1. Introduction
Atomic level simulation of macromolecules is currently performed almost exclusively by molecular dynamics. Its success, due to it following the actual time course of the system, is also the source of its ultimate limitation—that of the time scale reachable by it. However, it was argued earlier that Monte Carlo, the preferred technique at the early times of molecular simulations, has the potential of overcome this limitation as it is not bound to time [
1]. In spite of this potential, there are few groups working along this line [
2,
3,
4]. A further advantage of the Monte Carlo approach over molecular dynamics is the ease of limiting the sampling to certain parts of the system and/or to a limited set of degrees of freedom.
Representing the field of rigid macromolecules on grids specific to the type of the interacting atoms is a standard technique employed in many docking softwares [
5,
6] where the target is held rigid and a flexible ligand is moved around the target. Assuming that there are N
R atoms in the rigid part, in the energy calculation for each moving atom, the grid representation of the rigid part of the macromolecule replaces the calculation of N
R distances with the calculation of the distances from the eight nearest gridpoints followed by interpolating the corresponding eight energy terms adding an effort equivalent to about two more distance calculations. Since the most likely macromolecule type to be thus modeled is a protein and protein domains have well over a thousand atoms (i.e., N
R ≃ 1000 is a low estimate in most cases), using the grid representation saves a factor of N
R/10, that is, likely a factor of ~100 or more. This idea has also been combined with the Monte Carlo approach for the simulation of protein loops [
