

CD44 Receptor-Mediated Ferroptosis Induction by Hyaluronic Acid Carbon Quantum Dots in Triple-Negative Breast Cancer Cells Through Downregulation of SLC7A11 Pathway


 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. HA-CQDs Synthesis
2.3. Characterization Analysis
2.4. Molecular Docking Study
2.5. Anticancer Effect of HA CQDs
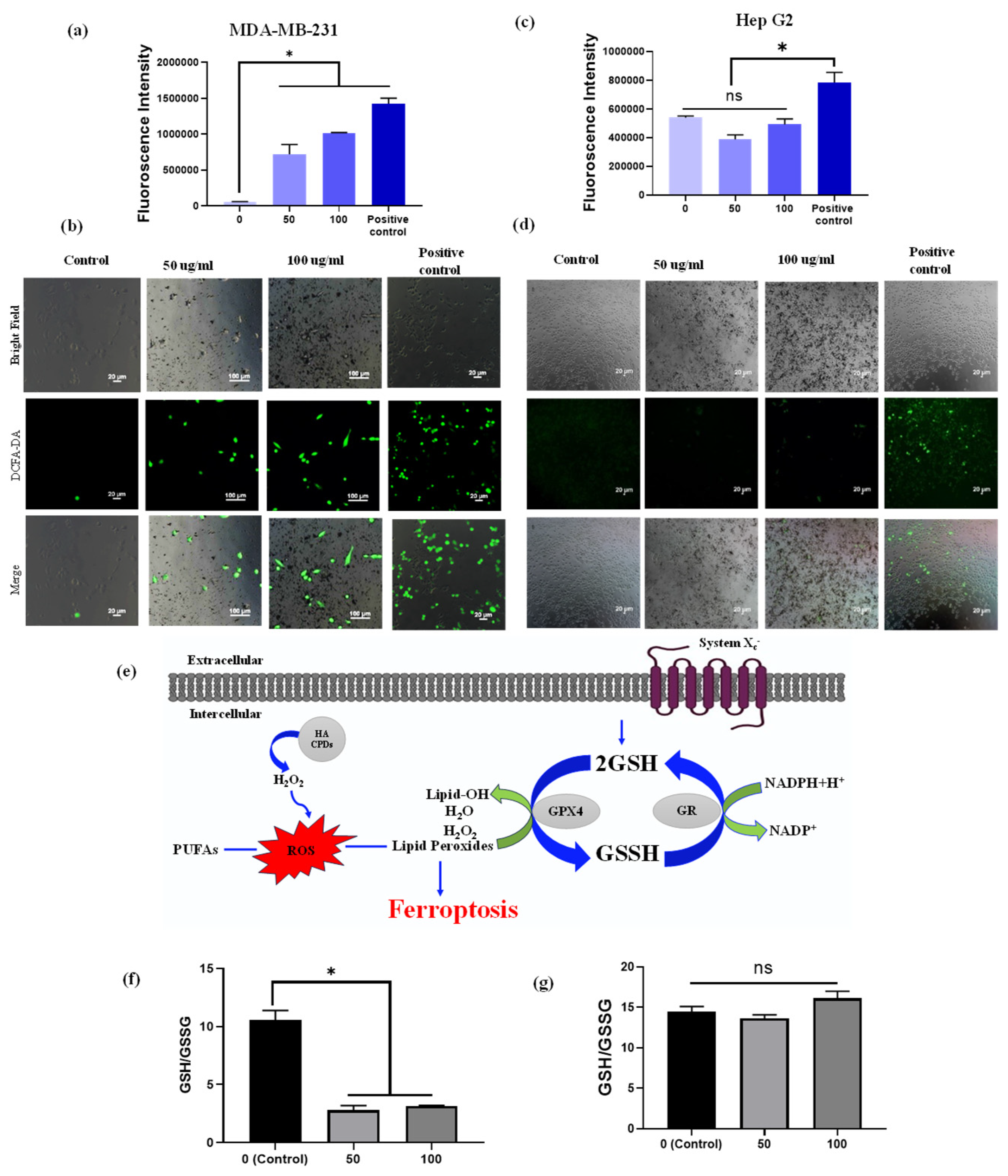
2.6. Intracellular ROS Generation
2.7. Ellman’s Analysis for Intracellular GSH Content
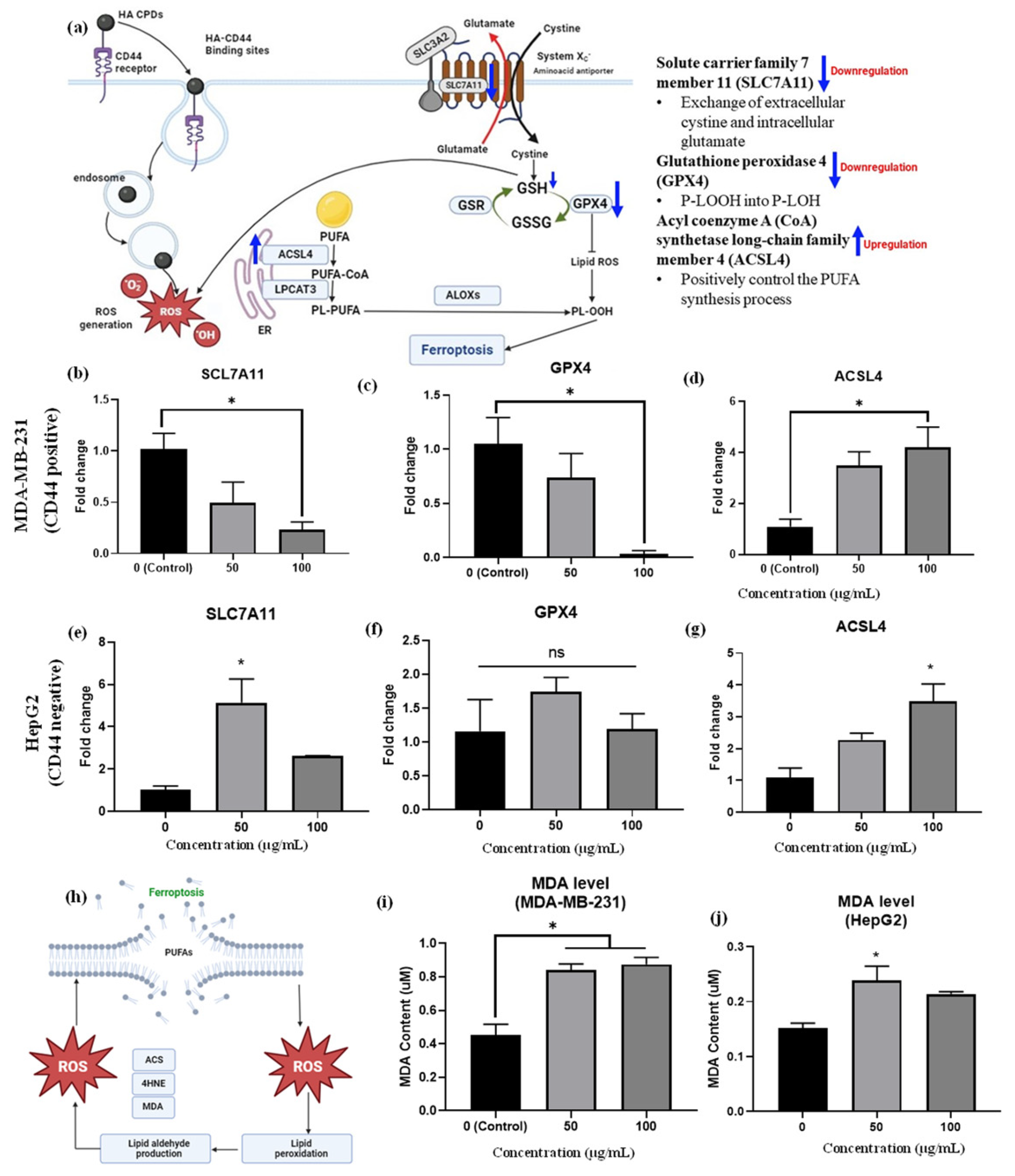
2.8. mRNA Expression of Ferroptosis Markers
2.9. Lipid Peroxidation
2.10. Statistical Analysis
3. Results and Discussion
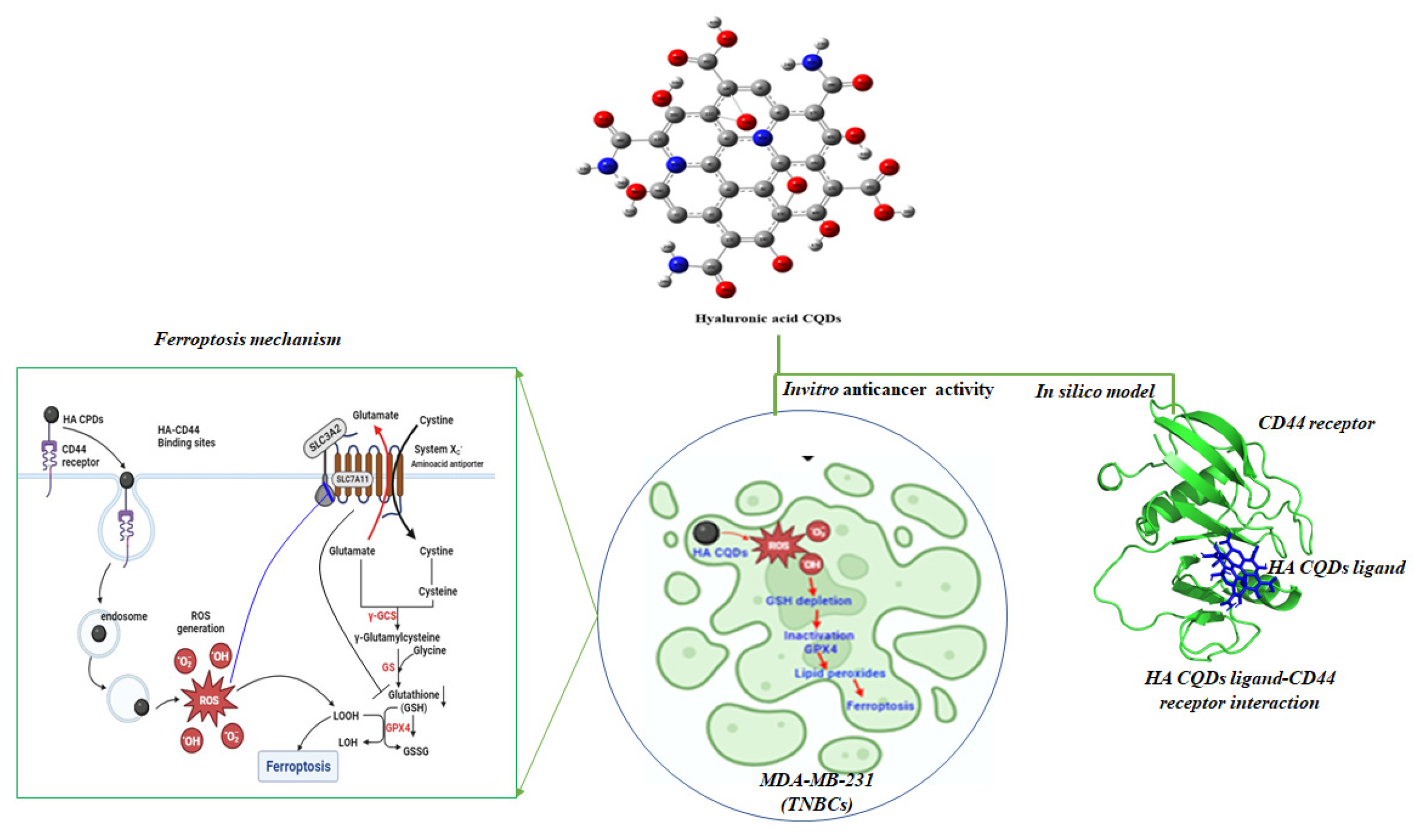
3.1. Formation Mechanism of HA Carbon Quantum Dots
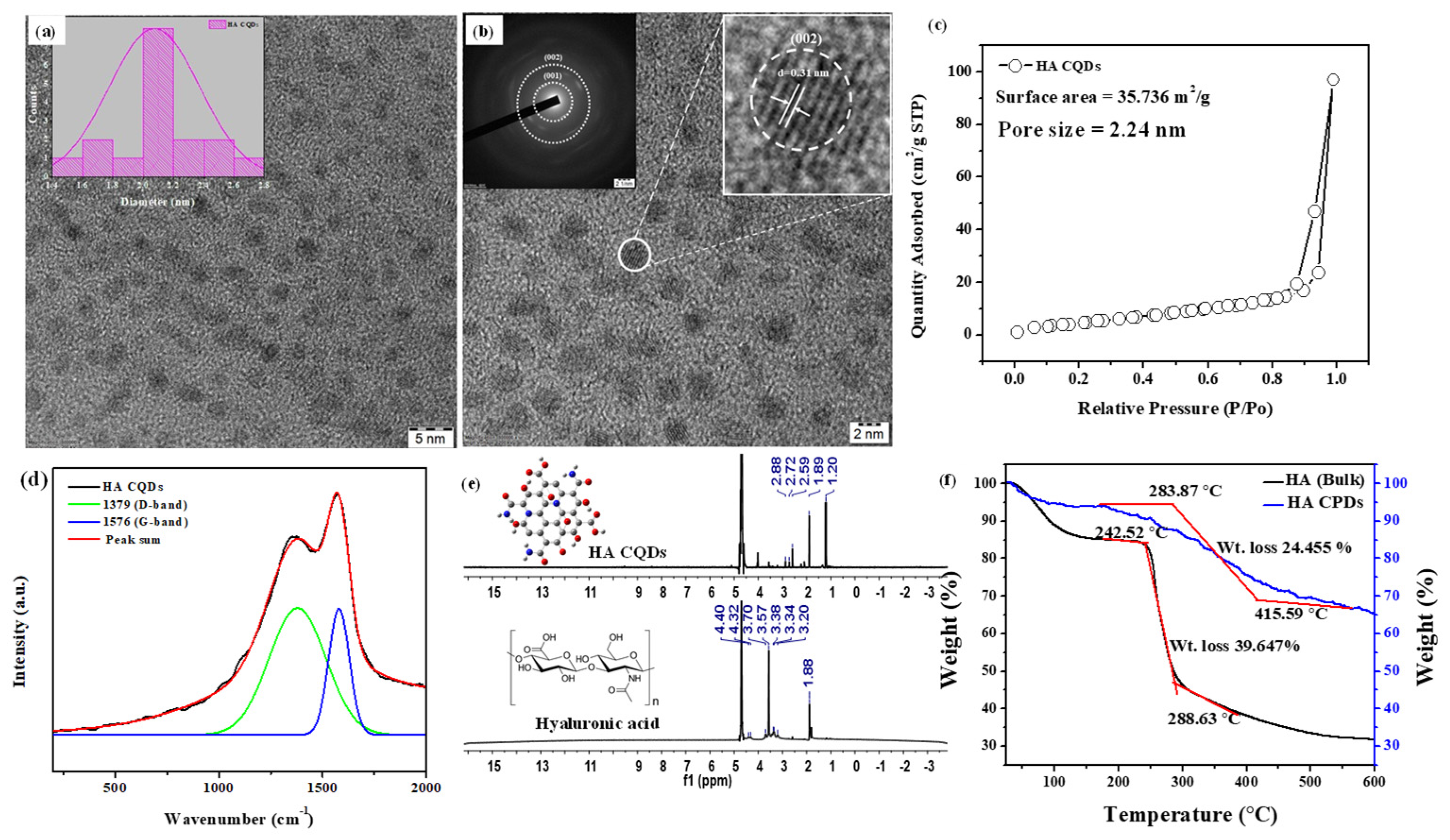
3.2. TEM and BET Analysis
3.3. Micro Raman Analysis, NMR, and TGA Analysis
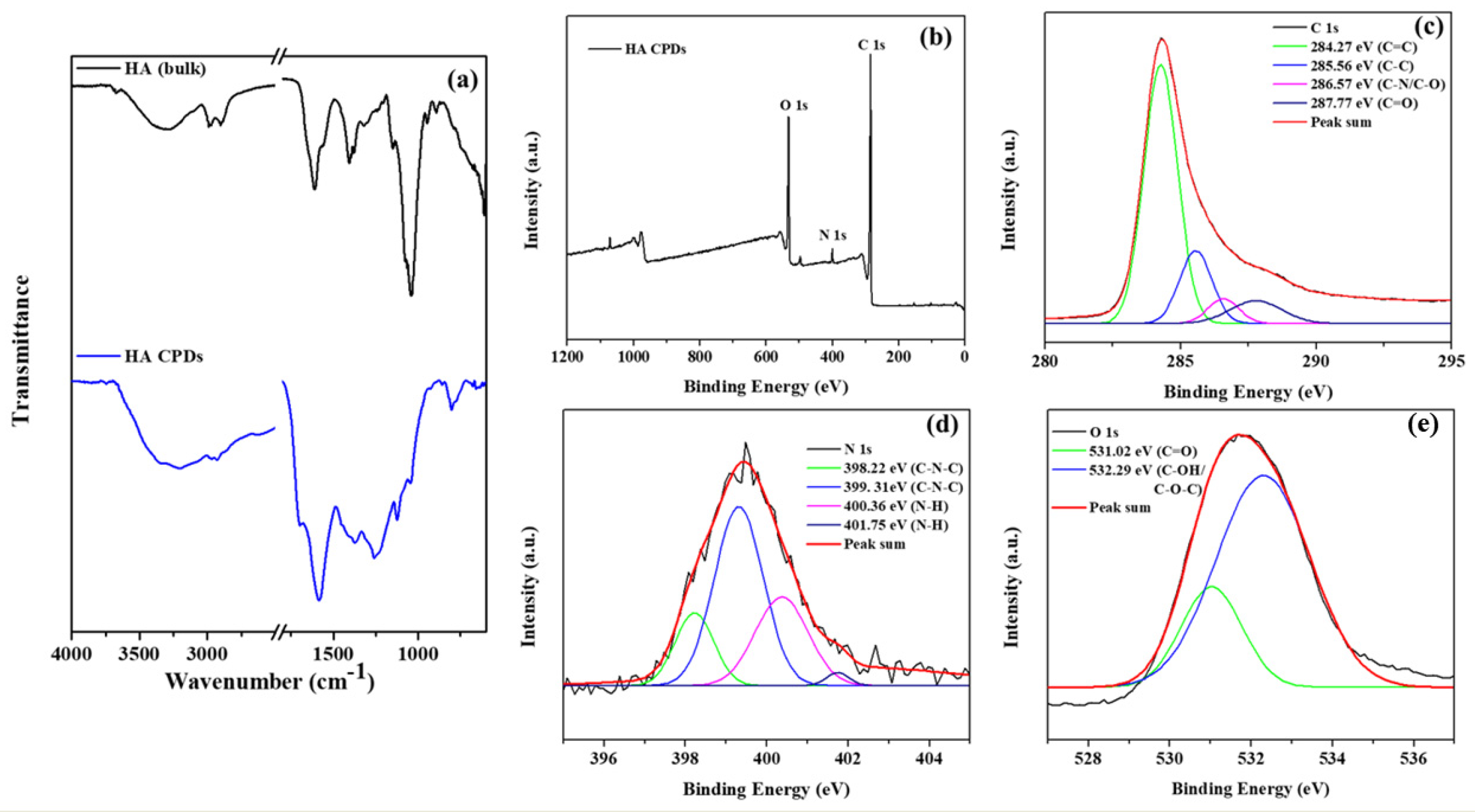
3.4. X-Ray Photoelectron Spectroscopy (XPS) Studies
3.5. Molecular Docking of HA and HA-CQDs
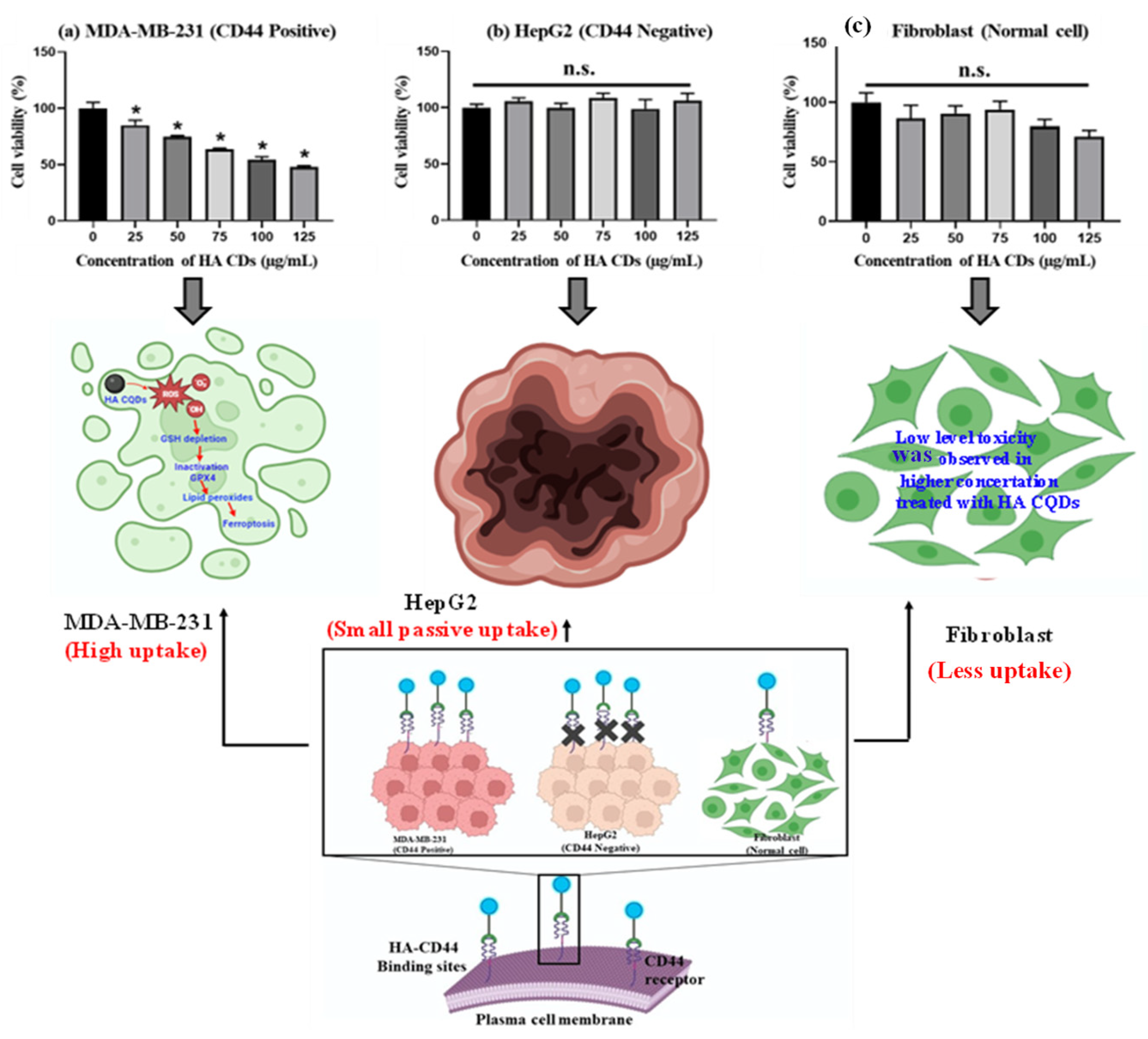
3.6. Biological Significance of HA System and HA-CD44 Binding Affinity
3.7. In Vitro Cytotoxicity
3.8. Detection of Intercellular ROS
3.9. GSH Depletion
3.10. mRNA Expression Profiles of Ferroptosis Markers and Lipid Peroxidation (MDA Assay)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No | Molecules | Target | Cancer Cells | Efficacy |
|---|---|---|---|---|
| 1 | CQD–HA–PEI@GNR–DOX | CD44 | Breast cancer cells (MCF7) [69] | 200 μg mL−1 |
| 2 | Verapamil-loaded hyaluronic acid-modified carbon quantum dots | CD44 | SH-Sy5y human cell line Neuro 2a mouse neuroblastoma cell line [70] | 3.75 μg/mL |
| 3 | Graphene quantum dot-hyaluronic acid nanocomposites | CD44 | Breast cancer (MCF-7) [71] | 500 µg/mL |
| 4 | HA CQDs | CD44 | Cervical cancer cells [72] | 100 μg/mL |
| 5 | HA-CQD@p-CBA-DOX | CD44 | Breast cancer cell line [73] | 90% |
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Armenta, D.A.; Dixon, S.J. Investigating nonapoptotic cell death using chemical biology approaches. Cell Chem. Biol. 2020, 27, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Shao, M.; Zhang, W.; Wang, F.; Wang, L.; Du, H. A Copper Silicate-Based Multifunctional Nanoplatform with Glutathione Depletion and Hypoxia Relief for Synergistic Photodynamic/Chemodynamic Therapy. Materials 2024, 17, 3495. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Sun, S.; Johnson, T.; Qi, R.; Zhang, S.; Zhang, J.; Yang, K. The glutathione peroxidase Gpx4 prevents lipid peroxidation and ferroptosis to sustain Treg cell activation and suppression of antitumor immunity. Cell Rep. 2021, 35, 109235. [Google Scholar] [CrossRef]
