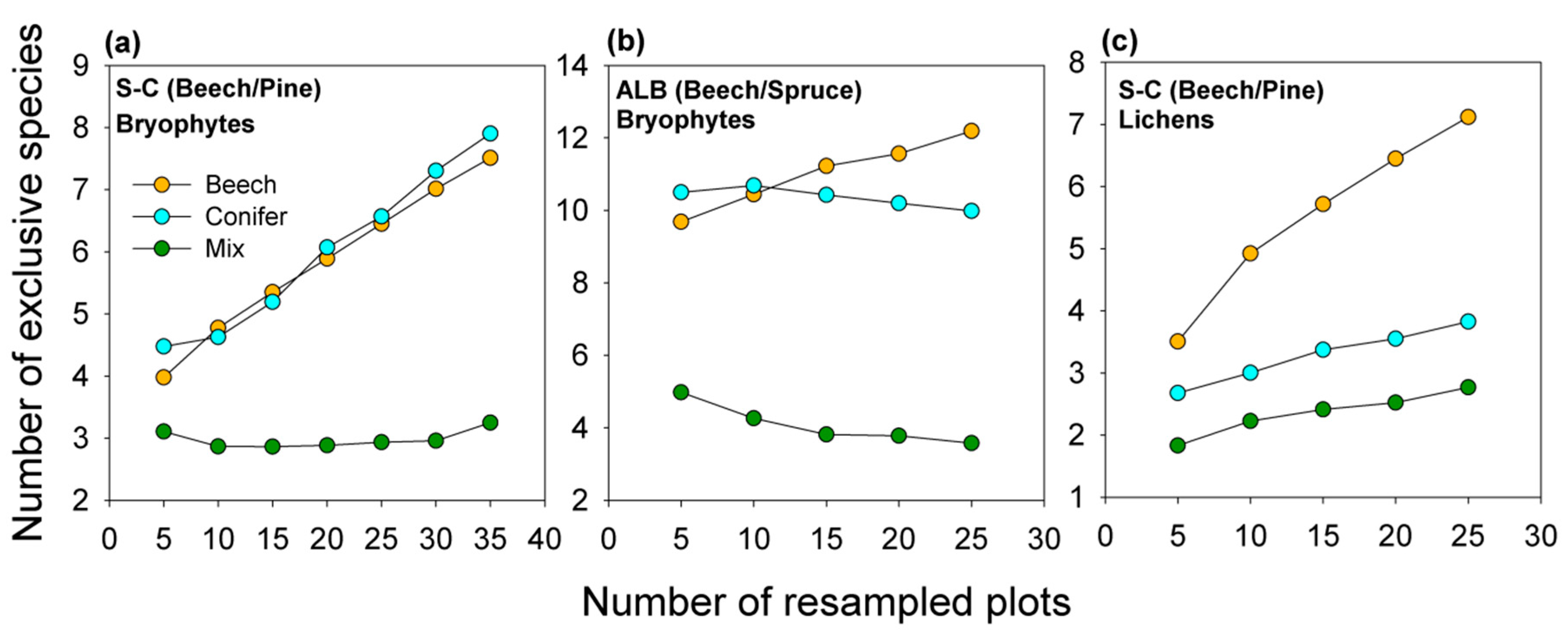
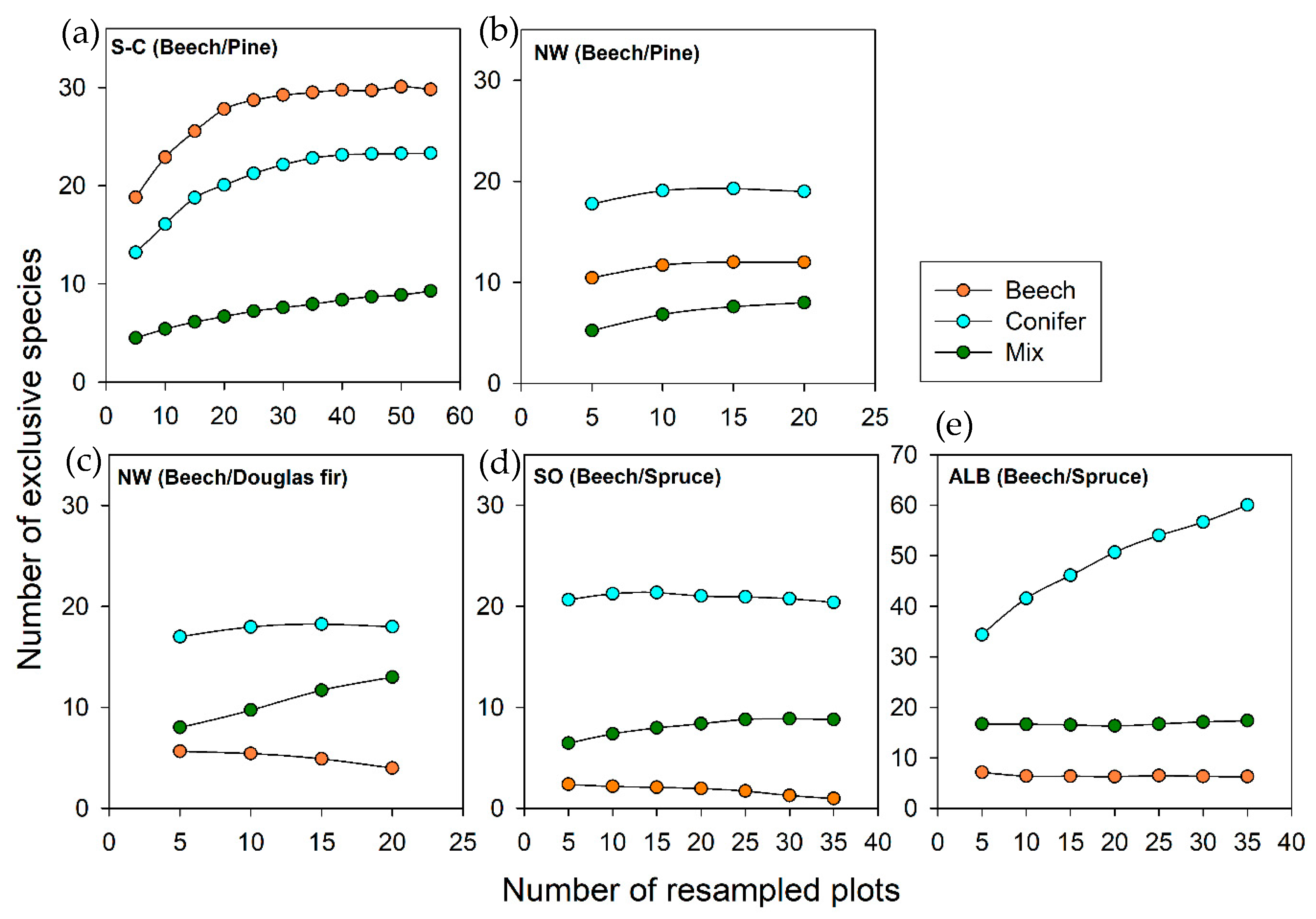
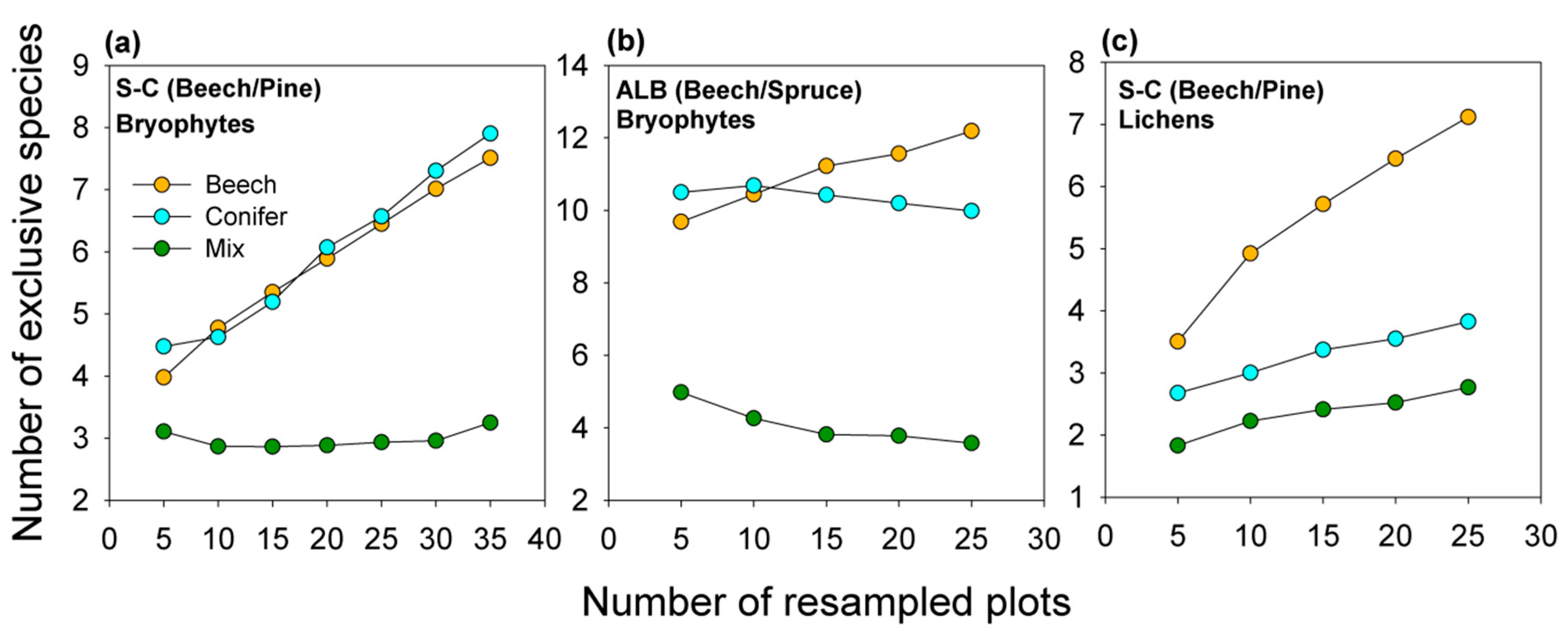
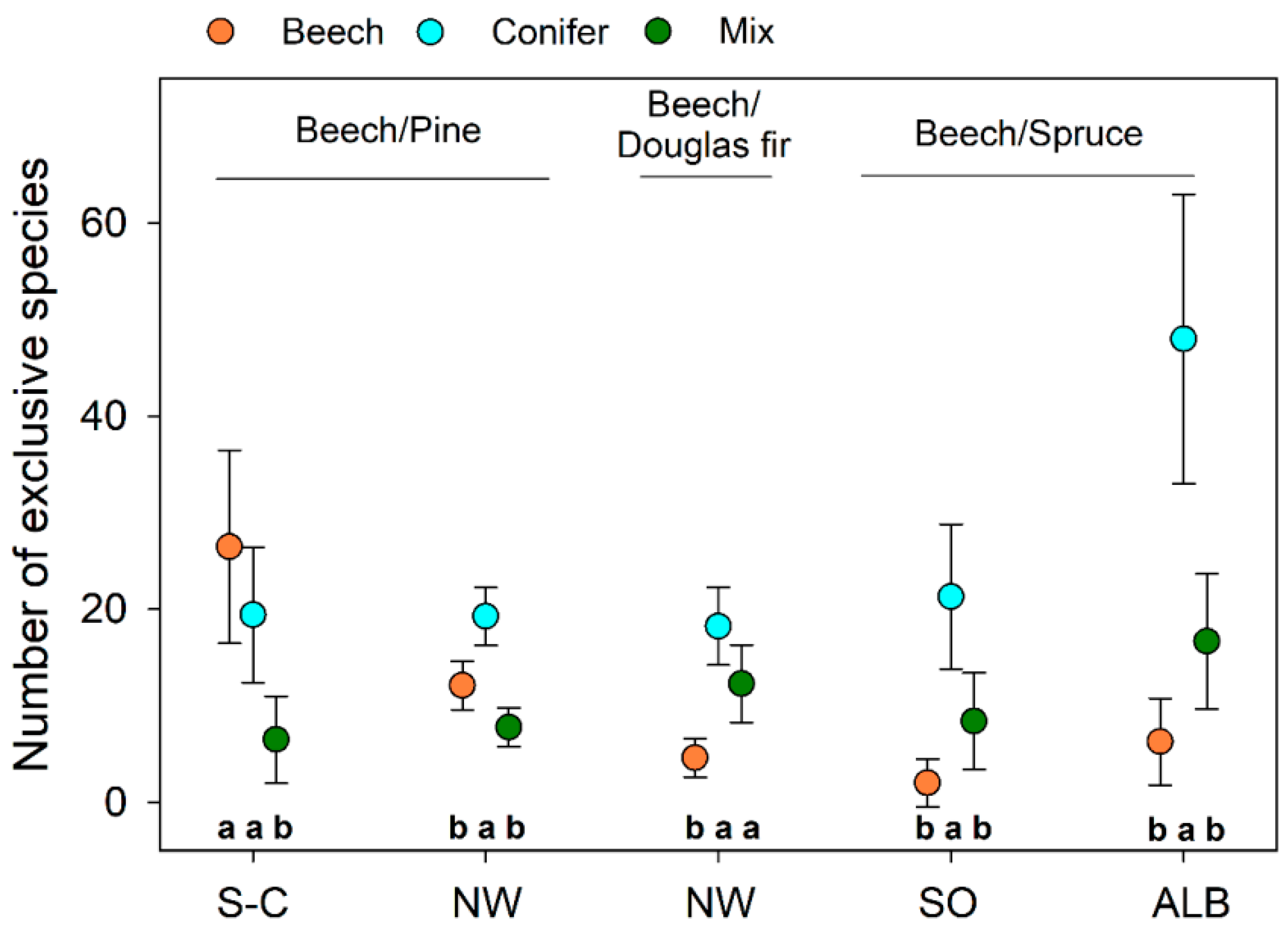
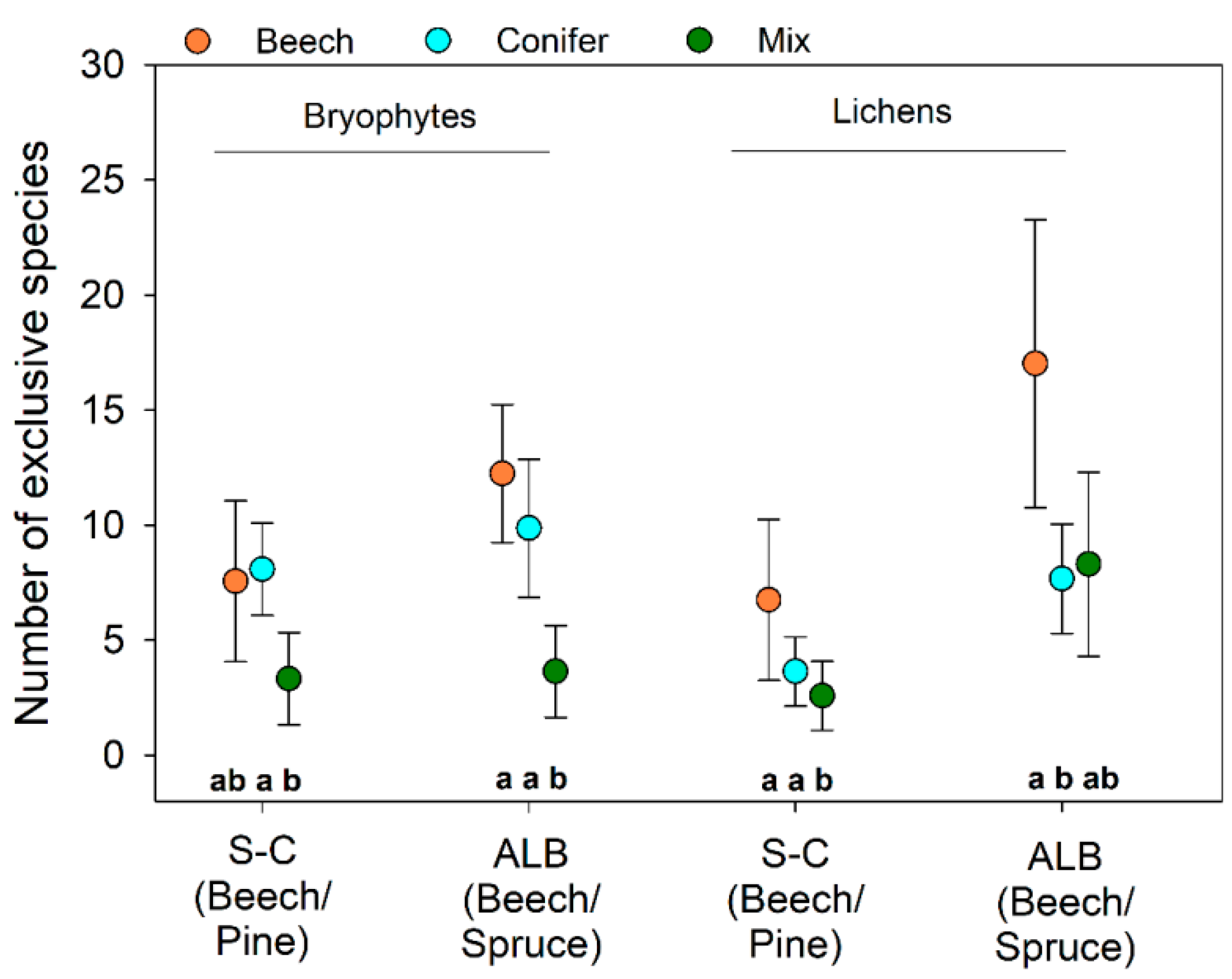
2.2.3. Quantification of Exclusive Species Numbers per Stand Type
2.2.4. Identification of Characteristic Species per Stand Type
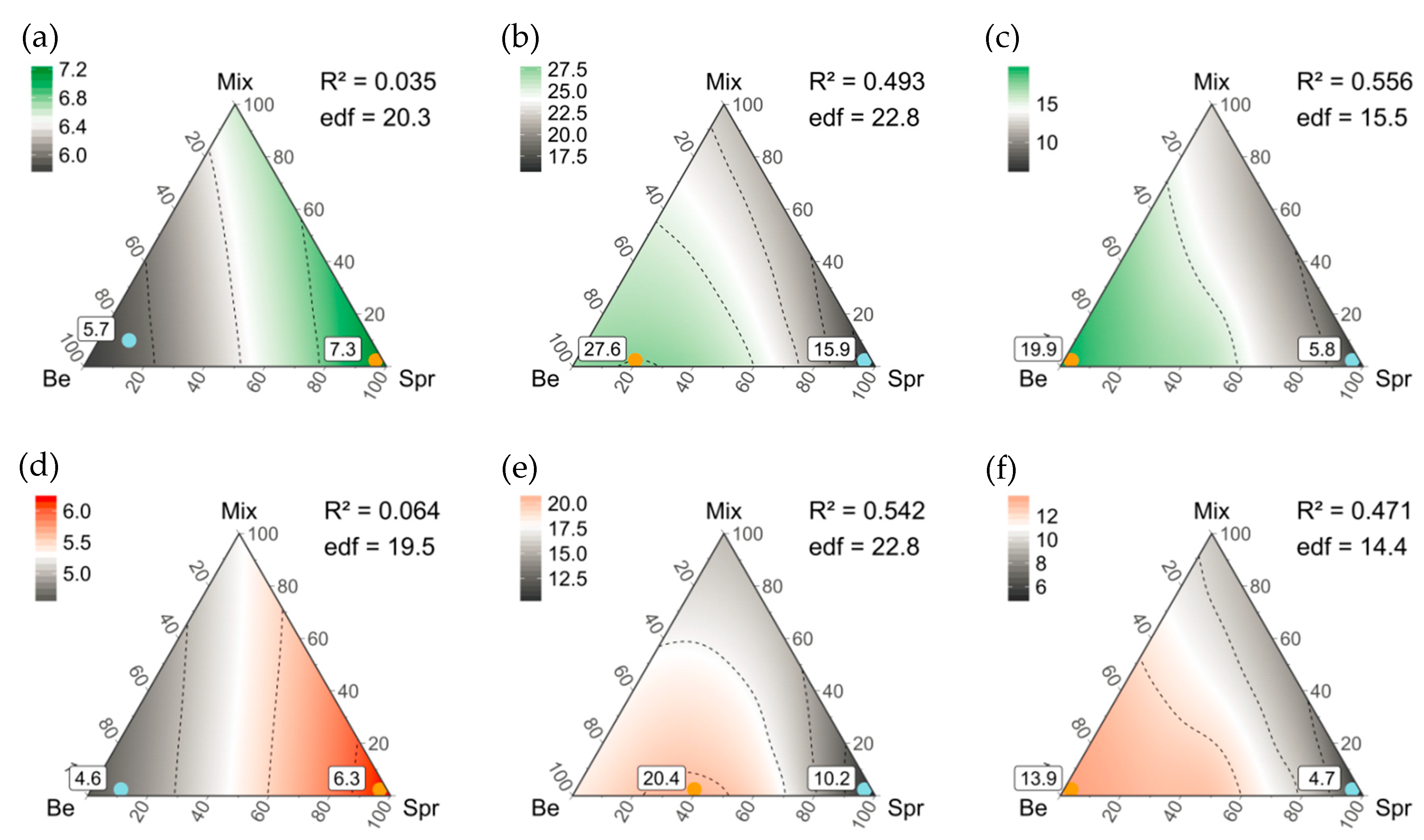
2.2.5. Finding an Optimized Composition of Stand Types for Regional Diversity
3. Results
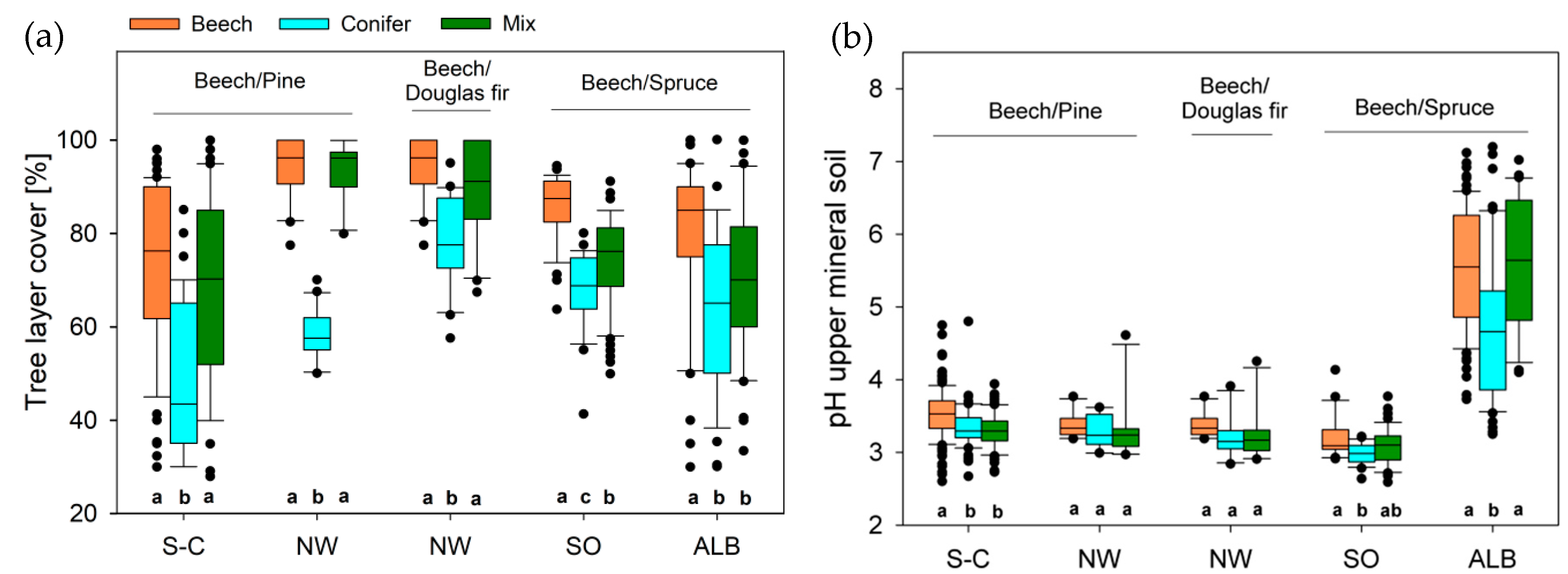
3.1. Environmental Conditions
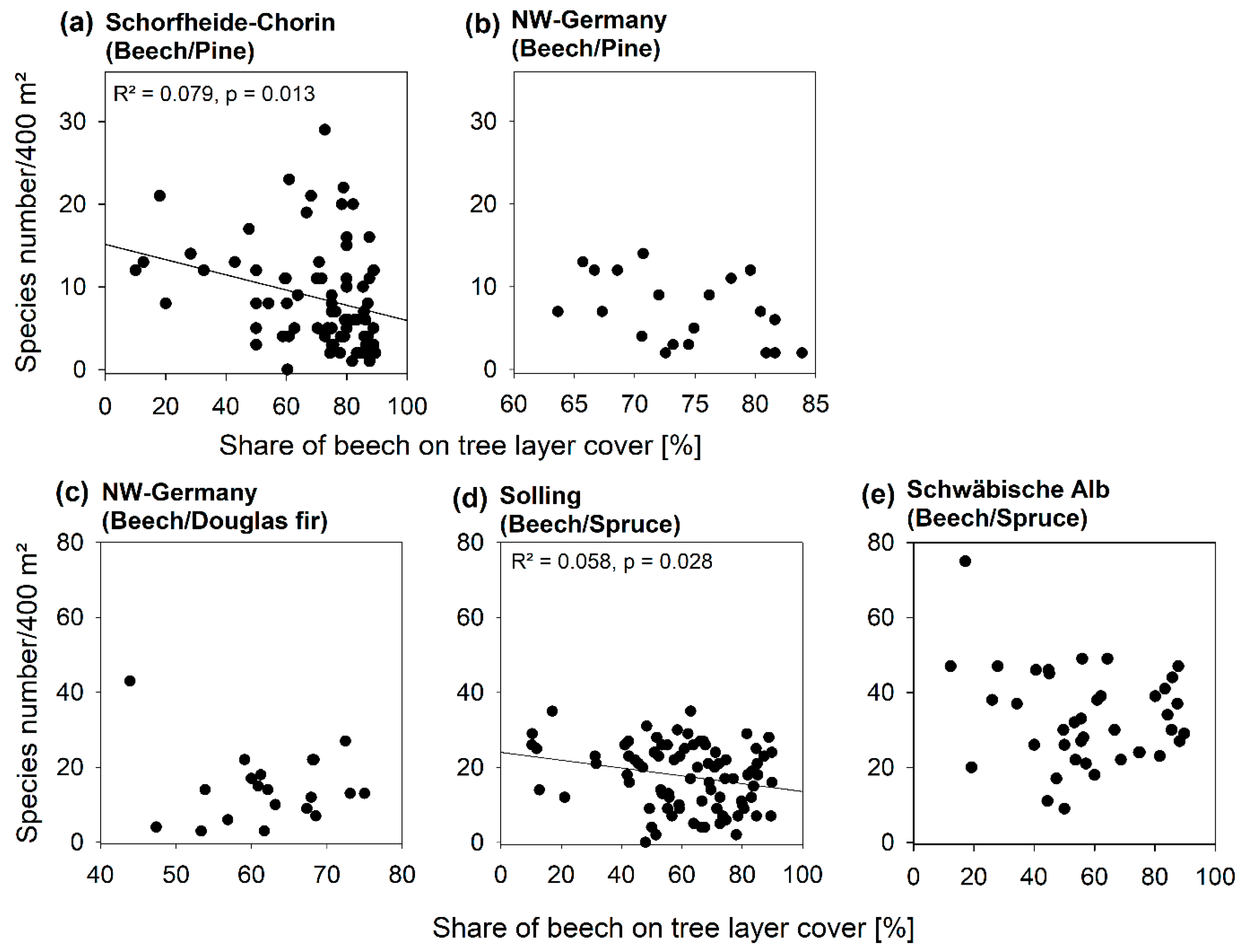
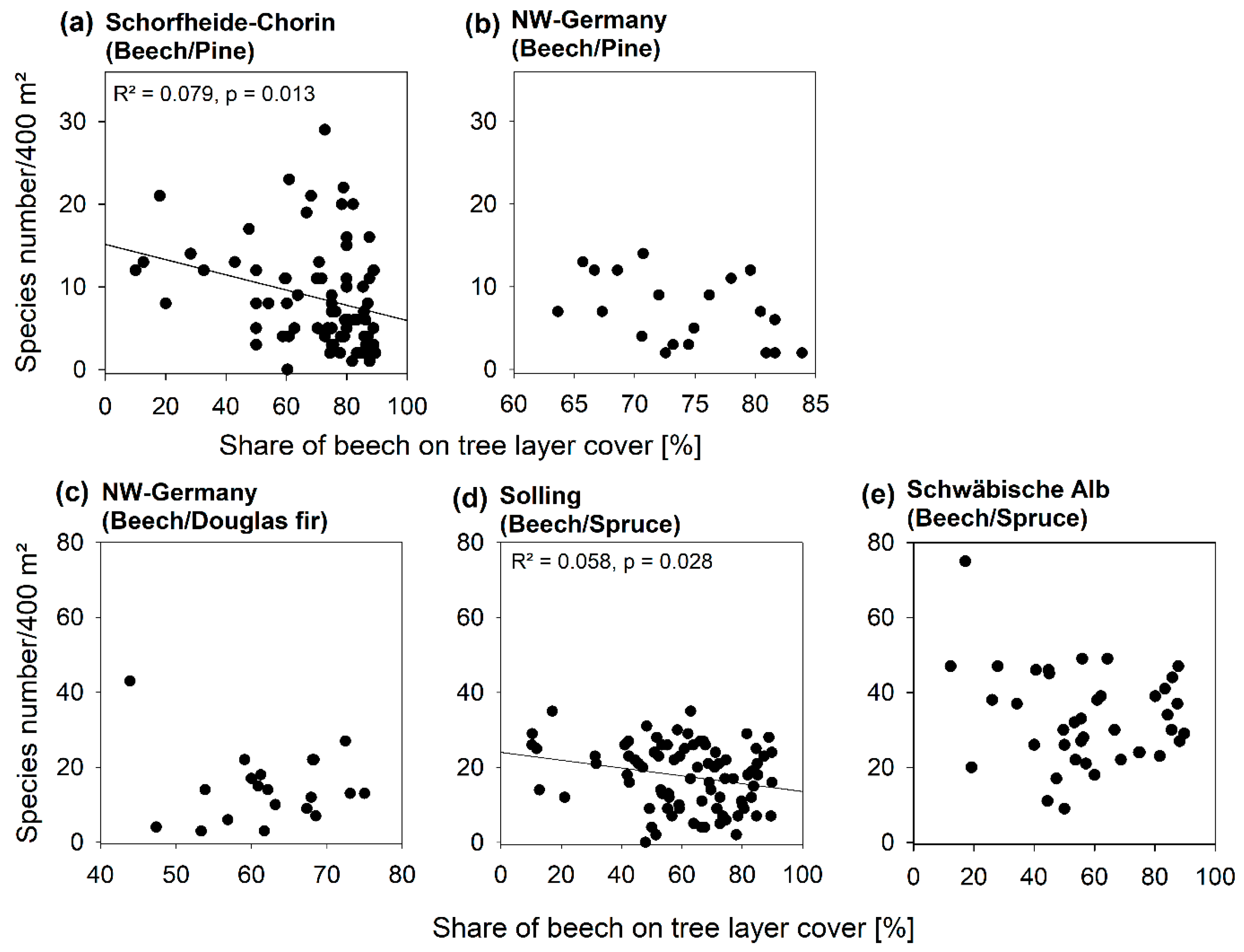
3.2. Diversity Patterns of Vascular Plants
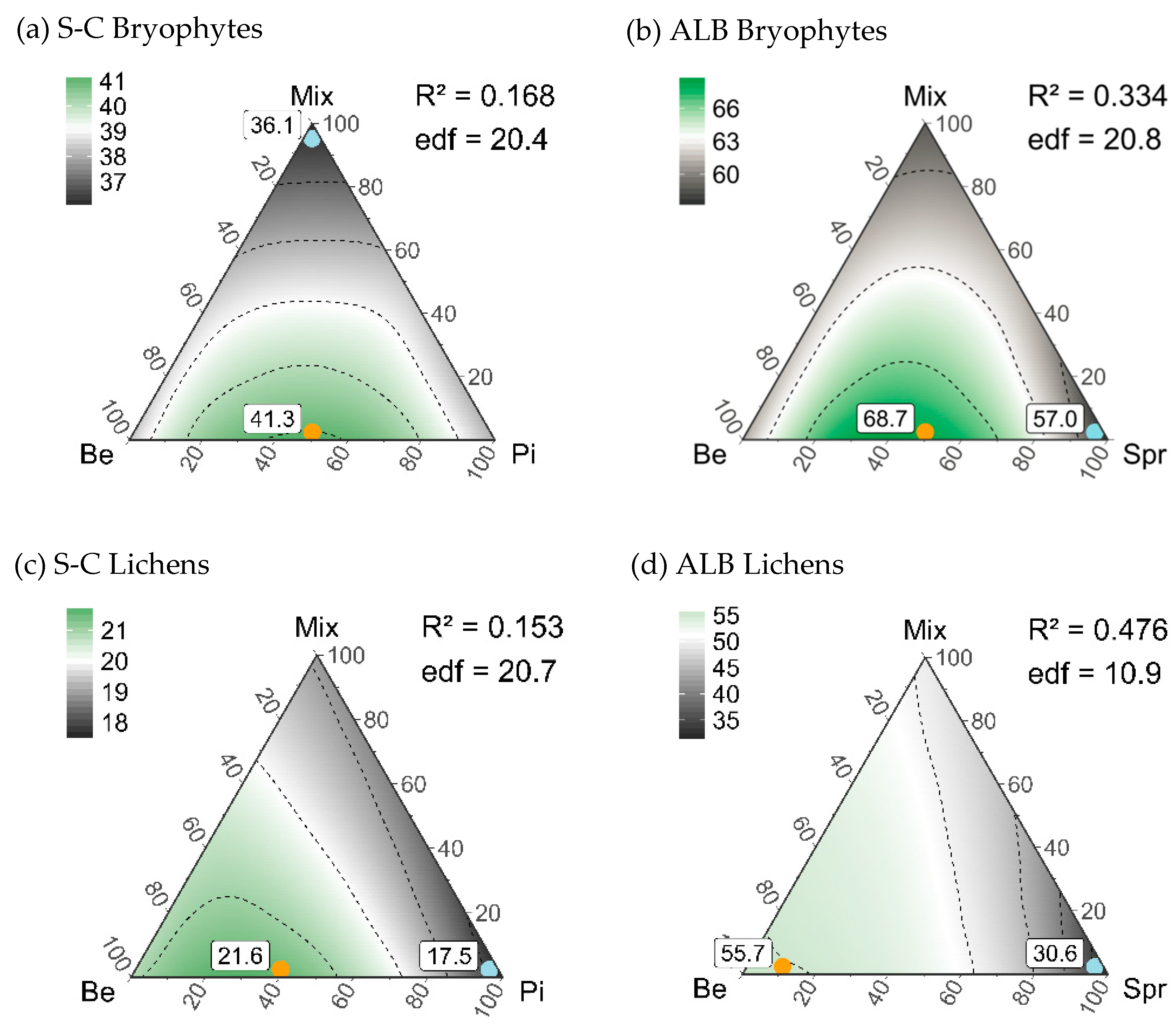
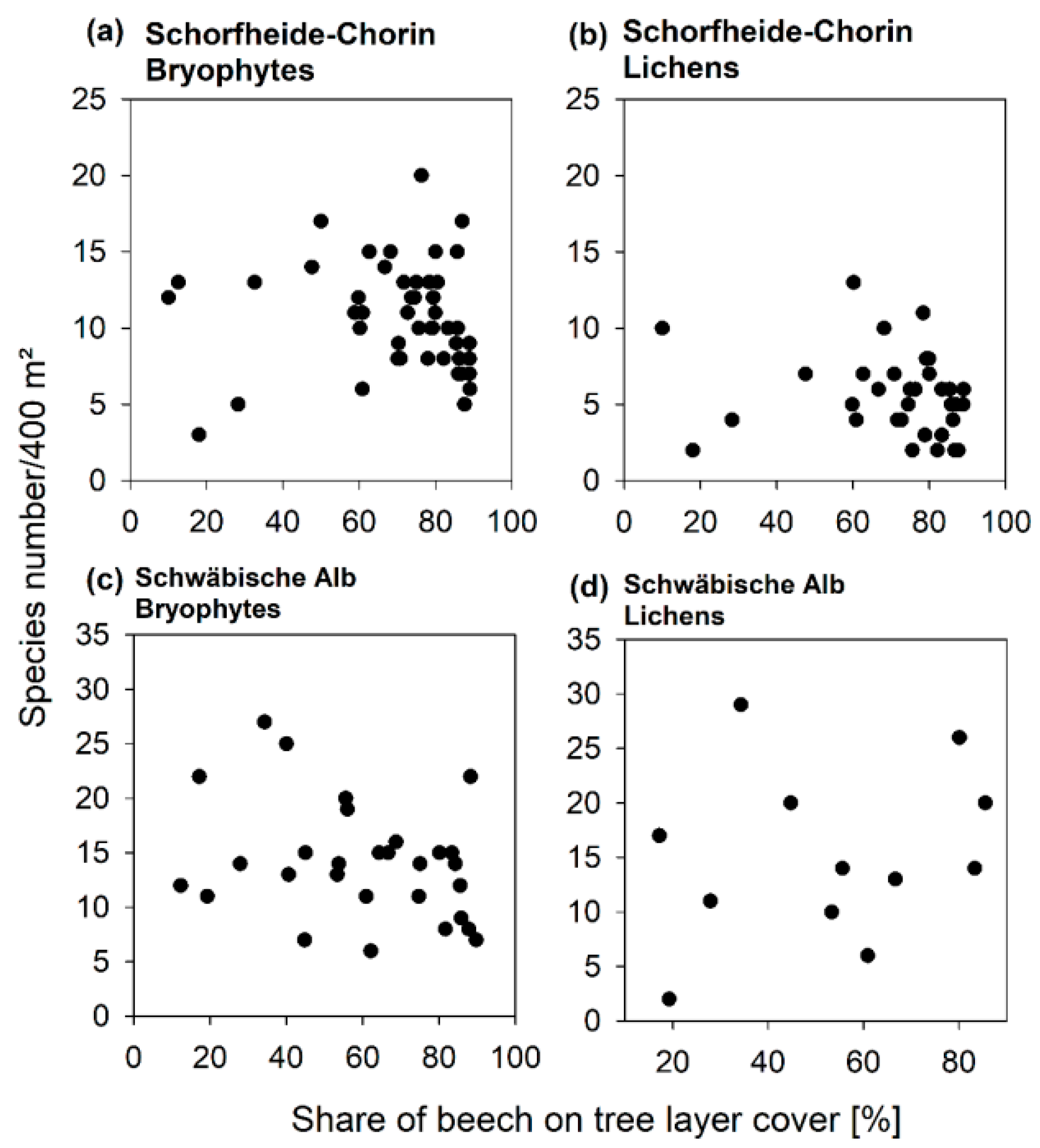
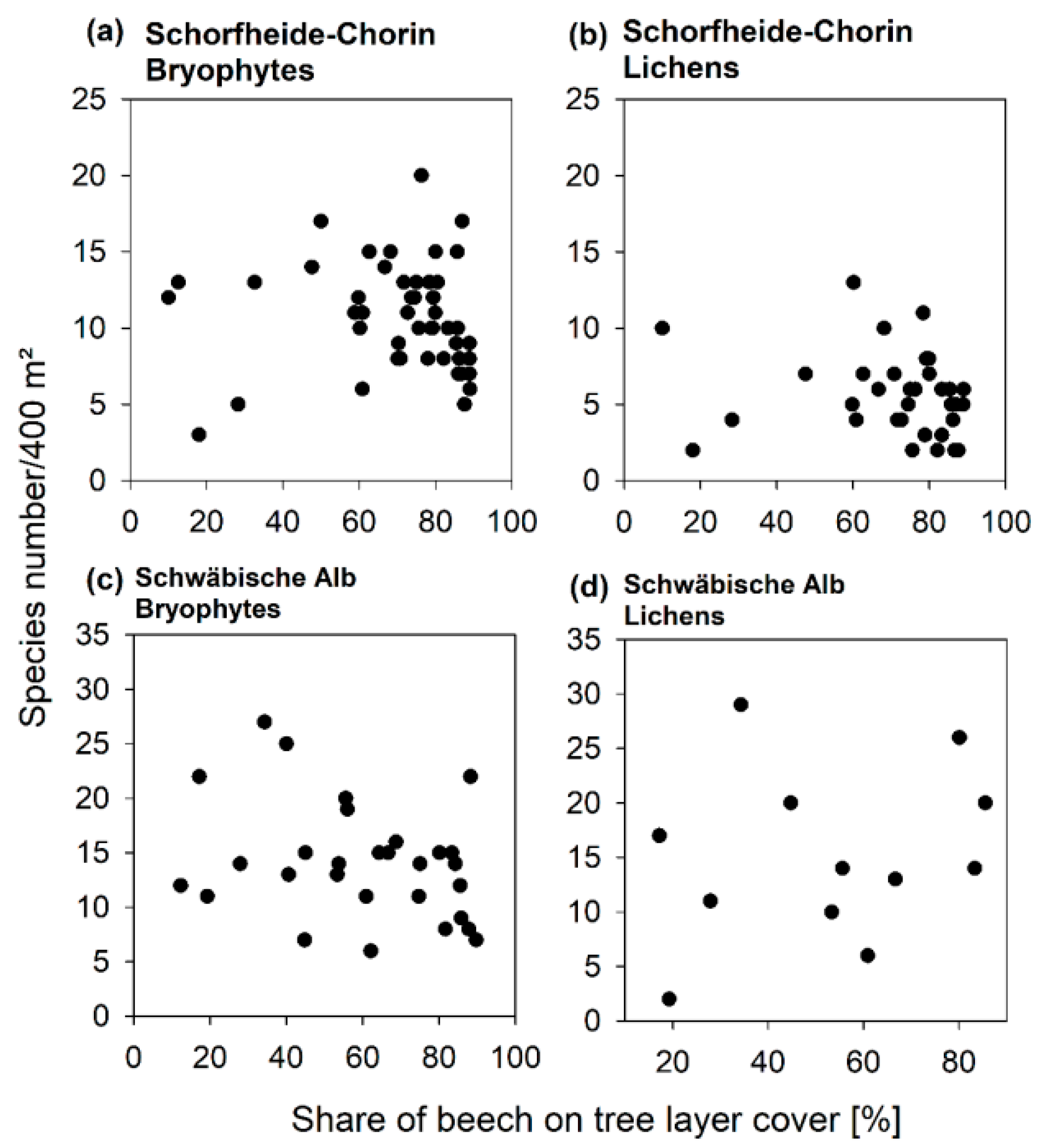
3.3. Diversity Patterns of Bryophytes and Lichens
3.4. Characteristic Species within Stand Types
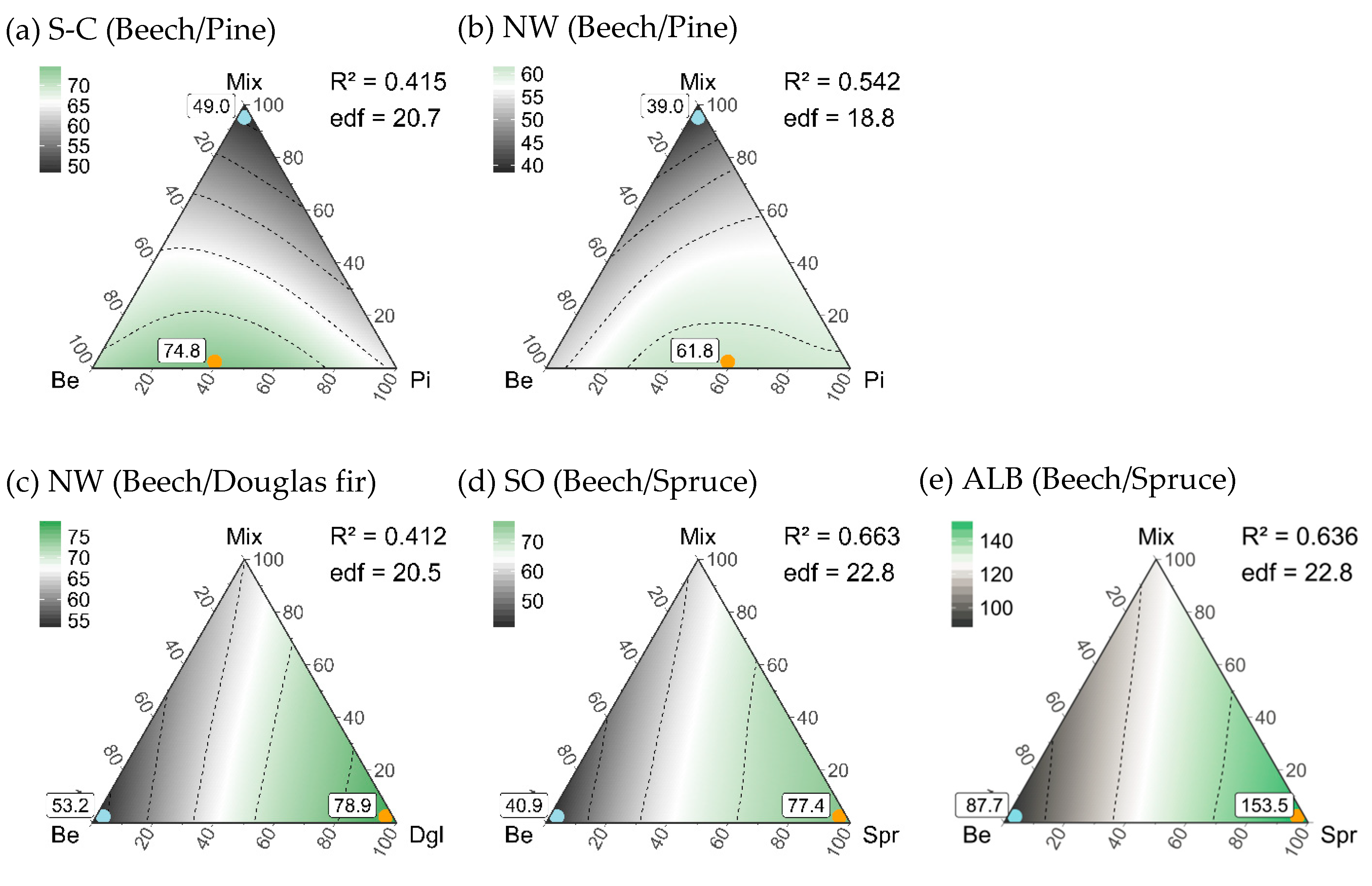
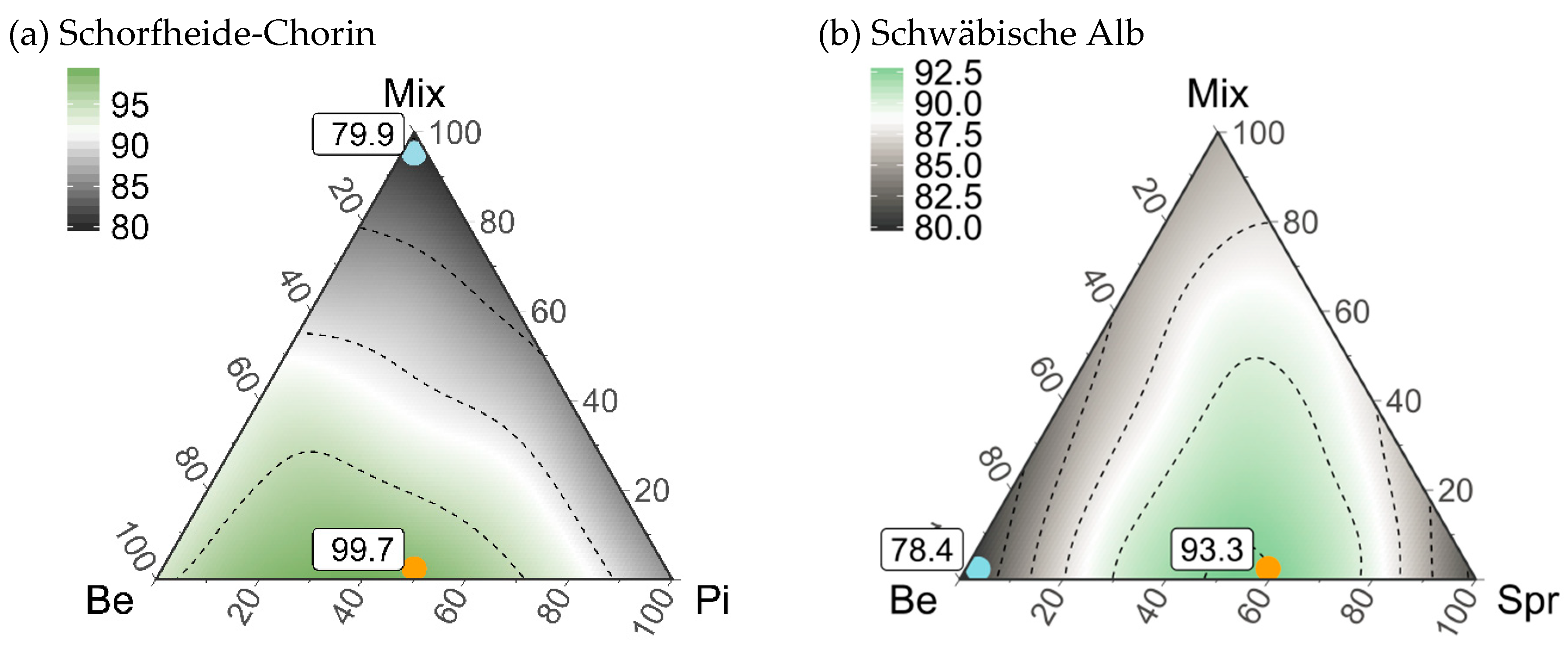
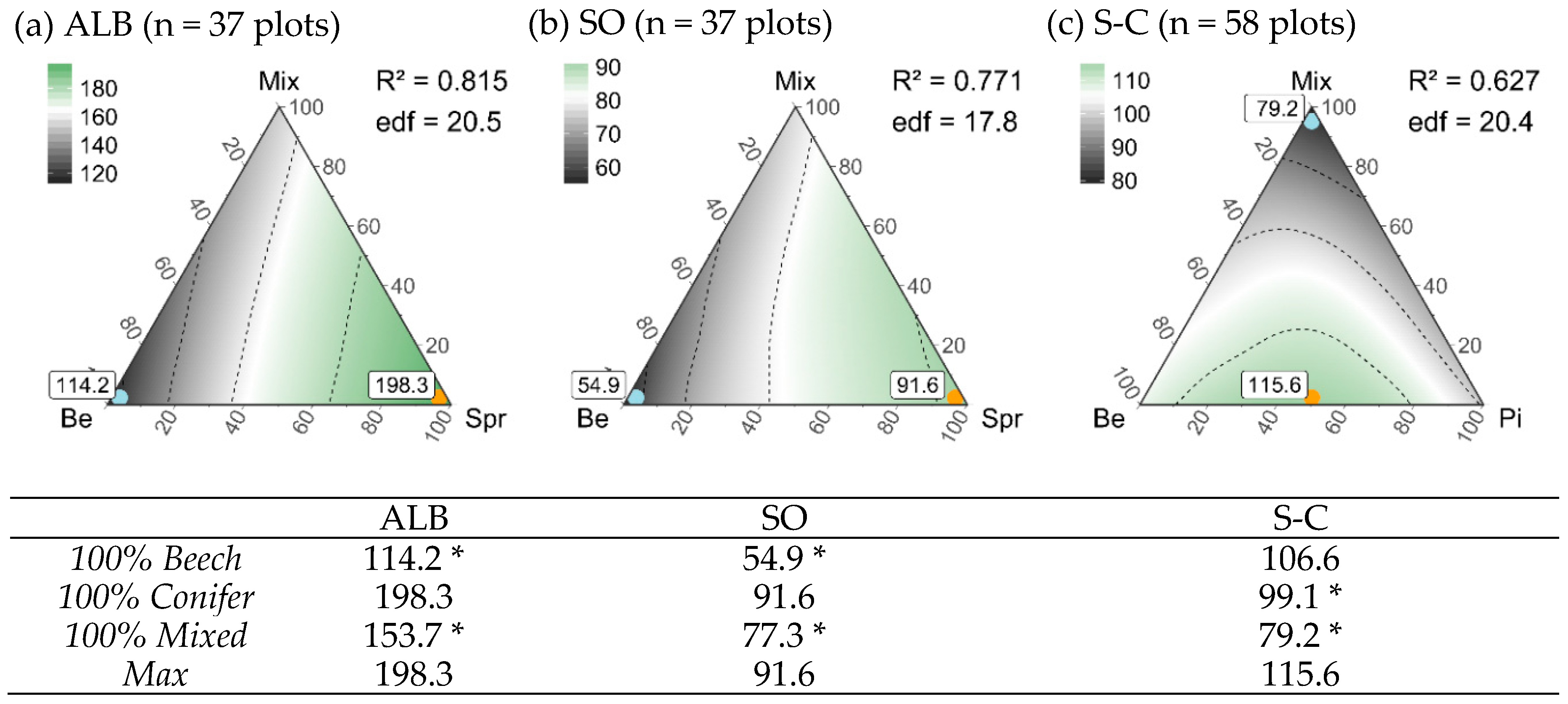
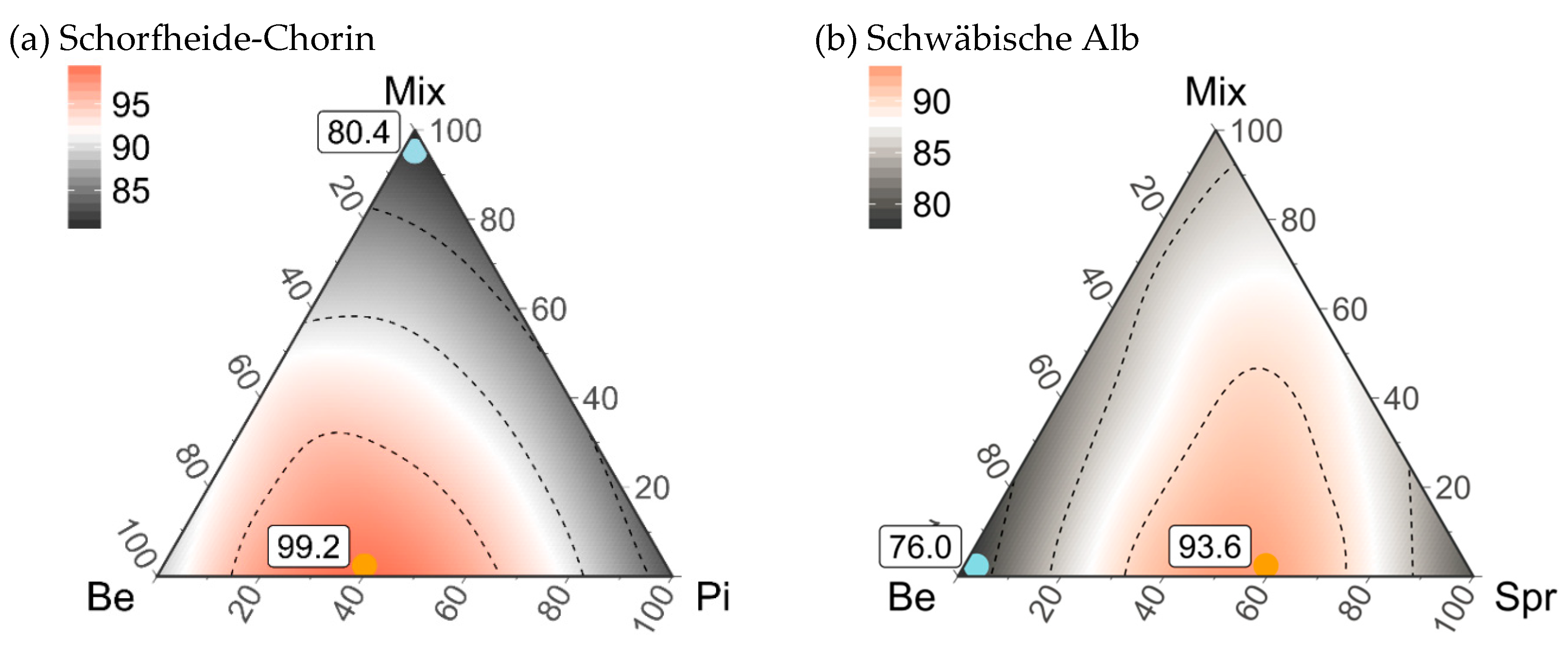
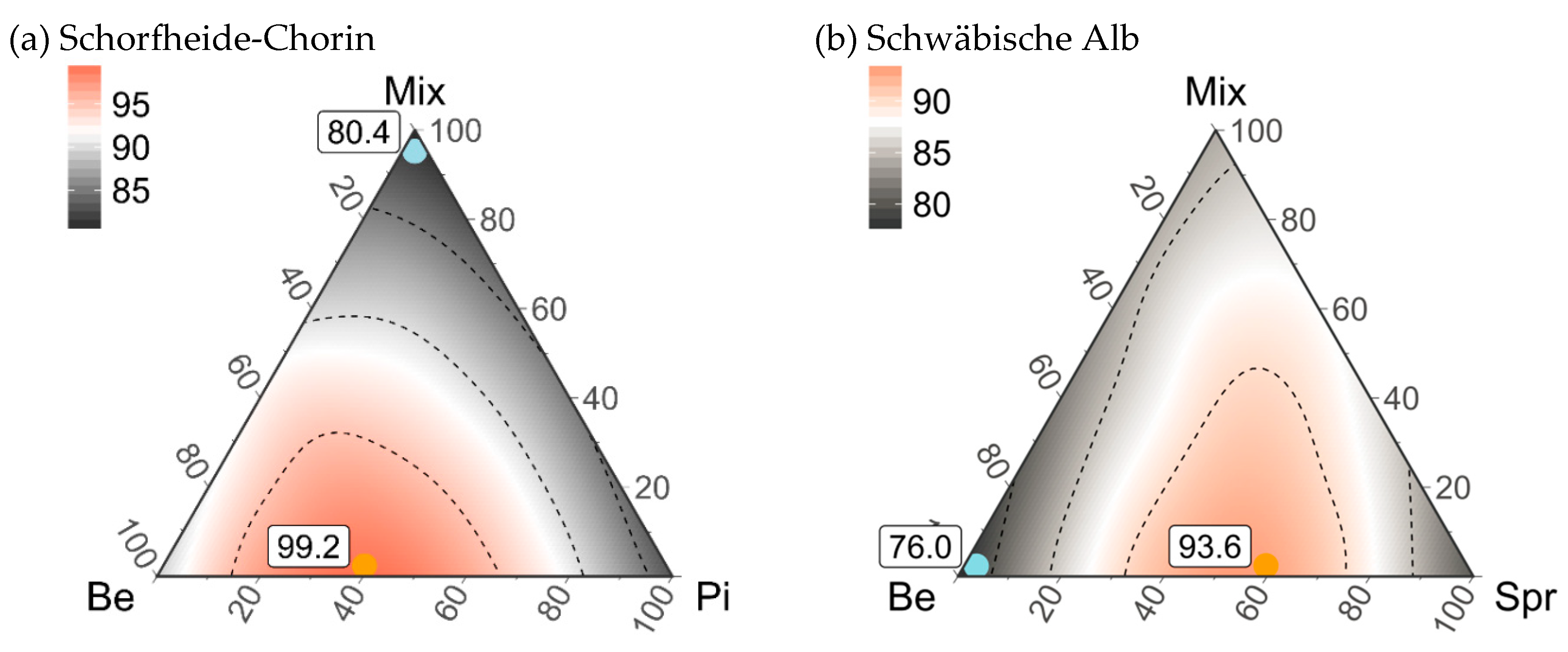
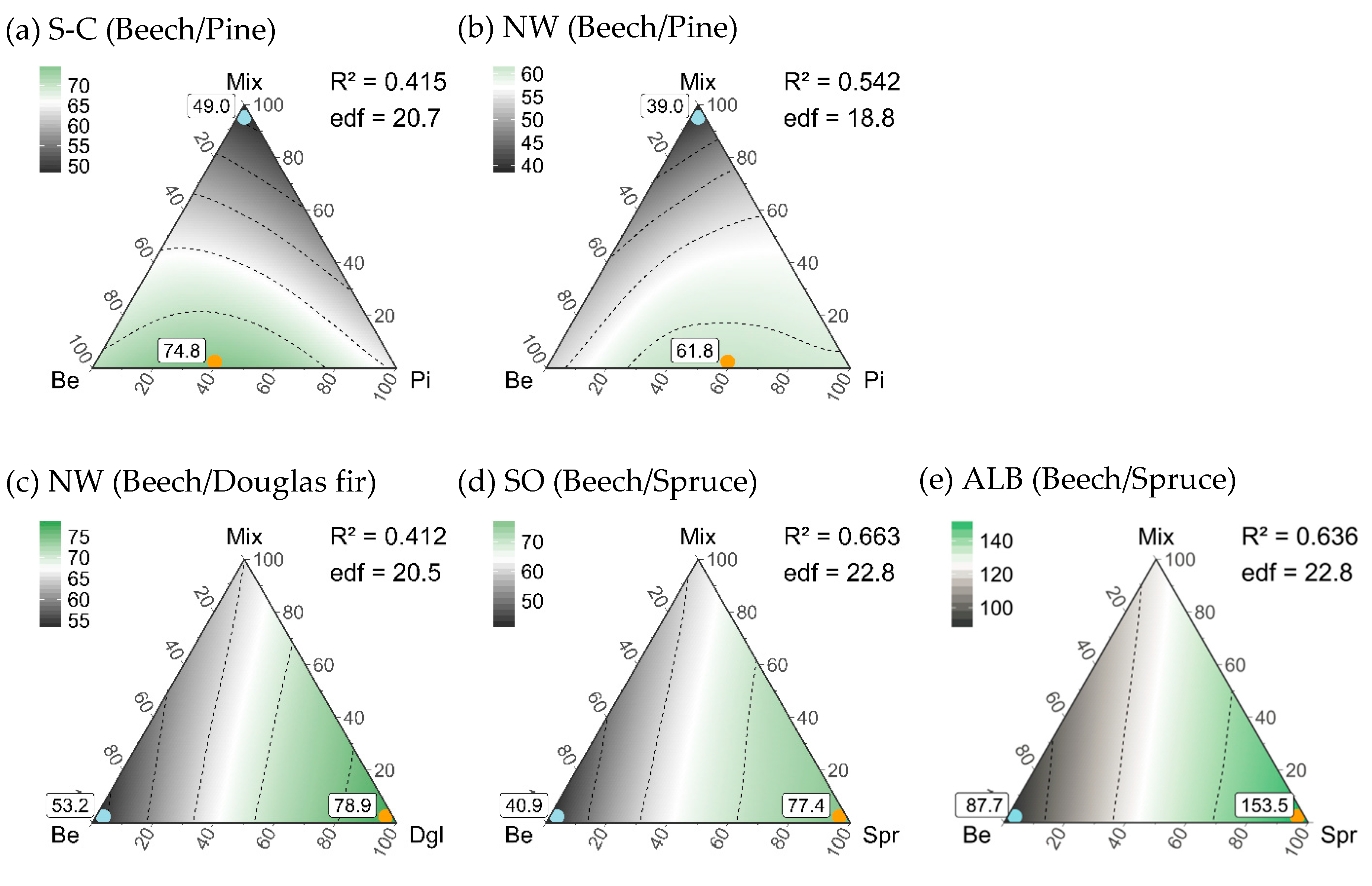
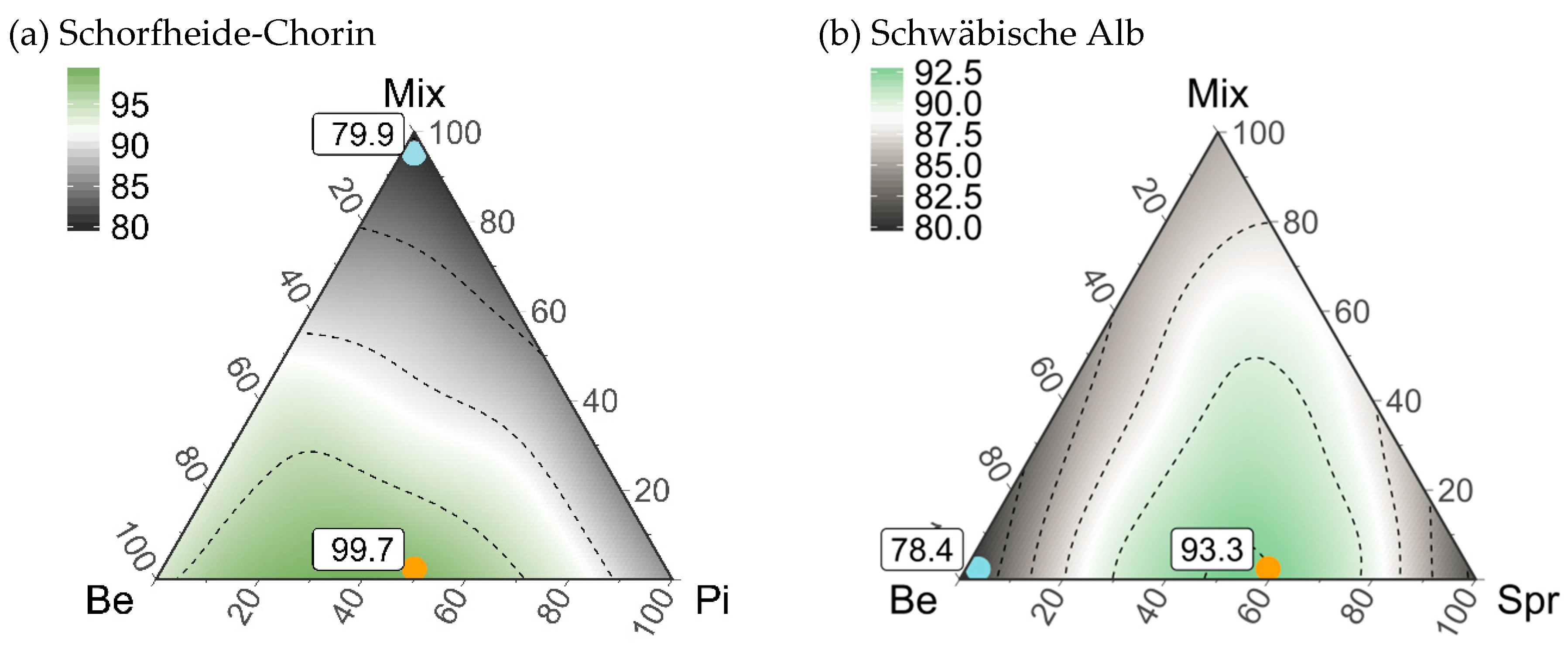
3.5. Composition of Stand Types for Highest Regional Diversity
4. Discussion
4.1. An Optimized Stand Type Composition for Vascular Plant Diversity
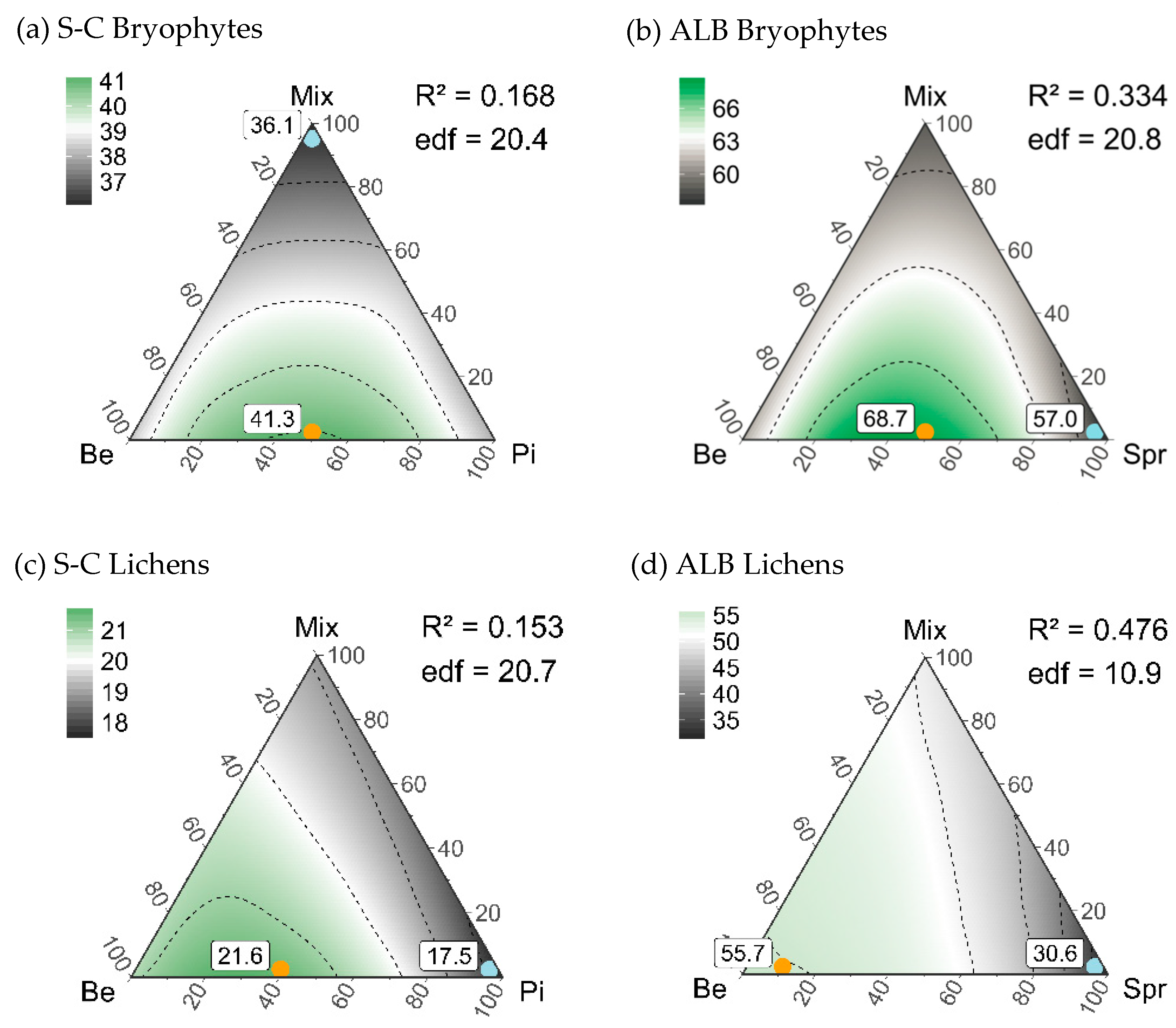
4.2. An Optimized Composition of Forest Stands for Bryophyte and Lichen Diversity
4.3. An Optimized Composition of Pure and Mixed Stands for Regional Diversity
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Northwestern Germany | Schorfheide-Chorin | Solling | Schwäbische Alb | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Stand Age [% Share] | Developmental Phase [% Share] | Stand Age [% Share] | Developmental Phase [% Share] | |||||||||
| n | <90 yrs | >90 yrs | n | Immature | Mature | n | <90 yrs | >90 yrs | n | Immature | Mature | |
| Pure stand types | ||||||||||||
| Beech | 20 | 20.0 | 80.0 | 141 Bry: 108 Li: 76 | 5.1 | 94.9 | 43 | 37.2 | 62.8 | 95 Bry: 64 Li: 34 | 31.8 | 68.2 |
| Pine | 20 | 100.0 | 0.0 | 60 Bry: 39 Li: 25 | 50.0 | 50.0 | - | - | - | - | - | - |
| Spruce | - | - | - | - | - | - | 40 | 62.5 | 37.5 | 61 Bry: 40 Li: 20 | 31.6 | 68.4 |
| Douglas fir | 20 | 100.0 | 0.0 | - | - | - | - | - | - | - | - | - |
| Mixed stand types | ||||||||||||
| Beech/Pine | 20 | 10.0 | 90.0 | 77 Bry: 48 Li: 34 | 2.6 | 97.4 | - | - | - | - | - | - |
| Beech dominance (>60% share) | 20 | 10.0 | 90.0 | 61 Bry: 39 Li: 29 | 1.6 | 98.4 | - | - | - | - | - | - |
| Pine dominance(>60% share) | - | - | - | 6 Bry: 5 Li: 3 | 16.7 | 83.3 | - | - | - | - | - | - |
| Equal share | - | - | - | 10 Bry: 4 Li: 2 | 0.0 | 100.0 | - | - | - | - | - | - |
| Beech/Douglas fir | 20 | 20.0 | 80.0 | - | - | - | - | - | - | - | - | - |
| Beech dominance (>60% share) | 13 | 15.4 | 84.6 | - | - | - | - | - | - | - | - | - |
| Douglas fir dominance (>60% share) | 0 | - | - | - | - | - | - | - | - | - | - | - |
| Equal share | 7 | 28.6 | 71.4 | - | - | - | - | - | - | - | - | - |
| Beech/Spruce | - | - | - | - | - | - | 84 | 54.8 | 45.2 | 40 Bry: 29 Li: 12 | 33.3 | 66.7 |
| Beech dominance (>60% share) | - | - | - | - | - | - | 46 | 52.2 | 47.8 | 17 Bry: 16 Li: 5 | 43.8 | 56.3 |
| Spruce dominance (>60% share) | - | - | - | - | - | - | 8 | 87.5 | 12.5 | 5 Bry: 4 Li: 3 | 20 | 80 |
| Equal share | - | - | - | - | - | - | 30 | 50.0 | 50.0 | 18 Bry: 9 Li: 4 | 26.7 | 73.3 |
| ALB | S-C | SO | NW-Germany | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Be | Be/Spr | Spr | Be | Be/Pi | Pi | Be | Be/Spr | Spr | Be | Be/Dgl | Be/Pi | Dgl | Pi | |