7] where only a small part of the molecule is moved during the simulation, achieving similar speedup. The fact that the rigid part of the macromolecule is also kept in place was an important aspect for the success of these applications since it allowed keeping the grid(s) unchanged during the entire simulation. The algorithm described here is designed to maintain the gains achievable by the use of the grids for the case when a macromolecule is moved.
When modeling the interaction of two partially rigid macromolecules, the straightforward extension of these algorithms would run into the difficulty of having to move at least one of the sets of grids. This introduces additional computation expenses: (a) finding the nearest eight gridpoints would be more involved since the rotated grids would not be aligned with the Cartesian axes and (b) the additional computational burden of transforming Nt * Ng3 grid values (with Nt being the number of grid types and Ng the number of gridpoints along one Cartesian direction) whenever a whole macromolecule is moved. The proposed algorithm would eliminate these problems at negligible added computational cost.
2. The Algorithm Proposed
This note describes an algorithm that allows the movement of the rigid part while maintaining the grids in their original positions, giving rise to the same savings as obtained for the single molecule system.
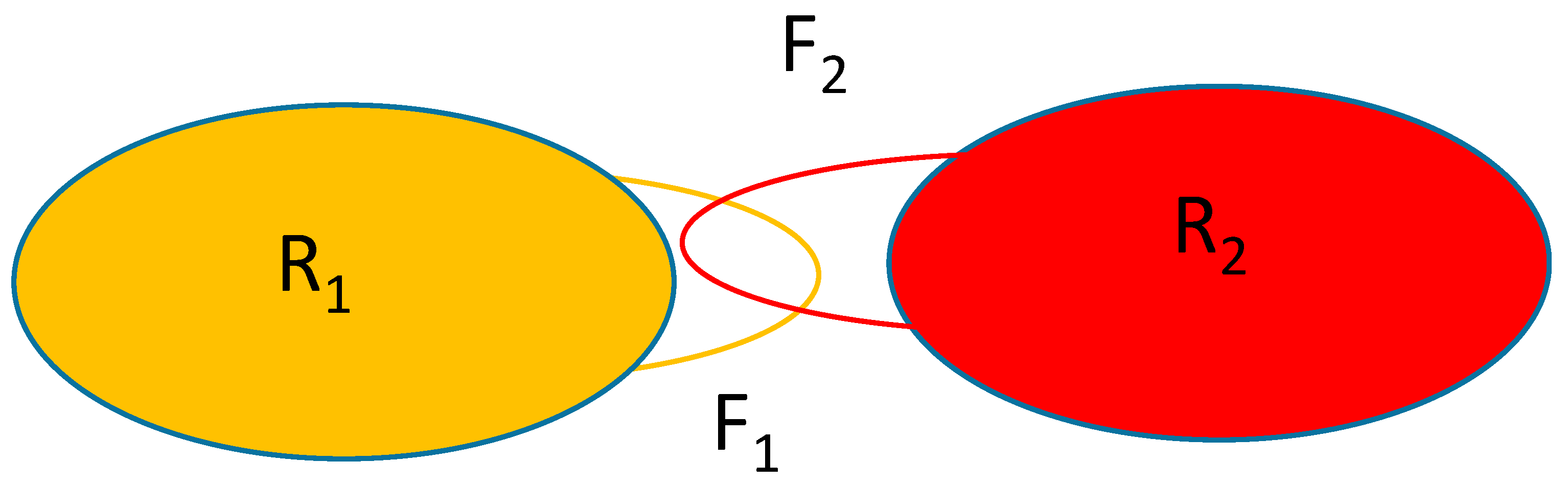
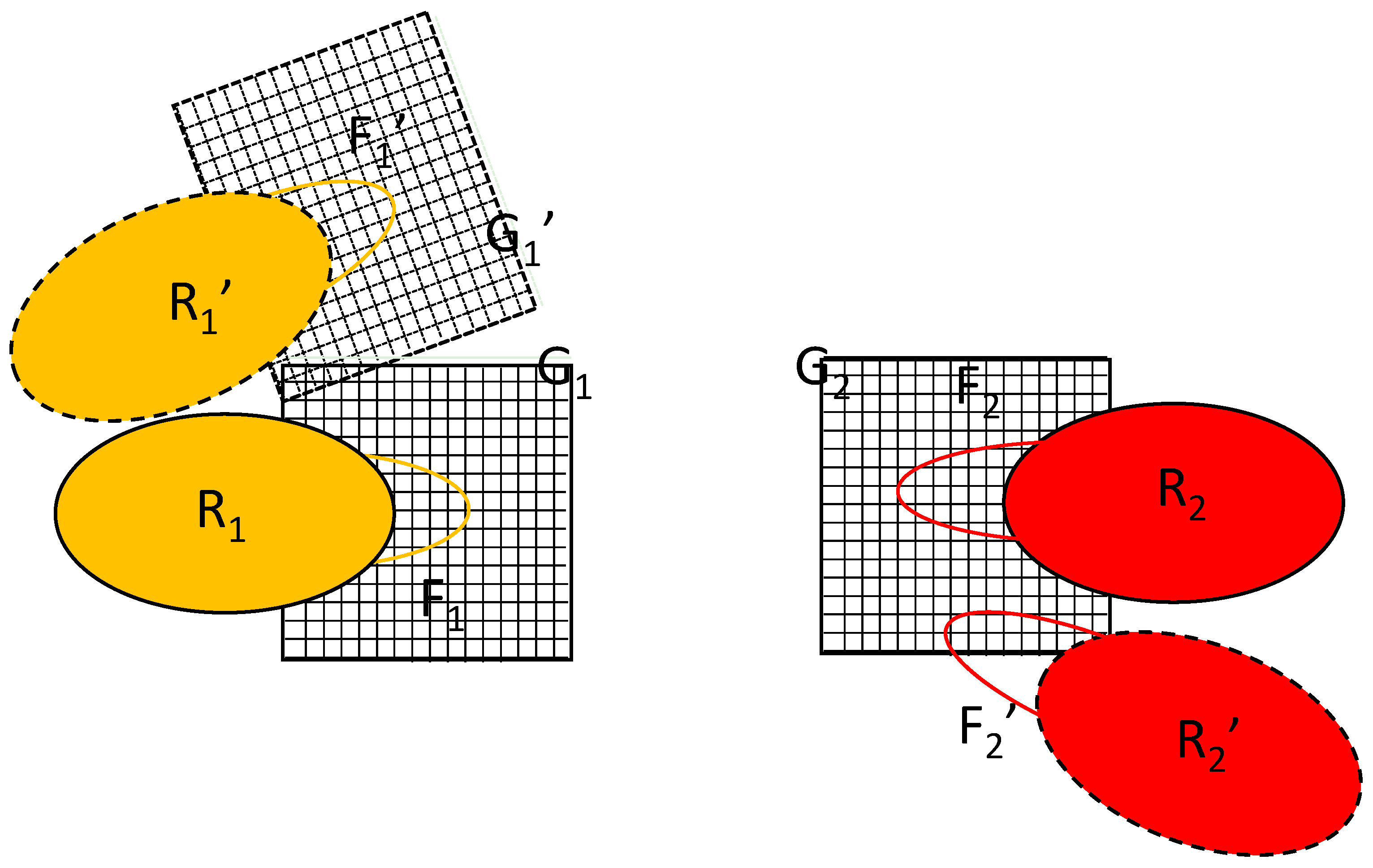
Figure 1 shows the schematics of the system considered. The rigid parts of the macromolecules are represented by the oblongs labeled with R
1 and R
2 and the flexible parts are represented by the loops F
1 and F
2. Macromolecule 2 will be held in place with only the F
2 part moving while macromolecule 1 is allowed to translate/rotate whilst also changing the conformation of the part represented by the atoms forming F
1.