- Wu, L.; Xian, X.; Tan, Z.; Dong, F.; Xu, G.; Zhang, M.; Zhang, F. The role of iron metabolism, lipid metabolism, and redox homeostasis in Alzheimer’s disease: From the perspective of ferroptosis. Microsyst. Nanoeng. 2023, 60, 2832–2850. [Google Scholar] [CrossRef]
- Bao, Y.; Li, G.; Li, S.; Zhang, H.; Wu, X.; Yan, R.; Wang, Z.; Guo, C.; Jin, Y. Multifunctional tumor-targeting Carbon dots for Tumor Microenvironment activated ferroptosis and immunotherapy in Cancer Treatment. ACS Appl. Mater. Interfaces 2023, 15, 56834–56845. [Google Scholar] [CrossRef]
- Zhou, M.; Yang, Z.; Yin, T.; Zhao, Y.; Wang, C.-Y.; Zhu, G.-Y.; Bai, L.-P.; Jiang, Z.-H.; Zhang, W. Functionalized Fe-Doped Carbon Dots Exhibiting Dual Glutathione Consumption to Amplify Ferroptosis for Enhanced Cancer Therapy. ACS Appl. Mater. Interfaces 2023, 15, 53228–53241. [Google Scholar] [CrossRef]
- Saini, K.K.; Chaturvedi, P.; Sinha, A.; Singh, M.P.; Khan, M.A.; Verma, A.; Nengroo, M.A.; Satrusal, S.R.; Meena, S.; Singh, A. Loss of PERK function promotes ferroptosis by downregulating SLC7A11 (System Xc−) in colorectal cancer. Redox Biol. 2023, 65, 102833. [Google Scholar] [CrossRef]
- Xia, C.; Xing, X.; Zhang, W.; Wang, Y.; Jin, X.; Tian, M.; Ba, X.; Hao, F. Cysteine and homocysteine can be exploited by GPX4 in ferroptosis inhibition independent of GSH synthesis. Redox Biol. 2024, 69, 102999. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, Z.; Zhang, Y.; Ma, L.; Song, E.; Song, Y. “Iron free” zinc oxide nanoparticles with ion-leaking properties disrupt intracellular ROS and iron homeostasis to induce ferroptosis. Cell Death Dis. 2020, 11, 183. [Google Scholar] [CrossRef]
- Al-Hetty, H.R.A.K.; Jalil, A.T.; Al-Tamimi, J.H.Z.; Shakier, H.G.; Kandeel, M.; Saleh, M.M.; Naderifar, M. Engineering and surface modification of carbon quantum dots for cancer bioimaging. Inorg. Chem. Commun. 2023, 149, 110433. [Google Scholar] [CrossRef]
- Dutta, S.D.; Moniruzzaman, M.; Hexiu, J.; Sarkar, S.; Ganguly, K.; Patel, D.K.; Mondal, J.; Lee, Y.-K.; Acharya, R.; Kim, J. Polyphenolic Carbon Quantum Dots with Intrinsic Reactive Oxygen Species Amplification for Two-Photon Bioimaging and In Vivo Tumor Therapy. ACS Appl. Mater. Interfaces 2023, 15, 52083–52099. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Xu, M.; Sun, X.; Feliu, N.; Feng, L.; Parak, W.J.; Liu, S. Quantitative comparison of gold nanoparticle delivery via the enhanced permeation and retention (EPR) effect and mesenchymal stem cell (MSC)-based targeting. ACS Nano 2023, 17, 2039–2052. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yang, S.; Chen, L.; Li, Y.; He, P.; Wang, G.; Dong, H.; Ma, P.; Ding, G. Tumor diagnosis using carbon-based quantum dots: Detection based on the hallmarks of cancer. Bioact. Mater. 2024, 33, 174–222. [Google Scholar] [CrossRef] [PubMed]
