
Proteomics and EPS Compositional Analysis Reveals Desulfovibrio bisertensis SY-1 Induced Corrosion on Q235 Steel by Biofilm Formation

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Microorganism, Medium, and Cultivation
2.2. EPS Extraction, Quantification, Analysis, Staining, and Observation
2.3. Biofilm Observation on the Q235 Surface
2.4. Proteome Analysis
3. Results
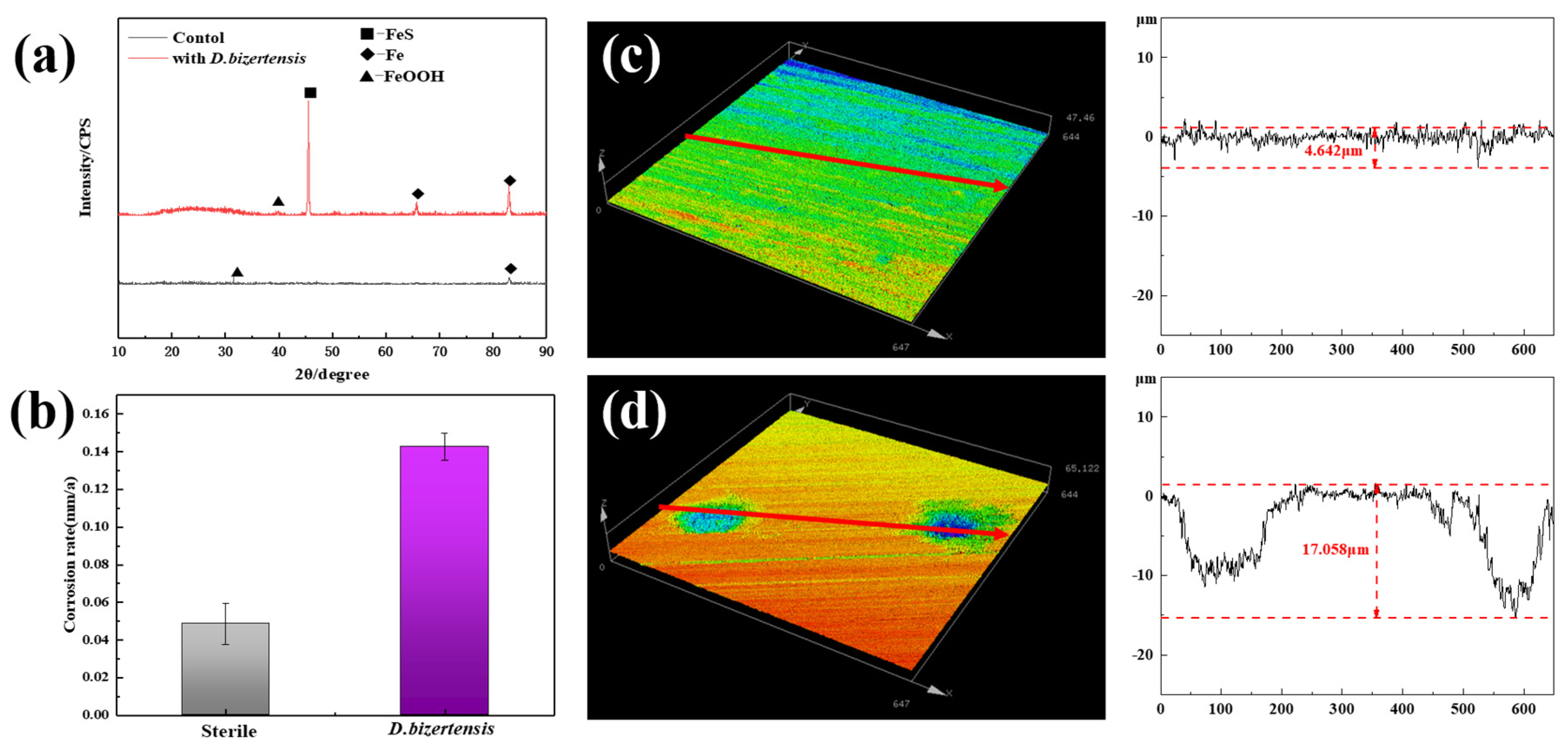
3.1. Biofilm Morphology and Growth, and the Composition Analysis of D. bizertensis on the Q235 Surface
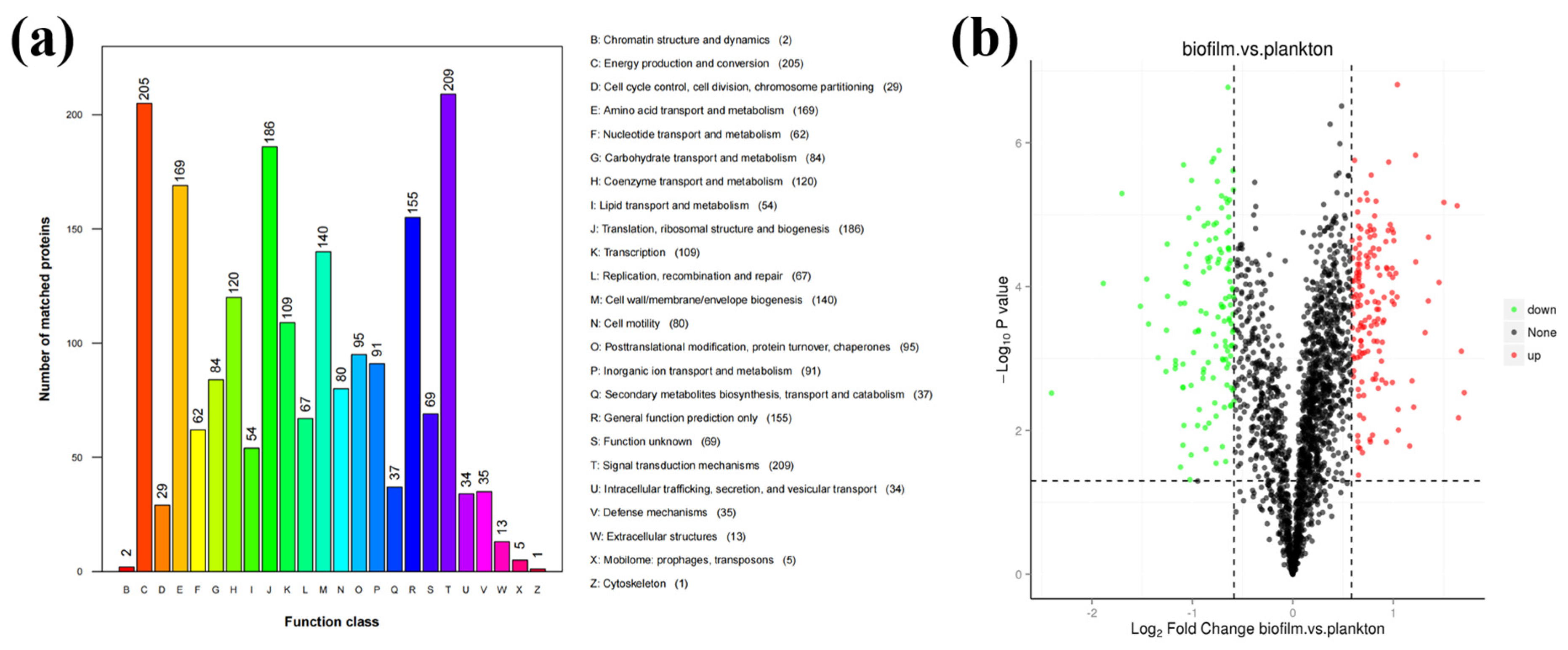
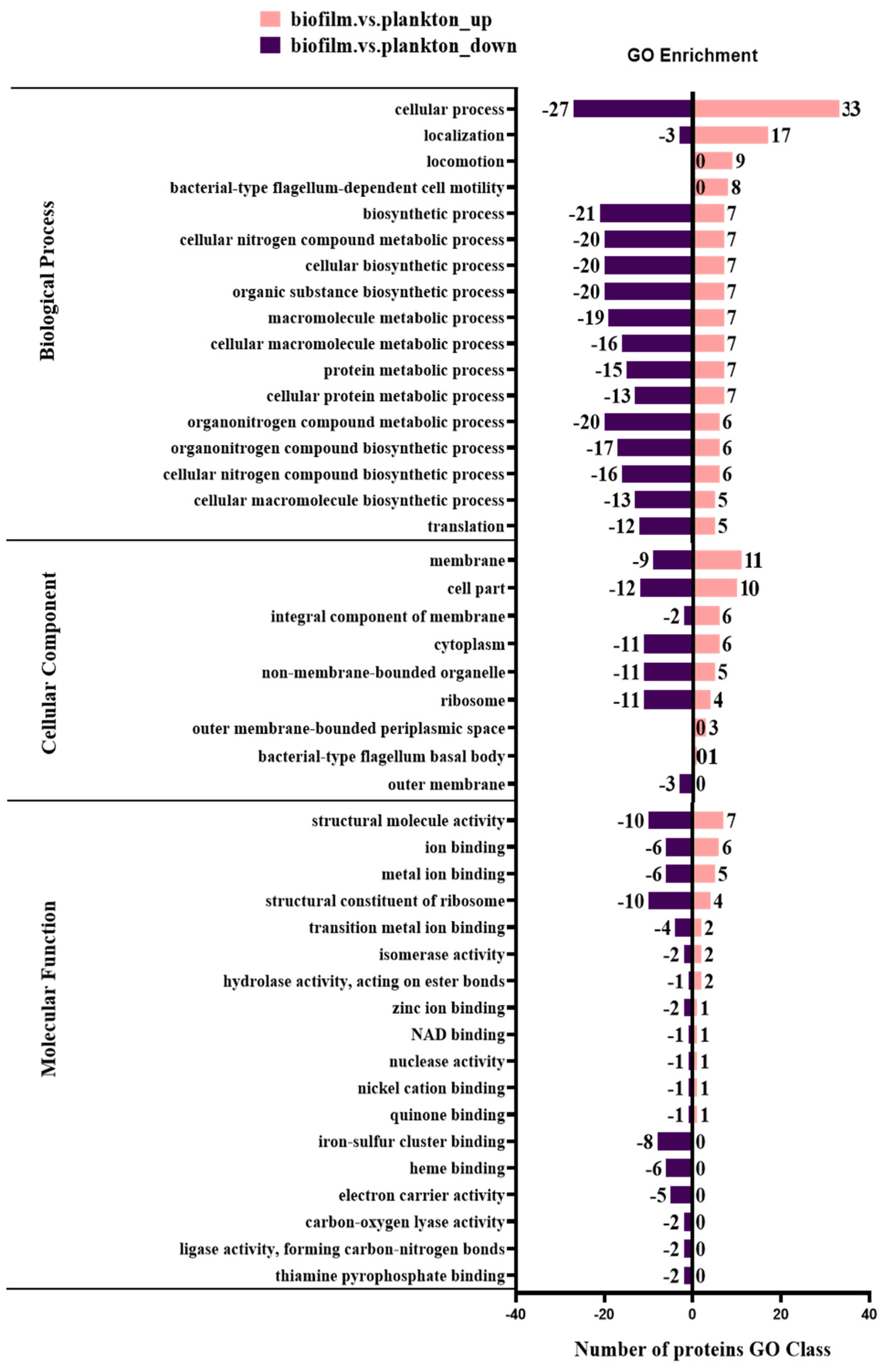
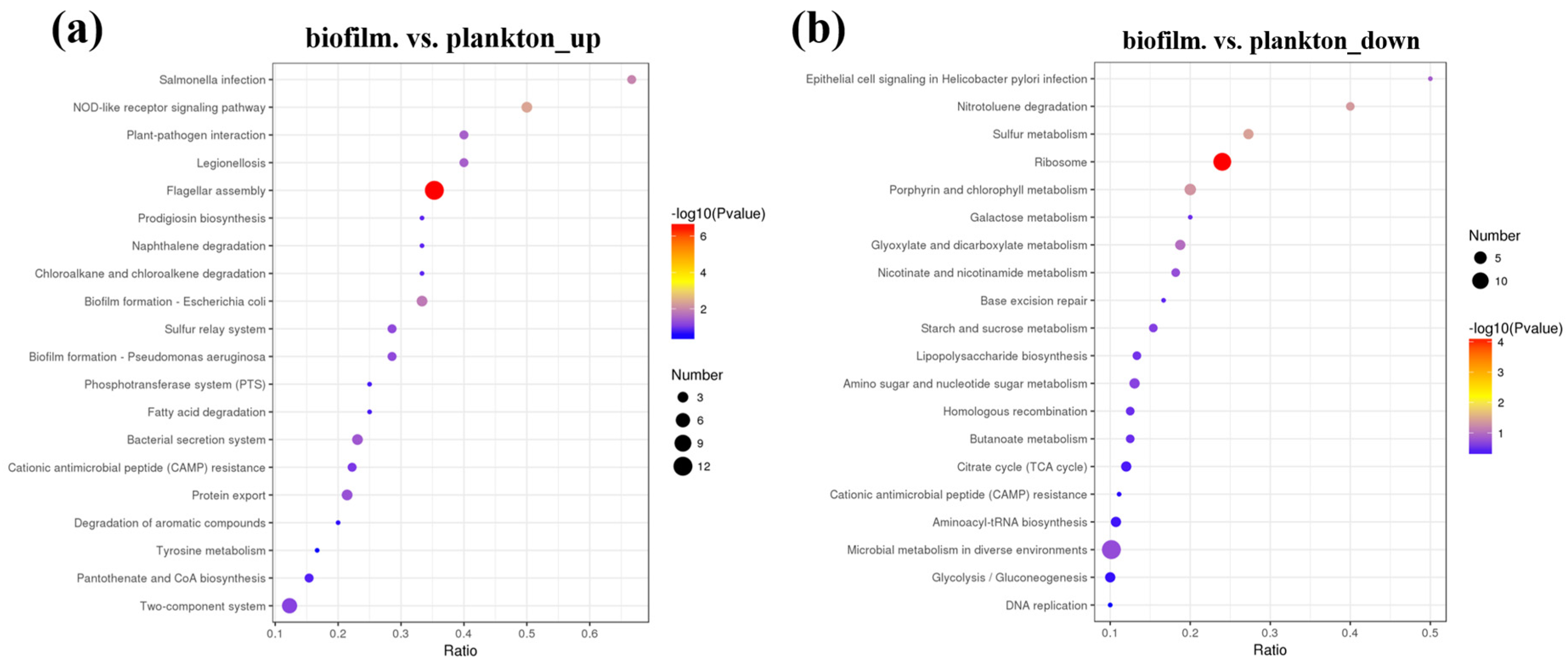
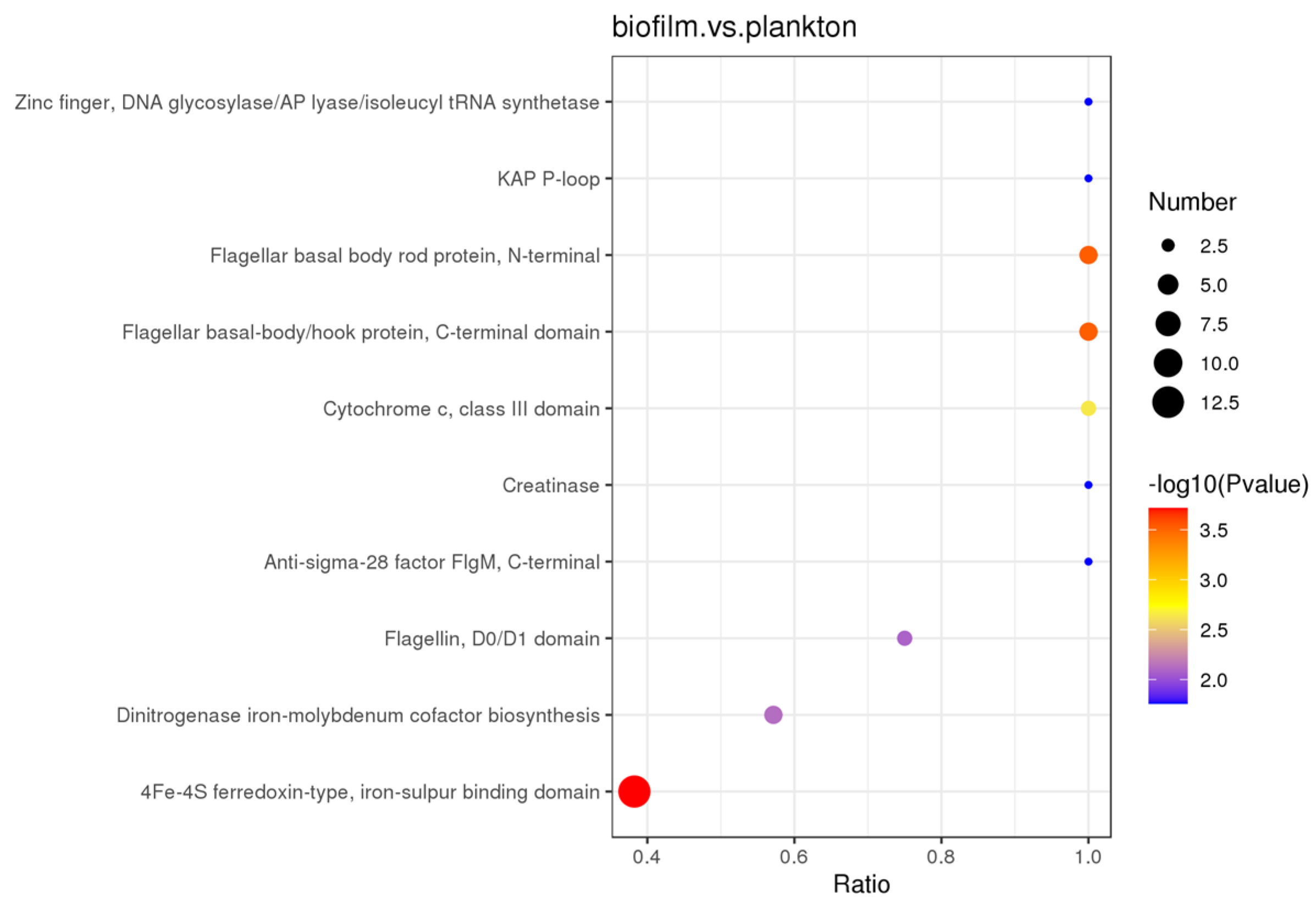