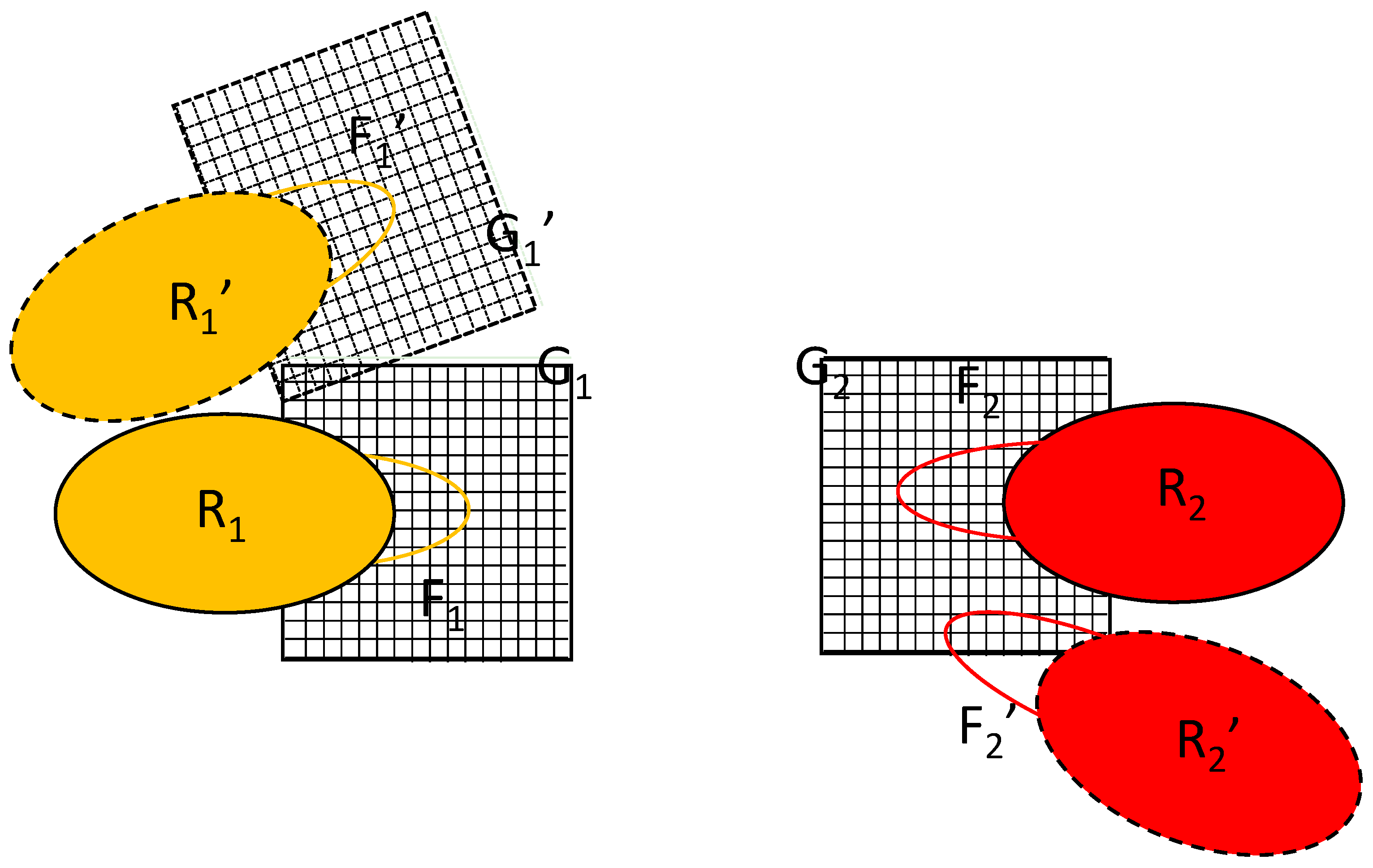
The key to the algorithm proposed is the simultaneous updating of two equivalent copies of the flexible parts, labeled F
1′ and F
2′ in
Figure 2. The difference between the primed and unprimed versions is just a translation/rotation of the rigid part; their conformations are identical. The initial position of macromolecule 1 is R
1 and its position after rigid translation/rotation is R
1′. Such a transformation would also require the transformation of grid G
1 into G
1′, as shown in
Figure 2, but the proposed algorithm avoids this. Instead, the algorithm introduces the alternative position of macromolecule 2, R
2′. It is constructed by translating/rotating the whole complex in such a way that the rigid part of macromolecule 1 is overlaid on its initial position/orientation. Note that, in reality, only the coordinates of the flexible part F
2 are involved in the calculation.