- Karakoçak, B.B.M.; Laradji, A.; Primeau, T.; Berezin, M.Y.; Li, S.; Ravi, N. Hyaluronan-conjugated carbon quantum dots for bioimaging use. ACS Appl. Mater. Interfaces 2020, 13, 277–286. [Google Scholar]
- Responte, D.J.; Natoli, R.M.; Athanasiou, K.A. Identification of potential biophysical and molecular signalling mechanisms underlying hyaluronic acid enhancement of cartilage formation. J. R. Soc. Interface 2012, 9, 3564–3573. [Google Scholar] [CrossRef]
- Răcuciu, M.; Oancea, S.; Barbu-Tudoran, L.; Drăghici, O.; Agavriloaei, A.; Creangă, D. A Study of Hyaluronic Acid’s Theoretical Reactivity and of Magnetic Nanoparticles Capped with Hyaluronic Acid. Materials 2024, 17, 1229. [Google Scholar] [CrossRef]
- Curcio, F.; Sanguedolce, M.; Filice, L.; Testa, F.; Catapano, G.; Giordano, F.; Trombino, S.; Cassano, R. Hybrid Nanoparticles Based on Mesoporous Silica and Functionalized Biopolymers as Drug Carriers for Chemotherapeutic Agents. Materials 2024, 17, 3877. [Google Scholar] [CrossRef]
- Johnson, M.; Song, R.; Li, Y.; Milne, C.; Lyu, J.; Lara-Sáez, I.; Wang, W. Hyaluronic acid/chondroitin sulfate-based dynamic thiol–aldehyde addition hydrogel: An injectable, self-healing, on-demand dissolution wound dressing. Materials 2024, 17, 3003. [Google Scholar] [CrossRef]
- Ganguly, S.; Margel, S. Fluorescent quantum dots-based hydrogels: Synthesis, Fabrication and multimodal biosensing. Talanta Open 2023, 8, 100243. [Google Scholar]
- Zhang, M.; Fang, Z.; Zhao, X.; Niu, Y.; Lou, J.; Zhao, L.; Wu, Y.; Zou, S.; Du, F.; Shao, Q. Hyaluronic acid functionalized nitrogen-doped carbon quantum dots for targeted specific bioimaging. RSC Adv. 2016, 6, 104979–104984. [Google Scholar] [CrossRef]
- Mattheolabakis, G.; Milane, L.; Singh, A.; Amiji, M.M. Hyaluronic acid targeting of CD44 for cancer therapy: From receptor biology to nanomedicine. J. Drug Target. 2015, 23, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Khalfaoui, M.; Knani, S.; Hachicha, M.; Lamine, A.B. New theoretical expressions for the five adsorption type isotherms classified by BET based on statistical physics treatment. J. Colloid Interface Sci. 2003, 263, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Sridharan, M.; Kamaraj, P.; Arockiaselvi, J.; Pushpamalini, T.; Vivekanand, P.; Kumar, S.H. Synthesis, characterization and evaluation of biosynthesized Cerium oxide nanoparticle for its anticancer activity on breast cancer cell (MCF 7). Mater. Today Proc. 2021, 36, 914–919. [Google Scholar] [CrossRef]