3.2. Identification and Comparison of Proteins
3.3. Changes in EPS during Biofilm Formation on the Q235 Surface
4. Discussion
4.1. D. bizertensis Motility Change
4.2. Protein Secretion and Transport in Cells
4.3. Survival and Electron Transfer Components of Biofilm Cells
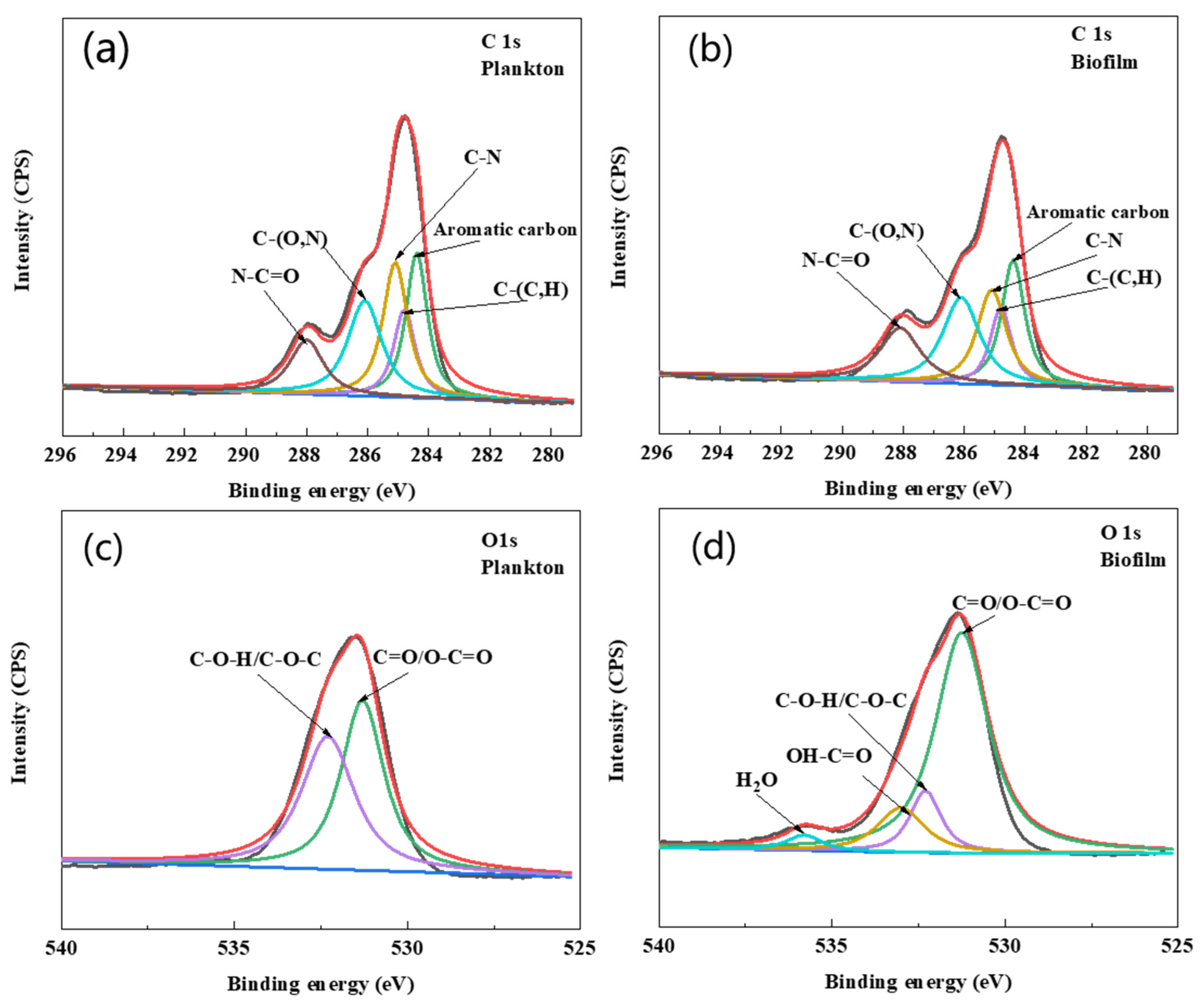
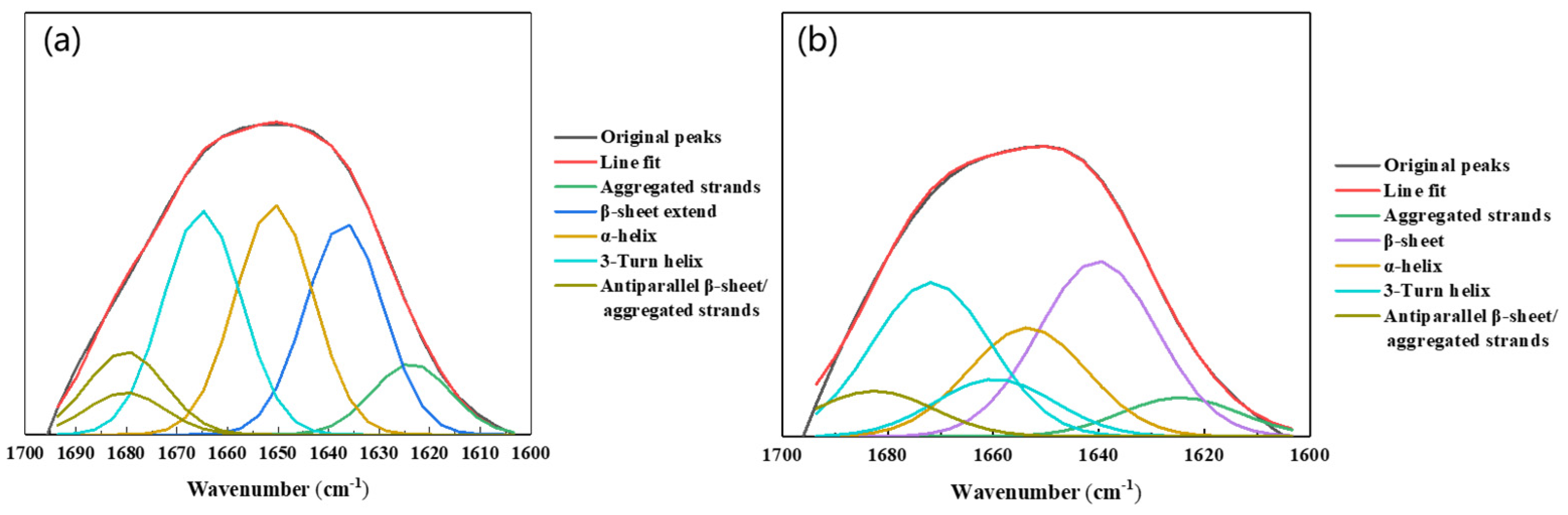