Since
E(F
1′,G
1′) =
E(F
1,G
1) and
E(F
2,G
1′) =
E(F
2′,G
1) where
E(F,G) represents the energy of flexible part F interacting with grid G, this arrangement allows the calculation of the energy of a new complex as follows:
where
E(F
1′,F
2) is the energy of the interaction between flexible parts F
1′ and F
2.
Two kinds of changes are considered: translate/rotate macromolecule 1 in the current conformation of its flexible part, F1, or change the conformation of the flexible part of one of the macromolecules, F1 or F2. When macromolecule 1 is translated/rotated, the coordinates of F1′ and F2′ have to be updated. When the flexible part of macromolecule 1 is changed, the coordinates of F1 and F1′ have to be updated. When the flexible part of macromolecule 2 is changed, the coordinates of F2 and F2′ have to be updated.
Note that in Equation (1), the energy of the interaction between the two rigid parts, being separated by the layers of the flexible part, is assumed to be negligible. For the Van der Waals interactions, this assumption holds to a good approximation. As for the electrostatic interactions, it is possible to include them, at negligible computational expense, using a multipole representation of the charge distribution since the two rigid parts can be enclosed into nonoverlapping spheres, ensuring the validity of the multipole representation.
3. Discussion
This note presented an algorithm that extends the use of grid-based force fields to the modeling of two partially flexible macromolecules by eliminating the added computation involved in determining the nearest gridpoints, and the computation to transform the grids. As for the determination of the nearest gridpoints, the proposed algorithm’s use of a transformed copy is adding a similar computational expense, so there is no net effect. The main saving is achieved by eliminating the transformation of the grids.
Assuming that there are 10 different grids corresponding to different atom types, the proposed algorithm would save the transformation of ~10 × Ng3 points at the negligible expense of transforming the additional NF atoms of the flexible part. Assuming that Ng ≃ 200 (a reasonable value in our experience) and that the flexible part is about 10% of the whole molecule, about ~8 × 105 transformations can be eliminated at the negligible expense of ~2 × 102 additional transformations of the flexible part. The estimate of the overall effect is less straightforward. It is reasonable to assume that the volume of the grid is proportional to the volume of the flexible atoms or, equivalently, that the number of grids along an axis is proportional to the cube root of the number of flexible atoms, i.e., Ng = c1 × NF1/3, and that the overall energy calculation will be dominated by the direct interactions of the flexible parts. Assuming further, as is generally true for Markov-chain Monte Carlo, that a move involves a small limited fraction of the atoms available for moving, the work in calculating the energy change is c2 × NF. This means that the ratio of the work involved in the energy calculation per step and the corresponding savings by using the algorithm proposed here is [c2 × (NF)]/[W × 10 × (c1 × NF1/3)3] = c2/(10 × W × c13) where W is the fraction of the time during which the whole molecule is moved. Thus, in the limit of a large number of flexible atoms, the expense of transforming the grid is commensurable to the rest of the energy calculation, thus the upper limit of the savings is a constant, depending, among others, on the frequency of whole molecule moves. Likely applications, however, are far from this limit: assuming (a relatively large) NF ≃ 100 and Ng ≃ 200 and the number of changed flexible atoms ~50 (an overestimate), the ratio of the energy calculation to the grid transformation calculation would be (50 × 100)/(W × 10 × 2003) = 10 −4(5/(W × 8) × F where F is the ratio of the computation expenses of calculating a distance and transforming a coordinate. Clearly, it shows a significant gain for systems of systems of likely size.