- Groppo, E.; Bonino, F.; Cesano, F.; Damin, A.; Manzoli, M. Metal-Free Functionalized Carbons in Catalysis: Synthesis, Characterization and Applications, 1st ed.; Villa, A., Dimitratos, N., Eds.; United Kingdom by CPI Group (UK) Ltd.: Croydon, UK, 2018; pp. 103–137. [Google Scholar]
- Zhang, L.; Lin, Z.; Yu, Y.-X.; Jiang, B.-P.; Shen, X.-C. Multifunctional hyaluronic acid-derived carbon dots for self-targeted imaging-guided photodynamic therapy. J. Mater. Chem. B 2018, 6, 6534–6543. [Google Scholar] [CrossRef]
- Li, J.; Qiao, M.; Ji, Y.; Lin, L.; Zhang, X.; Linhardt, R.J. Chemical, enzymatic and biological synthesis of hyaluronic acids. Int. J. Biol. Macromol. 2020, 152, 199–206. [Google Scholar] [CrossRef]
- Misra, S.; Heldin, P.; Hascall, V.C.; Karamanos, N.K.; Skandalis, S.S.; Markwald, R.R.; Ghatak, S. Hyaluronan–CD44 interactions as potential targets for cancer therapy. FEBS J. 2011, 278, 1429–1443. [Google Scholar] [CrossRef]
- Alkrad, J.A.; Mrestani, Y.; Stroehl, D.; Wartewig, S.; Neubert, R. Characterization of enzymatically digested hyaluronic acid using NMR, Raman, IR, and UV–Vis spectroscopies. J. Pharm. Biomed. Anal. 2003, 31, 545–550. [Google Scholar] [CrossRef]
- Sciabica, S.; Tafuro, G.; Semenzato, A.; Traini, D.; Silva, D.M.; Reis, L.G.D.; Canilli, L.; Terno, M.; Durini, E.; Vertuani, S. Design, synthesis, characterization, and in vitro evaluation of a new cross-linked hyaluronic acid for pharmaceutical and cosmetic applications. Pharmaceutics 2021, 13, 1672. [Google Scholar] [CrossRef]
- Sargazi, A.; Shiri, F.; Keikha, S.; Majd, M.H. Hyaluronan magnetic nanoparticle for mitoxantrone delivery toward CD44-positive cancer cells. Colloids Surf. B. Biointerfaces 2018, 171, 150–158. [Google Scholar] [CrossRef]
- Platt, V.M.; Szoka, F.C., Jr. Anticancer therapeutics: Targeting macromolecules and nanocarriers to hyaluronan or CD44, a hyaluronan receptor. Mol. Pharm. 2008, 5, 474–486. [Google Scholar] [CrossRef]
- Karousou, E.; Misra, S.; Ghatak, S.; Dobra, K.; Götte, M.; Vigetti, D.; Passi, A.; Karamanos, N.K.; Skandalis, S.S. Roles and targeting of the HAS/hyaluronan/CD44 molecular system in cancer. Matrix Biol. 2017, 59, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Truica, C.I.; Wang, B.; Wang, Y.; Ren, X.; Harvey, H.A.; Song, J.; Yang, J.