| n | 95 | 40 | 61 | 141 | 77 | 60 | 43 | 84 | 40 | 20 | 20 | 20 | 20 | 20 |
| Abies alba | 1 | 5 | 2 | 8 | ||||||||||
| Carex canescens | 1 | 8 | 8 | |||||||||||
| Cephalanthera damasonium* | 4 | 3 | 6 | |||||||||||
| Daphne mezereum* | 30 | 11 | 10 | |||||||||||
| Epipactis helleborine* | 2 | 3 | 1 | 1 | ||||||||||
| Epipactis purpurata | 2 | |||||||||||||
| Erica tetralix | 2 | |||||||||||||
| Helleborus foetidus* | 12 | 9 | 5 | |||||||||||
| Ilex aquifolium* | 2 | 6 | 4 | 1 | ||||||||||
| Juniperus communis | 1 | 8 | ||||||||||||
| Lilium martagon* | 3 | |||||||||||||
| Listera ovata* | 1 | 2 | ||||||||||||
| Neottia nidus-avis* | 2 | 1 | 1 | |||||||||||
| Platanthera bifolia | 1 | 1 | 2 | |||||||||||
| Primula elatior | 19 | 11 | 12 | |||||||||||
| Ulmus minor | 2 | |||||||||||||
| Taxus baccata | 3 | 1 | ||||||||||||
| Asplenium scolopendrium* | 1 | |||||||||||||
| Cephalanthera rubra | 1 | |||||||||||||
| Corallorhiza trifida | 1 | |||||||||||||
| Hypericum montanum | 1 | |||||||||||||
| Liparis loeselii | 1 | |||||||||||||
| Lunaria rediviva | 1 | |||||||||||||
| Luzula campestris | 1 | |||||||||||||
| Lycopodium annotinum | 1 | |||||||||||||
| Monotropa hypophegea | 1 | |||||||||||||
| Orthilia secunda | 1 | |||||||||||||
| Platanthera chlorantha | 1 | 1 | 1 | |||||||||||
| Total species number | 13 | 11 | 14 | 1 | 0 | 2 | 1 | 3 | 2 | 2 | 2 | 1 | 2 | 2 |
| Schwäbische Alb | Schorfheide-Chorin | |||||
|---|---|---|---|---|---|---|
| Bryophytes | Pure Beech | Mixed | Pure Spruce | Pure Beech | Mixed | Pure Pine |
| n | 64 | 29 | 40 | 108 | 48 | 39 |
| Amblystegium subtile | 20 | 2 | ||||
| Anomodon viticulosus | 2 | |||||
| Brachythecium oedipodium | 5 | 3 | 3 | |||
| Ctenidium molluscum | 2 | 1 | 2 | |||
| Fissidens exilis | 2 | |||||
| Frullania dilatata | 22 | 6 | 1 | 1 | ||
| Homalia trichomanoides | 19 | 3 | ||||
| Hylocomium splendens | 3 | 10 | 33 | 2 | 3 | |
| Isothecium alopecuroides | 55 | 18 | 4 | |||
| Leucobryum glaucum | 2 | 7 | 1 | |||
| Leucodon sciuroides | 5 | |||||
| Metzgeria furcata | 31 | 9 | 2 | |||
| Mnium marginatum | 4 | |||||
| Nowellia curvifolia | 1 | 3 | ||||
| Orthotrichum affine | 38 | 11 | 8 | 14 | 3 | |
| Orthotrichum lyellii | 9 | |||||
| Orthotrichum pallens | 5 | |||||
| Orthotrichum patens | 2 | 1 | ||||
| Orthotrichum pumilum | 18 | 3 | 3 | 3 | ||
| Orthotrichum speciosum | 13 | 3 | 1 | |||
| Orthotrichum stramineum | 8 | 4 | 1 | |||
| Orthotrichum striatum | 9 | 2 | ||||
| Plagiochila asplenioides | 4 | 3 | 20 | |||
| Plagiothecium undulatum | 6 | 1 | ||||
| Platygyrium repens | 8 | 2 | 1 | 13 | 4 | |
| Porella platyphylla | 4 | |||||
| Pseudoleskeella nervosa | 13 | 2 | 1 | |||
| Pterigynandrum filiforme | 28 | 9 | ||||
| Ptilium crista-castrensis | 2 | 1 | ||||
| Pylaisia polyantha | 7 | 5 | ||||
| Radula complanata | 46 | 21 | 4 | 1 | ||
| Rhytidiadelphus loreus | 3 | 8 | 20 | 1 | ||
| Rhytidiadelphus triquetrus | 4 | 10 | 24 | |||
| Tortella tortuosa | 3 | |||||
| Ulota bruchii | 57 | 24 | 8 | 7 | 2 | |
| Ulota crispa | 7 | |||||
| Anomodon attenuatus | 1 | |||||
| Campylium calcareum | 1 | |||||
| Dicranum flagellare | 1 | |||||
| Homomallium incurvatum | 1 | 1 | ||||
| Marchantia polymorpha | 1 | |||||
| Metzgeria temperata | 1 | |||||
| Orthotrichum obtusifolium | 1 | 1 | ||||
| Pleuridium acuminatum | 1 | |||||
| Pleuridium subulatum | 1 | |||||
| Polytrichum commune | 1 | |||||
| Ptilidium ciliare | 1 | |||||
| Rhodobryum roseum | 1 | |||||
| Sanionia uncinata | 1 | 1 | ||||
| Total species number | 36 | 25 | 22 | 10 | 9 | 7 |
| Schwäbische Alb | Schorfheide-Chorin | |||||
|---|---|---|---|---|---|---|
| Lichens | Pure Beech | Mixed | Pure Spruce | Pure Beech | Mixed | Pure Pine |