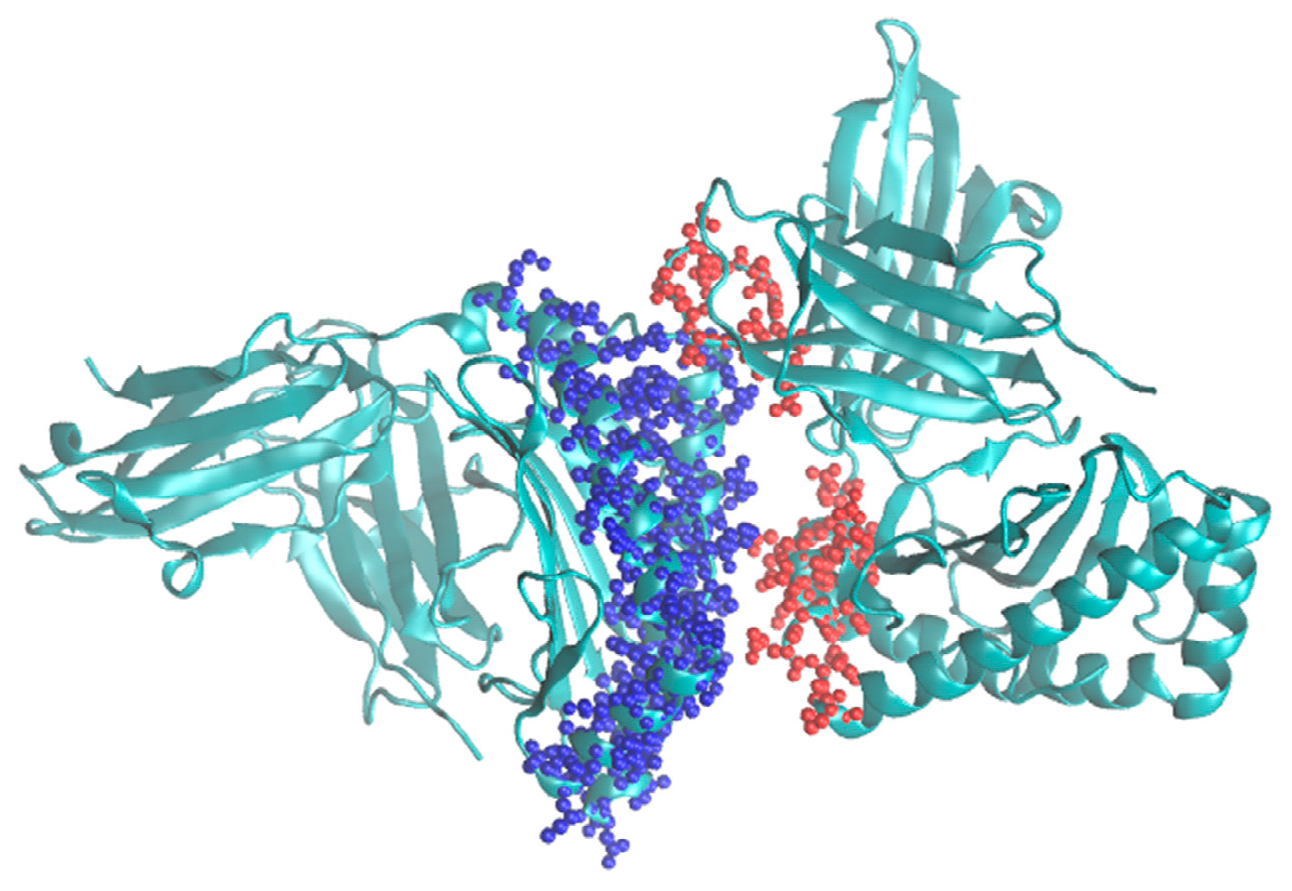
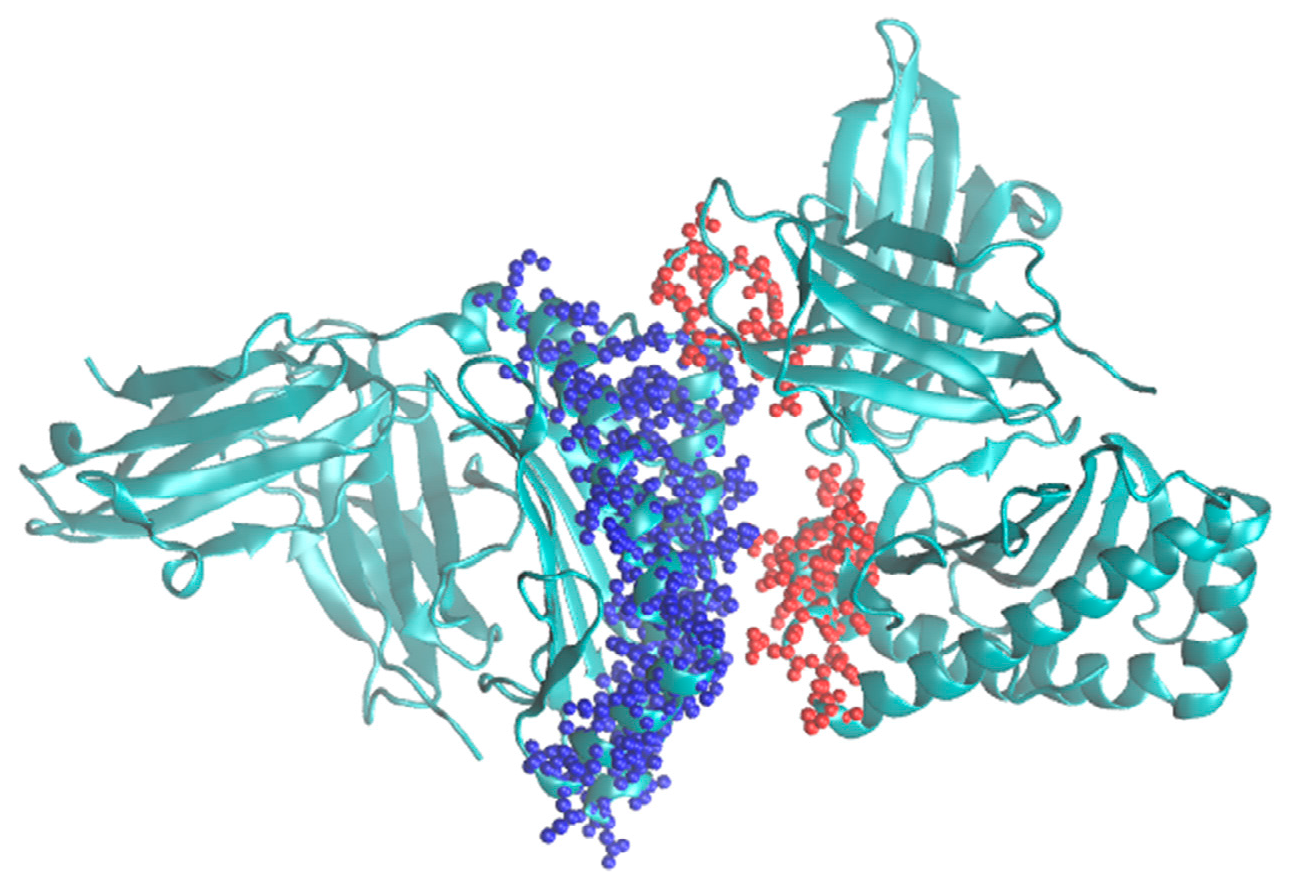
Thus, it is expected that this algorithm would significantly speed up the modeling of protein complexes where the interaction region is relatively small compared to the overall volume of the complex and foster further developments in the Monte Carlo methodology. It is planned to implement it into the program called MMC [
4] that already includes the single grid version. A typical example for its use would be modeling major histocompatibility complexes (MHC) [
8]. An example for such a system is shown in
Figure 3.



{kind=link}
{kind=link}
{kind=link}