-M. Immunotherapy for triple-negative breast cancer: Existing challenges and exciting prospects. Drug Resist. Updates 2017, 32, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Abdoli Shadbad, M.; Hosseinkhani, N.; Asadzadeh, Z.; Derakhshani, A.; Karim Ahangar, N.; Hemmat, N.; Lotfinejad, P.; Brunetti, O.; Silvestris, N.; Baradaran, B. A systematic review to clarify the prognostic values of CD44 and CD44+ CD24-phenotype in triple-negative breast cancer patients: Lessons learned and the road ahead. Front. Oncol. 2021, 11, 689839. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, D.S.; Svechkarev, D.; Souchek, J.; Hill, T.K.; Taylor, M.; Natarajan, A.; Mohs, A.M. Impact of structurally modifying hyaluronic acid on CD44 interaction. J. Mater. Chem. B 2017, 5, 8183–8192. [Google Scholar] [CrossRef]
- Amorim, S.; Reis, C.A.; Reis, R.L.; Pires, R.A. Extracellular matrix mimics using hyaluronan-based biomaterials. Trends Biotechnol. 2021, 39, 90–104. [Google Scholar] [CrossRef]
- Stern, R.; Jedrzejas, M.J. Carbohydrate polymers at the center of life’s origins: The importance of molecular processivity. Chem. Rev. 2008, 108, 5061–5085. [Google Scholar] [CrossRef]
- Cai, J.; Fu, J.; Li, R.; Zhang, F.; Ling, G.; Zhang, P. A potential carrier for anti-tumor targeted delivery-hyaluronic acid nanoparticles. Carbohydr. Polym. 2019, 208, 356–364. [Google Scholar] [CrossRef]
- Kim, H.; Park, Y.; Stevens, M.M.; Kwon, W.; Hahn, S.K. Multifunctional hyaluronate–nanoparticle hybrid systems for diagnostic, therapeutic and theranostic applications. J. Control. Release 2019, 303, 55–66. [Google Scholar] [CrossRef]
- Misra, S.; Hascall, V.C.; Markwald, R.R.; Ghatak, S. Interactions between hyaluronan and its receptors (CD44, RHAMM) regulate the activities of inflammation and cancer. Front. Immunol. 2015, 6, 201. [Google Scholar] [CrossRef]
- Mohkam, M.; Sadraeian, M.; Lauto, A.; Gholami, A.; Nabavizadeh, S.H.; Esmaeilzadeh, H.; Alyasin, S. Exploring the potential and safety of quantum dots in allergy diagnostics. Microsyst. Nanoeng. 2023, 9, 145. [Google Scholar] [CrossRef]
- Yao, L.; Zhao, M.-M.; Luo, Q.-W.; Zhang, Y.-C.; Liu, T.-T.; Yang, Z.; Liao, M.; Tu, P.; Zeng, K.-W. Carbon quantum dots-based nanozyme from coffee induces cancer cell ferroptosis to activate antitumor immunity. ACS Nano 2022, 16, 9228–9239. [Google Scholar] [CrossRef] [PubMed]
- Bagayoko, S.; Meunier, E. Emerging roles of ferroptosis in infectious diseases. FEBS J. 2022, 289, 7869–7890. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.J.; Tang, D.; Wang, Y.C.; Li, X.; Bao, H.; Tang, C.B.; Dong, X.J.; Li, X.N.; Yang, Q.X.; Yan, Y.; et al. The mechanism of ferroptosis and its related diseases. Mol. Biomed. 2023, 4, 33. [Google Scholar]
- Mo, Y.; Zou, Z.; Chen, E. Targeting ferroptosis in hepatocellular carcinoma. Hepatol. Int. 2024, 18, 32–49. [Google Scholar] [CrossRef]