| n | 34 | 12 | 20 | 76 | 34 | 25 |
| Arthonia punctiformis | 2 | |||||
| Arthonia radiata | 30 | 5 | ||||
| Bacidina arnoldiana* | 12 | 2 | 4 | 15 | 2 | |
| Bacidina delicata* | 3 | 1 | ||||
| Chaenotheca trichialis | 2 | |||||
| Graphis scripta | 28 | 8 | 1 | 6 | 1 | |
| Lecania cyrtella* | 4 | 1 | ||||
| Lecanora intumescens | 9 | 1 | ||||
| Lecanora subcarpinea | 9 | |||||
| Lecanora subrugosa | 5 | |||||
| Mycobilimbia epixanthoides | 2 | |||||
| Opegrapha rufescens | 2 | |||||
| Parmelia saxatilis* | 2 | |||||
| Parmelia sulcata* | 23 | 6 | 9 | |||
| Peltigera praetextata | 4 | |||||
| Pertusaria leioplaca | 29 | 5 | ||||
| Pertusaria pertusa | 2 | |||||
| Physcia aipolia | 3 | |||||
| Pyrenula nitida | 7 | 2 | ||||
| Pyrrhospora quernea | 10 | 1 | ||||
| Ramalina farinacea* | 5 | 1 | 2 | |||
| Strigula stigmatella | 2 | |||||
| Bacidia arceutina | 1 | |||||
| Bacidia rubella | 1 | |||||
| Bacidina chloroticula* | 1 | |||||
| Calicium salicinum | 1 | |||||
| Cetrelia olivetorum | 1 | |||||
| Chaenotheca chrysocephala | 1 | 1 | 1 | |||
| Chaenotheca stemonea | 1 | |||||
| Chrysothrix candelaris | 1 | |||||
| Cladonia ramulosa | 1 | |||||
| Cliostomum griffithii | 1 | |||||
| Fellhanera bouteillei | 1 | |||||
| Lecanora albella | 1 | |||||
| Lecanora argentata | 1 | 1 | ||||
| Leptogium plicatile | 1 | |||||
| Melanelia exasperata | 1 | 1 | ||||
| Parmelia ernstiae* | 1 | |||||
| Pertusaria hymenea | 1 | |||||
| Placynthiella uliginosa | 1 | |||||
| Sarea resinae | 1 | |||||
| Usnea filipendula | 1 | 1 | ||||
| Total species number | 32 | 14 | 9 | 7 | 3 | 1 |
References
- Gamfeldt, L.; Snall, T.; Bagchi, R.; Jonsso, M.; Gustafsson, L.; Kjellander, P.; Ruiz-Jaen, M.C.; Froberg, M.; Stendahl, J.; Philipson, C.D.; et al. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun. 2013, 4, 1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuler, L.J.; Bugmann, H.; Snell, R.S. From monocultures to mixed-species forests: Is tree diversity key for providing ecosystem services at the landscape scale? Landsc. Ecol. 2017, 32, 1499–1516. [Google Scholar] [CrossRef]
- Ratcliffe, S.; Wirth, C.; Jucker, T.; van der Plas, F.; Scherer-Lorenzen, M.; Verheyen, K.; Allan, E.; Benavides, R.; Bruelheide, H.; Ohse, B.; et al. Biodiversity and ecosystem functioning relations in European forests depend on environmental context. Ecol. Lett. 2017, 20, 1414–1426. [Google Scholar] [CrossRef]
- Felipe-Lucia, M.R.; Soliveres, S.; Penone, C.; Manning, P.; van der Plas, F.; Boch, S.; Prati, D.; Ammer, C.; Schall, P.; Gossner, M.M.; et al. Multiple forest attributes underpin the provision of multiple ecosystem services. Nat. Commun. 2018, 9, 4839. [Google Scholar] [CrossRef] [PubMed]
- Chamagne, J.; Tanadini, M.; Frank, D.; Matula, R.; Paine, C.E.T.; Philipson, C.D.; Svátek, M.; Turnbull, L.A.; Volařík, D.; Hector, A. Forest diversity promotes individual tree growth in central European forest stands. J. Appl. Ecol. 2017, 54, 71–79. [Google Scholar] [CrossRef]
- Jucker, T.; Bouriaud, O.; Avacaritei, D.; Coomes, D.A. Stabilizing effects of diversity on aboveground wood production in forest ecosystems. Linking patterns and processes. Ecol. Lett. 2014, 17, 1560–1569. [Google Scholar] [CrossRef] [PubMed]
- Ammer, C. Diversity and forest productivity in a changing climate. New Phytol. 2019, 221, 50–66. [Google Scholar] [CrossRef] [PubMed]
- Mina, M.; Huber, M.O.; Forrester, D.I.; Thürig, E.; Rohner, B. Multiple factors modulate tree growth complementarity in central European mixed forests. J. Ecol. 2018, 106, 1106–1119. [Google Scholar] [CrossRef]
- Pretzsch, H.; Dieler, J.; Seifert, T.; Rötzer, T. Climate effects on productivity and resource-use efficiency of Norway spruce (Picea abies [L.] Karst.) and European beech (Fagus sylvatica [L.]) in stands with different spatial mixing patterns. Trees 2012, 26, 1343–1360. [Google Scholar] [CrossRef]
- Fichtner, A.; Härdtle, W.; Bruelheide, H.; Kunz, M.; Li, Y.; von Oheimb, G. Neighbourhood interactions drive overyielding in mixed-species tree communities. Nat. Commun. 2018, 9, 1144. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G.; Uhl, E. Resistance of European tree species to drought stress in mixed versus pure forests. Evidence of stress release by inter-specific facilitation. Plant Biol. 2013, 15, 483–495. [Google Scholar] [CrossRef]
- Metz, J.; Annighöfer, P.; Schall, P.; Zimmermann, J.; Kahl, T.; Schulze, E.-D.; Ammer, C. Site-adapted admixed tree species reduce drought susceptibility of mature European beech. Glob. Chang. Biol. 2016, 22, 903–920. [Google Scholar] [CrossRef]
- Simmons, E.A.; Buckley, G.P. Ground vegetation under planted mixtures of trees. In The Ecology of Mixed-Species Stands of Trees; Cannell, M.G.R., Malcolm, D.C., Robertson, P.A., Eds.; Blackwell Scientific Publications: Oxford, UK, 1992; pp. 211–231. [Google Scholar]
- Felton, A.; Lindbladh, M.; Brunet, J.; Fritz, Ö. Replacing coniferous monocultures with mixed-species production stands: An assessment of the potential benefits for forest biodiversity in northern Europe. For. Ecol. Manag. 2010, 260, 939–947. [Google Scholar] [CrossRef]
- Scherer-Lorenzen, M. The functional role of biodiversity in forests. In Integrative Approaches as an Opportunity for the Conservation of Forest Biodiversity; Kraus, D., Krumm, F., Eds.; European Forest Institute: Freiburg, Germany, 2013; pp. 216–223. [Google Scholar]
- Gilliam, F.S. The ecological significance of the herbaceous layer in temperate forest ecosystems. BioScience 2007, 57, 845–858. [Google Scholar] [CrossRef]
- Scherber, C.; Eisenhauer, N.; Weisser, W.W.; Schmid, B.; Voigt, W.; Fischer, M.; Schulze, E.-D.; Roscher, C.; Weigelt, A.; Allan, E.; et al. Bottom-up effects of plant diversity on multitrophic interactions in a biodiversity experiment. Nature 2010, 468, 553–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handa, I.T.; Aerts, R.; Berendse, F.; Berg, M.P.; Bruder, A.; Butenschoen, O.; Chauvet, E.; Gessner, M.O.; Jabiol, J.; Makkonen, M.; et al. Consequences of biodiversity loss for litter decomposition across biomes. Nature 2014, 509, 218–221. [Google Scholar] [CrossRef] [Green Version]
- Ódor, P.; Király, I.; Tinya, F.; Bortignon, F.; Nascimbene, J. Patterns and drivers of species composition of epiphytic bryophytes and lichens in managed temperate forests. For. Ecol. Manag. 2013, 306, 256–265. [Google Scholar] [CrossRef] [Green Version]
- Lücke, K.; Schmidt, W. Vegetation und Standortsverhältnisse in Buchen-Fichten Mischbeständen des Sollings. Forstarchiv 1997, 68, 135–143. [Google Scholar]
- Augusto, L.; Dupouey, J.; Ranger, J. Effects of tree species on understory vegetation and environmental conditions in temperate forests. Ann. For. Sci. 2003, 60, 823–831. [Google Scholar] [CrossRef] [Green Version]
- Sydes, C.; Grime, J.P. Effects of tree leaf litter on herbaceous vegetation in deciduous woodland. I. Field investigations. J. Ecol. 1981, 69, 237–248. [Google Scholar] [CrossRef]
- Wilson, S.D. Heterogeneity, diversity and scale in plant communities. In The Ecological Consequences of Environmental Heterogeneity; Hutchings, M.J., John, E.A., Stewart, A.J.A., Eds.; Blackwell Science: Oxford, UK, 2000; pp. 53–69. [Google Scholar]