- Conrad, M.; Sato, H. The oxidative stress-inducible cystine/glutamate antiporter, system xc−: Cystine supplier and beyond. Amino Acids 2012, 42, 231–246. [Google Scholar] [CrossRef]
- Koppula, P.; Zhuang, L.; Gan, B.Y. Cystine transporter SLC7A11/xCT in cancer: Ferroptosis, nutrient dependency, and cancer therapy. Protein Cell 2021, 12, 599–620. [Google Scholar] [CrossRef]
- Maqbool, M.; Bekele, F.; Fekadu, G. Treatment strategies against triple-negative breast cancer: An updated review. Breast Cancer 2023, 14, 15–24. [Google Scholar] [CrossRef]
- Fu, W.; Sun, H.; Zhao, Y.; Chen, M.; Yang, L.; Yang, X.; Jin, W. Targeted delivery of CD44s-siRNA by ScFv overcomes de novo resistance to cetuximab in triple negative breast cancer. Mol. Immunol. 2018, 99, 124–133. [Google Scholar] [CrossRef]
- Ponta, H.; Sherman, L.; Herrlich, P.A. CD44: From adhesion molecules to signalling regulators. Nat. Rev. Mol. Cell Biol. 2003, 4, 33–45. [Google Scholar] [CrossRef]
- Sahar, S.; Sun, S.; Zeb, A.; Jin, L.; Gao, Y.; Tian, L.; Wang, W.; Xu, A.-W.; Mao, Z. Multi-enzyme mimics–cracking the code of subcellular cascade reactions and their potential biological applications. Mater. Chem. Front. 2023, 7, 3037–3072. [Google Scholar] [CrossRef]
- Zheng, G.; Zhang, J.; Zhang, X.; Zhang, Z.; Liu, S.; Zhang, S.; Zhang, C. Implications of ferroptosis in silver nanoparticle-induced cytotoxicity of macrophages. Ecotoxicol. Environ. Saf. 2023, 259, 115057. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Ruan, L.; Zhou, S.; Wu, L.; Cao, J.; Qi, X.; Zhang, X.; Shen, S. Appropriate size of Fe3O4 nanoparticles for cancer therapy by ferroptosis. ACS Appl. Bio Mater. 2022, 5, 1692–1699. [Google Scholar] [CrossRef] [PubMed]
- D’arcy, M.S. Cell death: A review of the major forms of apoptosis, necrosis and autophagy. Cell Biol. Int. 2019, 43, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Chaabane, W.; User, S.D.; El-Gazzah, M.; Jaksik, R.; Sajjadi, E.; Rzeszowska-Wolny, J.; Łos, M.J. Autophagy, apoptosis, mitoptosis and necrosis: Interdependence between those pathways and effects on cancer. Arch. Immunol. Ther. Exp. 2013, 61, 43–58. [Google Scholar] [CrossRef]
- Lee, Y.-S.; Lee, D.-H.; Choudry, H.A.; Bartlett, D.L.; Lee, Y.J. Ferroptosis-induced endoplasmic reticulum stress: Cross-talk between ferroptosis and apoptosis. Mol. Cancer Res. 2018, 16, 1073–1076. [Google Scholar] [CrossRef]
- Forcina, G.C.; Dixon, S.J. GPX4 at the crossroads of lipid homeostasis and ferroptosis. Proteomics 2019, 19, 1800311. [Google Scholar] [CrossRef]