- Stein, A.; Gerstner, K.; Kreft, H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol. Lett. 2014, 17, 866–880. [Google Scholar] [CrossRef] [Green Version]
- Cavard, X.; Macdonald, S.E.; Bergeron, Y.; Chen, H.Y.H. Importance of mixedwoods for biodiversity conservation: Evidence for understory plants, songbirds, soil fauna, and ectomycorrhizae in northern forests. Environ. Rev. 2011, 19, 142–161. [Google Scholar] [CrossRef]
- Gosselin, M.; Fourcin, D.; Dumas, Y.; Gosselin, F.; Korboulewsky, N.; Toïgo, M.; Vallet, P. Influence of forest tree species composition on bryophytic diversity in mixed and pure pine (Pinus sylvestris L.) and oak (Quercus petraea (Matt.) Liebl.) stands. For. Ecol. Manag. 2017, 406, 318–329. [Google Scholar] [CrossRef]
- Brassard, B.W.; Chen, H.Y.H.; Cavard, X.; Langanière, J.; Reich, P.B.; Bergeron, Y.; Paré, D.; Yuan, Z. Tree species diversity increases fine root productivity through increased soil volume filling. J. Ecol. 2013, 101, 210–219. [Google Scholar] [CrossRef]
- Barbier, S.; Gosselin, F.; Balandier, P. Influence of tree species on understory vegetation diversity and mechanisms involved—A critical review for temperate and boreal forests. For. Ecol. Manag. 2008, 254, 1–15. [Google Scholar] [CrossRef]
- MacDonald, S.E.; Fenniak, T.E. Understory plant communities of boreal mixedwood forests in western Canada: Natural patterns and response to variable-retention harvesting. For. Ecol. Manag. 2007, 242, 34–48. [Google Scholar] [CrossRef]
- Spiecker, H. Silvicultural management in maintaining biodiversity and resistance of forests in Europe—temperate zone. J. Environ. Manag. 2003, 67, 55–65. [Google Scholar] [CrossRef]
- Spathelf, P.; Ammer, C. Forest management of Scots pine (Pinus sylvestris L.) in northern Germany—A brief review of the history and current trends. Forstarchiv 2015, 86, 59–66. [Google Scholar]
- Knoke, T.; Ammer, C.; Stimm, B.; Mosandl, R. Admixing broadleaved to coniferous tree species: A review on yield, ecological stability and economics. Eur. J. For. Res. 2008, 127, 89–101. [Google Scholar] [CrossRef]
- Brang, P.; Spathelf, P.; Larsen, J.B.; Bauhus, J.; Boncčìna, A.; Chauvin, C.; Drössler, L.; García-Güemes, C.; Heiri, C.; Kerr, G.; et al. Suitability of close-to-nature silviculture for adapting temperate European forests to climate change. Forestry 2014, 87, 492–503. [Google Scholar] [CrossRef] [Green Version]
- Von Lüpke, B.; Ammer, C.; Braciamacchie, M.; Brunner, A.; Ceitel, J.; Collet, C.; Deuleuze, C.; Di Placido, J.; Huss, J.; Jankovic, J.; et al. Silvicultural strategies for conversion. In Norway Spruce Conversion—Options and Consequences; European Forest Institute Research Report 18; Spiecker, H., Hansen, J., Klimo, E., Skovsgaard, J.P., Sterba, H., von Teuffel, K., Eds.; Brill: Leiden, The Netherlands, 2004; pp. 121–164. [Google Scholar]
- Ammer, C.; Bickel, E.; Kölling, C. Converting Norway spruce stands with beech—A review on arguments and techniques. Austrian J. For. Sci. 2008, 125, 3–26. [Google Scholar]
- Leuschner, C.; Ellenberg, H. Ecology of Central European Forests; Springer International Publishing: Cham, Switzerland, 2017; ISBN 978-3-319-43040-9. [Google Scholar]
- Emmer, I.M.; Fanta, J.; Kobus, A.T.; Kooijman, A.; Sevink, J. Reversing borealization as a means to restore biodiversity in Central-European mountain forests—An example from the Krkonoše Mountains, Czech Republic. Biodivers. Conserv. 1998, 7, 229–247. [Google Scholar] [CrossRef]
- MLUR-Ministerium für Landwirtschaft, Umweltschutz und Raumordnung des Landes Brandenburg. Waldbaurichtlinie 2004—“Grüner Ordner“ der Landesforstverwaltung Brandenburg; Ministerium für Landwirtschaft, Umweltschutz und Raumordnung: Berlin, Germany, 2004; Available online: http://forst.brandenburg.de/media_fast/4055/wbr2004.pdf (accessed on 18. January 2019).