- Zhou, B.; Liu, J.; Kang, R.; Klionsky, D.J.; Kroemer, G.; Tang, D. Ferroptosis is a type of autophagy-dependent cell death. Semin. Cancer Biol. 2020, 66, 89–100. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Singh, S.P.; Häder, D.-P.; Sinha, R.P. Detection of reactive oxygen species (ROS) by the oxidant-sensing probe 2′, 7′-dichlorodihydrofluorescein diacetate in the cyanobacterium Anabaena variabilis PCC 7937. Biochem. Biophys. Res. Commun. 2010, 397, 603–607. [Google Scholar] [CrossRef]
- Bartosz, G. Use of spectroscopic probes for detection of reactive oxygen species. Clin. Chim. Acta 2006, 368, 53–76. [Google Scholar] [CrossRef]
- Yu, D.; Zha, Y.; Zhong, Z.; Ruan, Y.; Li, Z.; Sun, L.; Hou, S. Improved detection of reactive oxygen species by DCFH-DA: New insight into self-amplification of fluorescence signal by light irradiation. Sens. Actuators B Chem. 2021, 339, 129878. [Google Scholar] [CrossRef]
- Schafer, F.Q.; Buettner, G.R. Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple. Free Radic. Biol. Med. 2001, 30, 1191–1212. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Liang, X.; Liu, X.; Li, Y.; Wang, Y.; Kong, L.; Tang, M. Induction of ferroptosis in response to graphene quantum dots through mitochondrial oxidative stress in microglia. Part. Fibre Toxicol. 2020, 17, 30. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Lin, X.; Huang, C. Activation of the reverse transsulfuration pathway through NRF2/CBS confers erastin-induced ferroptosis resistance. Br. J. Cancer 2020, 122, 279–292. [Google Scholar] [CrossRef]
- He, J.; Wang, X.; Chen, K.; Zhang, M.; Wang, J. The amino acid transporter SLC7A11-mediated crosstalk implicated in cancer therapy and the tumor microenvironment. Biochem. Pharmacol. 2022, 205, 115241. [Google Scholar] [CrossRef]
- Liu, X.; Chen, C.; Han, D.; Zhou, W.; Cui, Y.; Tang, X.; Xiao, C.; Wang, Y.; Gao, Y. SLC7A11/GPX4 inactivation-mediated ferroptosis contributes to the pathogenesis of triptolide-induced cardiotoxicity. Oxid. Med. Cell. Longev. 2022, 2022, 3192607. [Google Scholar] [CrossRef]
- Tang, Y.; Zhou, J.; Hooi, S.C.; Jiang, Y.-M.; Lu, G.-D. Fatty acid activation in carcinogenesis and cancer development: Essential roles of long-chain acyl-CoA synthetases. Oncol. Lett. 2018, 16, 1390–1396. [Google Scholar] [CrossRef]
- Lin, Z.; Liu, J.; Kang, R.; Yang, M.; Tang, D. Lipid metabolism in ferroptosis. Adv. Biol. 2021, 5, 2100396. [Google Scholar] [CrossRef]
- Khoshnood, A.; Farhadian, N.; Abnous, K.; Matin, M.M.; Ziaee, N.; Iranpour, S. Polyethyleneimine/gold nanorods conjugated with carbon quantum dots and hyaluronic acid for chemo-photothermal therapy of breast cancer. J. Mater. Chem. B 2025, 13, 4893–4909. [Google Scholar] [CrossRef]
- Mosalam, E.M.; Abdel-Bar, H.M.; Elberri, A.I.; Abdallah, M.S.; Zidan, A.A.A.; Batakoushy, H.A.; Mansour, H.E.A. Enhanced neuroprotective effect of verapamil-loaded hyaluronic acid modified carbon quantum dots in an in-vitro model of amyloid-induced Alzheimer’s disease. Int. J. Biol. Macromol. 2024, 275, 133742. [Google Scholar] [CrossRef]
- Soltani, M.; Ahmadzadeh, N.; Rajabi, S.; Besharati, N.; Khatamian, N.; Homayouni Tabrizi, M. Efficacy of graphene quantum dot-hyaluronic acid nanocomposites containing quinoline for target therapy against cancer cells. Sci. Rep. 2025, 15, 8494. [Google Scholar] [CrossRef]
- Aung, Y.-Y.; Wibrianto, A.; Sianturi, J.S.; Ulfa, D.K.; Sakti, S.C.W.; Irzaman, I.; Yuliarto, B.; Chang, J.-Y.; Kwee, Y.; Fahmi, M.Z. Comparison direct synthesis of hyaluronic acid-based carbon nanodots as dual active targeting and imaging of HeLa cancer cells. ACS Omega 2021, 6, 13300–13309. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, M.; Tian, L.; Qiu, Y.; Yu, Q.; Wang, X.; Guo, R.; He, Q. Facile strategy by hyaluronic acid functional carbon dot-doxorubicin nanoparticles for CD44 targeted drug delivery and enhanced breast cancer therapy. Int. J. Pharm. 2020, 578, 119122. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chandrasekaran, K.; Lee, C.E.; Yun, S.; Jangid, A.K.; Kim, S.; Kim, K. CD44 Receptor-Mediated Ferroptosis Induction by Hyaluronic Acid Carbon Quantum Dots in Triple-Negative Breast Cancer Cells Through Downregulation of SLC7A11 Pathway. Materials 2025, 18, 2139. https://doi.org/10.3390/ma18092139
Chandrasekaran K, Lee CE, Yun S, Jangid AK, Kim S, Kim K. CD44 Receptor-Mediated Ferroptosis Induction by Hyaluronic Acid Carbon Quantum Dots in Triple-Negative Breast Cancer Cells Through Downregulation of SLC7A11 Pathway. Materials. 2025; 18(9):2139. https://doi.org/10.3390/ma18092139
Chicago/Turabian StyleChandrasekaran, Karthikeyan, Chae Eun Lee, Seojeong Yun, Ashok Kumar Jangid, Sungjun Kim, and Kyobum Kim. 2025. "CD44 Receptor-Mediated Ferroptosis Induction by Hyaluronic Acid Carbon Quantum Dots in Triple-Negative Breast Cancer Cells Through Downregulation of SLC7A11 Pathway" Materials 18, no. 9: 2139. https://doi.org/10.3390/ma18092139
APA StyleChandrasekaran, K., Lee, C. E., Yun, S., Jangid, A. K., Kim, S., & Kim, K. (2025). CD44 Receptor-Mediated Ferroptosis Induction by Hyaluronic Acid Carbon Quantum Dots in Triple-Negative Breast Cancer Cells Through Downregulation of SLC7A11 Pathway. Materials, 18(9), 2139. https://doi.org/10.3390/ma18092139