- Borrass, L.; Kleinschmit, D.; Winkel, G. The “German model” of integrative multifunctional forest management—Analysing the emergence and political evolution of a forest management concept. For. Policy Econ. 2017, 77, 16–23. [Google Scholar] [CrossRef]
- Vitali, V.; Büntgen, U.; Bauhus, J. Silver fir and Douglas fir are more tolerant to extreme droughts than Norway spruce in south-western Germany. Glob. Chang. Biol. 2017, 23, 5108–5119. [Google Scholar] [CrossRef] [PubMed]
- Bartels, S.F.; Chen, H.Y.H. Is understory plant species diversity driven by resource quantity or resource heterogeneity? Ecology 2010, 91, 1931–1938. [Google Scholar] [CrossRef] [PubMed]
- Weckesser, M. Die Bodenvegetation von Buchen-Fichten-Mischbeständen im Solling—Struktur, Diversität und Stoffhaushalt; Cuvillier-Verlag: Göttingen, Germany, 2003; ISBN 3898737373. [Google Scholar]
- Budde, S. Auswirkungen des Douglasienanbaus auf die Bodenvegetation im Nordwestdeutschen Tiefland; Cuvillier-Verlag: Göttingen, Germany, 2006; ISBN 978-3-86727-079-3. [Google Scholar]
- Boch, S.; Prati, D.; Müller, J.; Socher, S.; Baumbach, H.; Buscot, F.; Gockel, S.; Hemp, A.; Hessenmöller, D.; Kalko, E.K.V.; et al. High plant species richness indicates management-related disturbances rather than the conservation status of forests. Basic Appl. Ecol. 2013, 14, 496–505. [Google Scholar] [CrossRef]
- Boch, S.; Prati, D.; Hessenmöller, D.; Schulze, E.-D.; Fischer, M. Richness of lichen species, especially of threatened ones, is promoted by management methods furthering stand continuity. PLoS ONE 2013, 8, e55461. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.; Boch, S.; Prati, D.; Socher, S.A.; Pommer, U.; Hessenmöllder, D.; Schall, P.; Schulze, E.-D.; Fischer, M. Effects of forest management on bryophyte species richness in Central European forests. For. Ecol. Manag. 2019, 432, 850–859. [Google Scholar] [CrossRef]
- Boch, S.; Müller, J.; Prati, D.; Blaser, S.; Fischer, M. Up in the tree—The over-looked richness of bryophytes and lichens in tree crowns. PLoS ONE 2013, 8, e84913. [Google Scholar] [CrossRef]
- Kiebacher, T.; Keller, C.; Scheidegger, C.; Bergamini, A. Hidden crown jewels: The role of tree crowns for bryophyte and lichen species richness in sycamore maple wooded pastures. Biodivers. Conserv. 2016, 25, 1605–1624. [Google Scholar] [CrossRef]
- Gauer, J.; Aldinger, E. Waldökologische Naturräume Deutschlands—Forstliche Wuchsgebiete und Wuchsbezirke. Mitt. Ver. Forstl. Standortskde. Forstpflanzenz. 2005, 43, 1–324. [Google Scholar]
- Hessenmöller, D.; Nieschulze, J.; von Lüpke, N.; Schulze, E.-D. Identification of forest management types from ground-based and remotely sensed variables and the effects of forest management on forest structure and composition. Forstarchiv 2011, 82, 171–183. [Google Scholar]
- Schall, P.; Schulze, E.-D.; Fischer, M.; Ayasse, M.; Ammer, C. Relations between forest management, stand structure and productivity across different types of Central European forests. Basic Appl. Ecol. 2018, 32, 39–52. [Google Scholar] [CrossRef]
- Ganz, M. Entwicklung von Baumartenzusammensetzung und Struktur der Wälder vom Schwarzwald bis auf die Schwäbische Alb—mit besonderer Berücksichtigung der Buche. Dissertation Thesis, University of Freiburg, Freiburg im Breisgau, Germany, 2004. Available online: https://freidok.uni-freiburg.de/data/1616 (accessed on 18. January 2019).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 18. January 2019).
- Wisskirchen, R.; Haeupler, H. Standardliste der Farn-und Blütenpflanzen Deutschlands; Ulmer: Stuttgart, Germany, 1998; ISBN 978-3800133604. [Google Scholar]
- Koperski, M.; Sauer, M.; Braun, W.; Gradstein, S.R. Referenzliste der Moose Deutschlands; Bundesamt für Naturschutz: Bonn, Germany, 2000; ISBN 3-7843-3504-7. [Google Scholar]
- Wirth, V. Flechtenflora, 2nd ed.; Ulmer: Stuttgart, Germany, 1995; ISBN 3-82521062-6. [Google Scholar]
- Crowley, P.H. Resampling methods for computation-intensive data analysis in ecology and evolution. Annu. Rev. Ecol. Syst. 1992, 23, 405–447. [Google Scholar] [CrossRef]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef]
- Wood, S.N. Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric generalized linear models. J. R. Stat. Soc. B 2011, 73, 3–36. [Google Scholar] [CrossRef]
- Hamilton, N. Ggtern: An Extension to ’ggplot2’, for the Creation of Ternary Diagrams. R package version 2.2.1. 2017. Available online: https://CRAN.R-project.org/package=ggtern (accessed on 18. January 2019).
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W. Zeigerwerte von Pflanzen in Mitteleuropa. Scr. Geobot. 2001, 18, 1–262. [Google Scholar]
- Schmidt, M.; Kriebitzsch, W.-U.; Ewald, J. Waldartenlisten der Farn-und Blütenpflanzen, Moose und Flechten Deutschlands; Bundesamt für Naturschutz: Bonn, Germany, 2011; ISBN 978-3-89624-034-7. [Google Scholar]
- Allan, E.; Bossdorf, O.; Dormann, C.F.; Prati, D.; Gossner, M.M.; Tscharntke, T.; Blüthgen, N.; Bellach, M.; Birkhofer, K.; Boch, S.; et al. Interannual variation in land-use intensity enhances grassland multidiversity. Proc. Natl. Acad. Sci. USA 2014, 111, 308–313. [Google Scholar] [CrossRef]
- Colwell, R.K.; Coddington, J.A. Estimating terrestrial biodiversity through extrapolation. Philos. Trans. R. Soc. B 1994, 345, 101–118. [Google Scholar] [Green Version]
- Landolt, E.; Bäumler, B.; Erhardt, A.; Hegg, O.; Klötzli, F.; Lämmler, W.; Nobis, M.; Rudmann-Maurer, K.; Schweingruber, F.H.; Theurillat, J.-P.; et al. Flora Indicative—Ecological Indicator values and Biological Attributes of the Flora of Switzerland and the Alps; Haupt Verlag: Bern, Switzerland, 2010; ISBN 978-3-258-07461-0. [Google Scholar]
- Schmidt, M.; Mölder, A.; Schönfelder, E.; Engel, F.; Schmiedel, I.; Culmsee, H. Determining ancient woodland indicator plants for practical use: A new approach developed in northwest Germany. For. Ecol. Manag. 2014, 330, 228–239. [Google Scholar] [CrossRef]
- Hofmann, G.; Pommer, U. Potentielle Natürliche Vegetation von Brandenburg und Berlin; Eberswalder Forstliche Schriftenreihe 24: Berlin, Germany, 2005; ISBN 3-933352-62-2. [Google Scholar]
- Budde, S.; Schmidt, W.; Weckesser, M. Impact of the admixture of European beech (Fagus sylvatica L.) on plant species diversity and naturalness of conifer stands in Lower Saxony. Wald. Landsch. Nat. 2011, 11, 49–61. [Google Scholar]
- Metz, J.; Seidel, D.; Schall, P.; Scheffer, D.; Schulze, E.-D.; Ammer, C. Crown modeling by terrestrial laser scanning as an approach to assess the effect of aboveground intra- and interspecific competition on tree growth. For. Ecol. Manag. 2013, 310, 275–288. [Google Scholar] [CrossRef]
- Pretzsch, H.; del Río, M.; Schütze, G.; Ammer, C.; Annighöfer, P.; Avdagic, A.; Barbeito, I.; Bielak, K.; Brazaitis, G.; Coll, L.; et al. Mixing of Scots pine (Pinus sylvestris L.) and European beech (Fagus sylvatica L.) enhances structural heterogeneity, and the effect increases with humidity. For. Ecol. Manag. 2016, 373, 149–166. [Google Scholar] [CrossRef]
- Forrester, D.; Ammer, C.; Annighöfer, P.; Barbeito, I.; Bielak, K.; Bravo-Oviedo, A.; Coll, L.; del Río, M.; Drössler, L.; Heym, M.; et al. Effects of crown architecture and stand structure on light absorption in mixed and monospecific Fagus sylvatica and Pinus sylvestris forests along a productivity and climate gradient through Europe. J. Ecol. 2018, 106, 746–760. [Google Scholar] [CrossRef]
- Jucker, T.; Bouriaud, O.; Coomes, D.A. Crown plasticity enables trees to optimize canopy packing in mixed-species forests. Funct. Ecol. 2015, 29, 1078–1086. [Google Scholar] [CrossRef] [Green Version]
- Bolte, A.; Villanueva, I. Interspecific competition impacts on the morphology and distribution of fine roots in European beech (Fagus sylvatica L.) and Norway Spruce (Picea abies (L.) Karst. Eur. J. For. Res. 2006, 125, 15–26. [Google Scholar] [CrossRef]
- Ma, Z.; Chen, H.Y.H. Effects of species diversity on fine root productivity increase with stand development and associated mechanisms in a boreal forest. J. Ecol. 2017, 105, 237–245. [Google Scholar] [CrossRef]
- Máliš, F.; Ujházy, K.; Vodálová, A.; Barka, I.; Čaboun, V.; Sitková, Z. The impact of Norway spruce planting on herb vegetation in the mountain beech forests on two bedrock types. Eur. J. For. Res. 2012, 131, 1551–1569. [Google Scholar] [CrossRef]
- Mölder, A.; Bernhardt-Römermann, M.; Schmidt, W. Herb-layer diversity in deciduous forests: Raised by tree richness or beaten by beech? For. Ecol. Manag. 2008, 256, 272–281. [Google Scholar] [CrossRef]
- Schmid, I.; Leuschner, C. Warum fehlt den Gipsbuchenwäldern des Kyffhäusers (Thüringen) eine Krautschicht? Forstwiss. Centralbl. 1998, 117, 277–288. [Google Scholar] [CrossRef]
- Röhrig, E.; Bartsch, N.; von Lüpke, B. Waldbau auf Ökologischer Grundlage; Ulmer: Stuttgart, Germany, 2006; ISBN 3825283100. [Google Scholar]
- Ulrich, B. Stabilität von Waldökosystemen unter dem Einfluss des Sauren Regens. Allg. Forstz. 1983, 26/27, 670–677. [Google Scholar]
- Höltermann, A.; Klingenstein, F.; Ssymank, A. Naturschutzfachliche Bewertung der Douglasie aus Sicht des Bundesamtes für Naturschutz (BfN). LWF Wissen 2008, 59, 74–81. [Google Scholar]
- Márialigeti, S.; Németh, B.; Tinya, F.; Ódor, P. The effects of stand structure on ground-floor bryophyte assemblages in temperate mixed forests. Biodivers. Conserv. 2009, 18, 2223–2241. [Google Scholar] [CrossRef]
- Tinya, F.; Márialigeti, S.; Király, I.; Németh, B.; Ódor, P. The effect of light conditions on herbs, bryophytes and seedlings of temperate mixed forests in Örség, Western Hungary. Plant Ecol. 2009, 204, 69–81. [Google Scholar] [CrossRef]
- Nebel, M.; Philippi, G. Die Moose Baden-Württembergs, Band 2; Verlag Eugen Ulmer: Stuttgart, Germany, 2001; ISBN 3800135302. [Google Scholar]
- Turetsky, M.R. The role of bryophytes in carbon and nitrogen cycling. Bryologist 2003, 106, 395–409. [Google Scholar] [CrossRef]
- Woziwoda, B.; Parzych, A.; Kopeć, D. Species diversity, biomass accumulation and carbon sequestration in the understorey of post-agricultural Scots pine forests. Silva Fenn. 2014, 48, 1119. [Google Scholar] [CrossRef]
- Boch, S.; Berlinger, M.; Fischer, M.; Knop, E.; Nentwig, W.; Türke, M.; Prati, D. Fern and bryophyte endozoochory by slugs. Oecologia 2013, 172, 817–822. [Google Scholar] [CrossRef] [PubMed]
- Fritz, Ö.; Gustafsson, L.; Larsson, K. Does forest continuity matter in conservation?—A study of epiphytic lichens and bryophytes in beech forests of southern Sweden. Biol. Conserv. 2008, 141, 655–668. [Google Scholar] [CrossRef]
- Brunialti, G.; Frati, L.; Aleffi, M.; Marignani, M.; Rosati, L.; Burrascano, S.; Ravera, S. Lichens and bryophytes as indicators of old-growth features in Mediterranean forests. Plant Biosyst. 2010, 144, 221–233. [Google Scholar] [CrossRef]
- Allouche, O.; Kalyuzhny, M.; Moreno-Rueda, G.; Pizarro, M.; Kadmon, R. Area–heterogeneity tradeoff and the diversity of ecological communities. Prod. Natl. Acad. Sci. USA 2012, 109, 17495–17500. [Google Scholar] [CrossRef] [PubMed]
- Kriebitzsch, W.-U.; Bültmann, H.; von Oheimb, G.; Schmidt, M.; Thiel, H.; Ewald, J. Forest-specific diversity of vascular plants, bryophytes, and lichens. In Integrative Approaches as an Opportunity for the Conservation of Forest Biodiversity; Kraus, D., Krumm, F., Eds.; European Forest Institute: Freiburg, Germany, 2013; pp. 158–169. [Google Scholar]
- Hofmeister, J.; Hošek, J.; Malíček, J.; Palice, Z.; Syrovátková, L.; Steinová, J.; Černajová, I. Large beech (Fagus sylvatica) trees as ‘lifeboats’ for lichen diversity in central European forests. Biodivers. Conserv. 2016, 25, 1073–1090. [Google Scholar] [CrossRef]
- Schall, P.; Gossner, M.M.; Heinrichs, S.; Fischer, M.; Boch, S.; Prati, D.; Jung, K.; Baumgartner, V.; Blaser, S.; Böhm, S.; et al. The impact of even-aged and uneven-aged forest management on regional biodiversity of multiple taxa in European beech forests. J. Appl. Ecol. 2018, 55, 267–278. [Google Scholar] [CrossRef]
- Neuner, S.; Albrecht, A.; Cullmann, D.; Engels, F.; Griess, V.C.; Hahn, W.A.; Hanewinkel, M.; Härtl, F.; Kölling, C.; Staupendahl, K.; Knoke, T. Survival of Norway spruce remains higher in mixed stands under a dryer and warmer climate. Glob. Chang. Biol. 2015, 21, 935–946. [Google Scholar] [CrossRef] [PubMed]
- Ammer, C.; Fichtner, A.; Fischer, A.; Gossner, M.M.; Meyer, P.; Seidl, R.; Thomas, F.M.; Annighöfer, P.; Kreyling, J.; Ohse, B.; Berger, U.; et al. Key ecological research questions for Central European forests. Basic Appl. Ecol. 2018, 32, 3–25. [Google Scholar] [CrossRef]

| Northwestern Germany | Schorfheide-Chorin | Solling Hills | Schwäbische Alb | |
|---|---|---|---|---|
| Area | ~25,000 km2 | ~1300 km2 | ~350 km2 | ~420 km2 |
| Coordinates | 53°18′ N–53°39′ N; 8°30′ E–10°40′ E | 52°52′ N–53°12′ N; 13°37′ E–14°1′ E | 51°40′ N–51°50′ N; 9°26′ E–9°44′ E | 48°21′ N–48°31′ N; 9°13′ E–9°31′ E |
| Elevation | 0–150 m a.s.l. | 3–140 m a.s.l. | 300–450 m a.s.l. | 460–860 m a.s.l. |
| Bedrock | Glacial sedimentary deposits (partly with loess cover); old Pleistocene | Glacial series; young Pleistocene | Red sandstone with loess cover | Jurassic limestone |
| Predominant soil type | Sandy-podsol-cambisol | Cambisol | Acid silty loam cambisol | Cambisol and leptosol |
| Mean annual temperature | 8.1–9.3 °C | 8.2–8.6 °C | 7.3–7.8 °C | 5.7–7.6 °C |
| Annual Precipitation | 560–840 mm | 555–590 mm | 915–1030 mm | 843–1096 mm |
| Potential natural vegetation | Acidic beech forest | Acidic to mesic beech forests (partly with sessile oak) | Acidic beech forest | Beech forests on limestone |
| Tree species composition | Forest area ~ 27% Share pine ~ 37% Spruce ~ 25% Beech ~ 5% Other conifers ~ 7% | Forest area ~ 48% Share pine ~ 69% Beech ~ 7% Oak ~ 6% | Forest area ~ 94% Share spruce ~ 62% Beech ~ 30% Oak ~ 8% | Forest area ~ 47% Share beech ~ 39% Spruce ~ 38% Other broadleaves ~ 12% |
| Investigated forest types (stand age, establish-ment) | Pure beech (76–120 yrs, natural regeneration) Pure pine (51–82 yrs, planted) Pure Douglas fir (53–64 yrs, planted) Beech/pine mixture (79–161 yrs, beech planted under pine) Beech/Douglas fir mixture (61–98 yrs, Douglas fir planted in beech regeneration) | Pure beech (~70–160 yrs, natural regeneration) Pure pine (~30–70 yrs, planted) Beech/pine mixture (46–130 yrs, beech planted under pine) | Pure beech (52–150 yrs, natural regeneration) Pure spruce (54–132 yrs, planted) Beech/spruce mixture (48–149 yrs, spruce planted in beech regeneration gaps) | Pure beech (~60–100 yrs, natural regeneration) Pure spruce (~40–60 yrs, planted) Beech/spruce mixture (~60–100 yrs, spruce planted in beech regeneration gaps) |
| No. of plots | 100 | 278 | 167 | 196 |
| Beech/Pine (Be/Pi) | Beech/Douglas fir (Be/Dgl) | Beech/Spruce (Be/Spr) | |||
|---|---|---|---|---|---|
| Region | Schorfheide-Chorin | Northwestern Germany | Solling | Schwäbische Alb | |
| n | 17 | 17 | 17 | 17 | 17 |
| Alpha Diversity | |||||
| Beech | 14.0a (7.2–20.8) | 9.0b (6.7–10.2) | 8.9c (6.7–10.2) | 10.4c (6.1–14.8) | 22.5b (16.8–28.8) |
| Conifer | 15.9a (13.1–18.7) | 18.8a (17.8–19.8) | 21.4a (18.6–23.4) | 26.1a (20.7–31.8) | 36.9a (25.8–48.7) |
| Mixed | 8.6b (4.9–13.1) | 7.1c (6.1–8.0) | 14.7b (11.9–16.7) | 17.7b (11.6–23.6) | 33.1a (25.7–40.6) |
| Gamma Diversity | |||||
| Beech | 71.2a (49–94) | 53.3b * (41–56) | 53.2c * (41–56) | 40.9c * (22–56) | 87.7c * (60–114) |
| Conifer | 64.5ab (48–83) | 60.9a (54–64) | 78.9a (67–83) | 77.4a (62–88) | 153.5a (117–187) |
| Mixed | 49.0b * (29–64) | 39.0c * (30–42) | 66.2b * (44–70) | 61.6b* (43–78) | 123.7b * (101–148) |
| Stand Type Composition | |||||
| Maximum gamma diversity | 60%Be-40%Pi-0%Mix | 40%Be-60%Pi-0%Mix | 0%Be-100%Dgl-0%Mix | 0%Be-100%Spr-0%Mix | 0%Be-100%Spr-0%Mix |
| 74.8 (49–97) | 61.8 (47–76) | 78.9 (67–83) | 77.4 (62–88) | 153.5 (117–187) | |
| Minimum gamma diversity | 0%Be-0%Pi-100%Mix | 0%Be-0%Pi-100%Mix | 100%Be-0%Dgl-0%Mix | 100%Be-0%Spr-0%Mix | 100%Be-0%Spr-0%Mix |
| 49.0 * (29–64) | 39.0 * (30–42) | 53.2 * (41–56) | 40.9 * (22–56) | 87.7 * (60–114) | |
| Schorfheide-Chorin (Beech(Be)/Pine(Pi)) | Schwäbische Alb (Beech(Be)/Spruce(Spr)) | |||
|---|---|---|---|---|
| Bryophytes | Lichens | Bryophytes | Lichens | |
| n | 36 | 22 | 26 | 10 |
| Alpha Diversity | ||||
| Beech | 9.8b (8.8–10.9) | 5.0a (3.8–6.3) | 13.4b (11.3–15.7) | 19.5a (15.4–24.0) |
| Conifer | 11.2a (10.8–11.7) | 5.4a (5.0–5.8) | 16.1a (14.1–17.8) | 9.0c (6.8–11.4) |
| Mixed | 10.7ab (9.9–11.7) | 5.5a (4.6–6.4) | 14.1b (13.0–14.9) | 15.1b (12.7–17.4) |
| Gamma Diversity | ||||
| Beech | 38.0a (29–48) | 20.9a (12–32) | 60.7a (49–69) | 55.7a (44–64) |
| Conifer | 38.1a (35–39) | 17.5a (15–18) | 57.0a * (50–63) | 30.6b * (23–37) |
| Mixed | 36.1a (31–38) | 19.0a (13–22) | 58.8a * (53–60) | 49.7a (41–54) |
| Stand Type Composition | ||||
| Maximum gamma diversity | 50%Be-50%Pi-0%Mix | 60%Be-40%Pi-0%Mix | 50%Be-50%Spr-0%Mix | 90%Be-10%Spr-0%Mix |
| 41.3 (34–50) | 21.6 (15–31) | 68.7 (57–79) | 55.7 (44–67) | |
| Minimum gamma diversity | 0%Be-0%Pi-100%Mix | 0%Be-100%Pi-0%Mix | 0%Be-100%Spr-0%Mix | 0%Be-100%Spr-0%Mix |
| 36.1 (31–38) | 17.5 (15–18) | 57.0 (50–63) | 30.6 (23–37) | |
| Characteristic Species of | Beech/Pine | Beech/Douglas Fir | Beech/Spruce | EIV | FA | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| S-C | NW | NW | SO | ALB | L | M | R | N | |||
| pure conifer stands | |||||||||||
| Calluna vulgaris | C | 8 | x | 1 | 1 | 2.1 | |||||
| Epilobium angustifolium | C | C | 8 | 5 | 5 | 8 | 1.2 | ||||
| Rumex acetosella | C | 8 | 3 | 2 | 2 | 2.2 | |||||
| Rubus fruticosus agg. b | C | C | 7.7 | 5.0 | 4.8 | 4.6 | 2.1 | ||||
| Agrostis capillaris | C | 7 | x | 4 | 4 | 2.1 | |||||
| Betula pendula | C | C | 7 | x | x | x | 2.1 | ||||
| Cirsium palustre | C | 7 | 8 | 4 | 3 | 2.1 | |||||
| Digitalis purpurea | C | 7 | 5 | 3 | 6 | 1.2 | |||||
| Galeopsis tetrahit | C | 7 | 5 | x | 6 | 2.1 | |||||
| Galium mollugo agg. c | C | 7 | 5 | 7 | 5 | 2.2 | |||||
| Galium saxatile | C | C | 7 | 5 | 2 | 3 | 2.1 | ||||
| Molinia caerulea | C | 7 | 7 | x | 2 | 2.1 | |||||
| Pinus sylvestris | C | 7 | x | x | x | 2.1 | |||||
| Quercus robur | C | 7 | x | x | x | 2.1 | |||||
| Taraxacum sect. Ruderalia | C | 7 | 5 | x | 8 | 2.1 | |||||
| Cerastium holosteoides | C | 6 | 5 | x | 5 | 2.2 | |||||
| Frangula alnus | C | 6 | 8 | 4 | x | 2.1 | |||||
| Myosotis sylvatica | C | 6 | 5 | x | 7 | 1.2 | |||||
| Stellaria media | C | 6 | x | 7 | 8 | 2.2 | |||||
| Veronica officinalis | C | 6 | 4 | 3 | 4 | 2.1 | |||||
| Ceratocapnos claviculata | C | 5 | 5 | 3 | 6 | 1.2 | |||||
| Dryopteris carthusiana | C | 5 | x | 4 | 3 | 2.1 | |||||
| Epilobium montanum | C | 4 | 5 | 6 | 6 | 2.1 | |||||
| Athyrium filix-femina | C | 3 | 7 | x | 6 | 1.1 | |||||
| Galium rotundifolium | C | 2 | 5 | 5 | 4 | 1.1 | |||||
| Ø | 6.3 | 5.4 | 4.1 | 4.8 | |||||||
| pure stands | |||||||||||
| Moehringia trinervia | C/B | 4 | 5 | 6 | 7 | 1.1 | |||||
| pure beech stands | |||||||||||
| Carex pilulifera | B | 5 | 5 | 3 | 3 | 2.1 | |||||
| Viola riviniana | B | 5 | 4 | 4 | x | 1.1 | |||||
| Impatiens parviflora | B | 4 | 5 | x | 6 | 1.1 | |||||
| Milium effusum | B | 4 | 5 | 5 | 5 | 1.1 | |||||
| Carex remota | B | 3 | 8 | x | x | 1.1 | |||||
| Ø | 4.2 | 5.4 | 4.0 | 4.7 | |||||||
| pure and mixed stands in at least one region | |||||||||||
| Urtica dioica | C/M | x | 6 | 7 | 9 | 2.1 | |||||
| Euphorbia cyparissias | C/M | 8 | 3 | x | 3 | 2.1 | |||||
| Hypericum perforatum | C/M | 7 | 4 | 6 | 4 | 2.1 | |||||
| Rubus idaeus | C | C/M | C/M | 7 | x | x | 6 | 1.2 | |||
| Sambucus nigra | C/M | 7 | 5 | x | 9 | 2.1 | |||||
| Sambucus racemosa | C/M | 6 | 5 | 5 | 8 | 2.1 | |||||
| Deschampsia flexuosa | C | C | C/M | C | 6 | x | 2 | 3 | 2.1 | ||
| Pteridium aquilinum | C | M | 6 | 5 | 3 | 3 | 1.1 | ||||
| Sorbus aucuparia | C | C | C/M | C | C | 6 | x | 4 | x | 2.1 | |
| Geranium robertianum | C/M | 5 | x | x | 7 | 2.1 | |||||
| Picea abies | C/M | C/M | 5 | x | x | x | 2.1 | ||||
| Trientalis europaea | C | C/M | C | 5 | x | 3 | 2 | 1.1 | |||
| Vaccinium myrtillus | C/M | C | C | C | 5 | x | 2 | 3 | 2.1 | ||
| Dryopteris dilatata | C | C/M | C/M | 4 | 6 | x | 7 | 1.1 | |||
| Impatiens noli-tangere | C/M | 4 | 7 | 7 | 6 | 1.1 | |||||
| Mycelis muralis | C/M | C | C/M | 4 | 5 | x | 6 | 2.1 | |||
| Abies alba | C/M | 3 | x | x | x | 1.1 | |||||
| Pseudotsuga menziesiid | C/M | 3 | 6 | 3 | 3 | 1.1 | |||||
| Oxalis acetosella | C/M | 1 | 5 | 4 | 6 | 1.1 | |||||
| Anemone nemorosa | B | B/M | x | 5 | x | x | 1.1 | ||||
| Anemone ranunculoides | B/M | 3 | 6 | 8 | 8 | 1.1 | |||||
| Fagus sylvatica | B/M | B/M | 3 | 5 | x | x | 1.1 | ||||
| Ø | 5.4 | 5.2 | 4.7 | 5.6 | |||||||
| Characteristic Species of | Beech/Pine | Beech/Spruce | EIV | FA | SUB | |||
|---|---|---|---|---|---|---|---|---|
| S-C | ALB | L | M | R | ||||
| pure conifer stands | ||||||||
| Hypnum jutlandicum | C | 7 | 2 | 2 | 2.1 | S,B,D | ||
| Lophocolea bidentata | C | 7 | 6 | 5 | 2.1 | S,B,D | ||
| Rhytidiadelphus squarrosus | C | 7 | 6 | 5 | 2.2 | S,D | ||
| Hylocomium splendens * | C | 6 | 4 | 5 | 2.1 | S,D | ||
| Pleurozium schreberi | C | C | 6 | 4 | 2 | 2.1 | S,D | |
| Scleropodium purum | C | C | 6 | 4 | 5 | 2.1 | S,D | |
| Dicranella heteromalla | C | 5 | 4 | 2 | 2.1 | S,B,D | ||
| Eurhynchium angustirete | C | 5 | 4 | 7 | 1.1 | S,B,D | ||
| Orthodontium lineare | C | 4 | 5 | 2 | 2.1 | S,B,D | ||
| Plagiochila asplenioides * | C | 4 | 6 | 6 | 1.1 | S,D | ||
| Plagiomnium undulatum | C | 4 | 6 | 6 | 2.1 | S,B,D | ||
| Thuidium tamariscinum | C | 4 | 6 | 4 | 2.1 | S,B,D | ||
| Ø | 5.4 | 4.8 | 4.3 | |||||
| pure beech stands | ||||||||
| Amblystegium subtile * | B | 7 | 5 | 6 | 1.1 | B | ||
| Atrichum undulatum | B | 6 | 6 | 4 | 2.1 | S,D | ||
| Dicranum montanum | B | 6 | 5 | 2 | 2.1 | S,B,D | ||
| Isothecium alopecuroides * | B | 5 | 5 | 6 | 1.1 | S,B,D | ||
| Ø | 6.0 | 5.3 | 4.5 | |||||
| pure and mixed stands in at least one region | ||||||||
| Rhytidiadelphus triquetrus * | C/M | 7 | 4 | 5 | 2.1 | S,D | ||
| Dicranum polysetum | C | C/M | 6 | 4 | 5 | 2.1 | S | |
| Dicranum scoparium | C/M | 5 | 4 | 4 | 2.1 | S,B,D | ||
| Herzogiella seligeri | C/M | 5 | 5 | 4 | 1.1 | S,B,D | ||
| Rhytidiadelphus loreus | C/M | 4 | 6 | 3 | 1.1 | S,D | ||
| Frullania dilatata * | B/M | 8 | 4 | 5 | 2.1 | B | ||
| Radula complanata * | B/M | 7 | 5 | 7 | 2.1 | S,B,D | ||
| Pterigynandrum filiforme | B/M | 6 | 5 | 4 | 1.1 | B | ||
| Metzgeria furcata * | B/M | 5 | 4 | 6 | 2.1 | S,B,D | ||
| Ulota bruchii * | B/M | 4 | 5 | 4 | 2.1 | B,D | ||
| Ø | 5.7 | 4.6 | 4.7 | |||||
| Characteristic Species of | Beech/Pine | Beech/Spruce | EIV | FA | SUB | ||||
|---|---|---|---|---|---|---|---|---|---|
| S-C | ALB | L | M | R | N | ||||
| pure conifer stands | |||||||||
| Platismatia glauca | C | 7 | 5 | 2 | 2 | 1.1 | B,D | ||
| Scoliciosporum chlorococcum | C | 6 | 3 | 3 | 6 | 2.1 | B | ||
| Micarea prasina | C | 3 | 4 | 4 | 4 | 1.1 | B,D | ||
| Ø | 5.3 | 4.0 | 3.0 | 4.0 | |||||
| pure beech stands | |||||||||
| Xanthoria polycarpa | B | 7 | 3 | 7 | 8 | 2.1 | B,D | ||
| Arthonia radiata * | B | 3 | 4 | 5 | 4 | 1.1 | B | ||
| Lecanora subcarpinea * | B | nd | nd | nd | nd | 2.2 | B | ||
| Lecanora subrugosa * | B | nd | nd | nd | nd | 2.1 | B | ||
| Ø | 5.0 | 3.5 | 6.0 | 6.0 | |||||
| pure and mixed stands in at least one region | |||||||||
| Lecanora conizaeoides | C/M | 7 | 3 | 2 | 5 | 2.2 | B | ||
| Hypocenomyce scalaris | C/M | 6 | 3 | 2 | 2 | 2.1 | B,D | ||
| Lecanora chlarotera | B/M | 6 | 3 | 6 | 5 | 2.2 | B | ||
| Phlyctis argena | B/M | 5 | 3 | 4 | 5 | 2.1 | B | ||
| Pertusaria leioplaca * | B/M | 4 | 4 | 5 | 2 | 1.1 | B | ||
| Graphis scripta * | B/M | 3 | 4 | 5 | 3 | 1.1 | B | ||
| Porina aenea | B/M | 3 | 4 | 5 | 4 | 1.1 | B | ||
| Arthonia spadicea | B/M | 2 | 4 | 4 | 3 | 1.1 | B,D | ||
| Ø | 4.5 | 3.5 | 4.1 | 3.6 | |||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heinrichs, S.; Ammer, C.; Mund, M.; Boch, S.; Budde, S.; Fischer, M.; Müller, J.; Schöning, I.; Schulze, E.-D.; Schmidt, W.; et al. Landscape-Scale Mixtures of Tree Species are More Effective than Stand-Scale Mixtures for Biodiversity of Vascular Plants, Bryophytes and Lichens. Forests 2019, 10, 73. https://doi.org/10.3390/f10010073
Heinrichs S, Ammer C, Mund M, Boch S, Budde S, Fischer M, Müller J, Schöning I, Schulze E-D, Schmidt W, et al. Landscape-Scale Mixtures of Tree Species are More Effective than Stand-Scale Mixtures for Biodiversity of Vascular Plants, Bryophytes and Lichens. Forests. 2019; 10(1):73. https://doi.org/10.3390/f10010073
Chicago/Turabian StyleHeinrichs, Steffi, Christian Ammer, Martina Mund, Steffen Boch, Sabine Budde, Markus Fischer, Jörg Müller, Ingo Schöning, Ernst-Detlef Schulze, Wolfgang Schmidt, and et al. 2019. "Landscape-Scale Mixtures of Tree Species are More Effective than Stand-Scale Mixtures for Biodiversity of Vascular Plants, Bryophytes and Lichens" Forests 10, no. 1: 73. https://doi.org/10.3390/f10010073
APA StyleHeinrichs, S., Ammer, C., Mund, M., Boch, S., Budde, S., Fischer, M., Müller, J., Schöning, I., Schulze, E.-D., Schmidt, W., Weckesser, M., & Schall, P. (2019). Landscape-Scale Mixtures of Tree Species are More Effective than Stand-Scale Mixtures for Biodiversity of Vascular Plants, Bryophytes and Lichens. Forests, 10(1), 73. https://doi.org/10.3390/f10010073