4.4. EPS Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Beech, I.B.; Cheung, C.S.; Chan, C.P.; Hill, M.A.; Franco, R.; Lino, A.-R. Study of parameters implicated in the biodeterioration of mild steel in the presence of different species of sulphate-reducing bacteria. Int. Biodeterior. Biodegrad. 1994, 34, 289–303. [Google Scholar] [CrossRef]
- Dinh, H.T.; Kuever, J.; Mussmann, M.; Hassel, A.W.; Stratmann, M.; Widdel, F. Iron corrosion by novel anaerobic microorganisms. Nature 2004, 427, 829–832. [Google Scholar] [CrossRef]
- Lewandowski, Z.; Beyenal, H. Mechanisms of Microbially Influenced Corrosion. In Marine and Industrial Biofoulin; Flemming, H.C., Murthy, P.S., Venkatesan, R., Cooksey, K., Eds.; Springer: Berlin, Germany, 2009. [Google Scholar]
- Videla, H.A.; Characklis, W.G. Biofouling and microbially influenced corrosion. Int. Biodeterior. Biodegrad. 1992, 29, 195–212. [Google Scholar] [CrossRef]
- Procópio, L. The role of biofilms in the corrosion of steel in marine environments. World J. Microbiol. Biotechnol. 2019, 35, 73. [Google Scholar] [CrossRef]
- Li, J.; Liu, X.; Zhang, J.; Zhang, R.; Wang, M.; Sand, W.; Duan, J.; Zhu, Q.; Zhai, S.; Hou, B. Effects of Inorganic Metabolites of Sulphate-Reducing Bacteria on the Corrosion of AZ31B and AZ63B Magnesium Alloy in 3.5 wt.% NaCl Solution. Materials 2022, 15, 2212. [Google Scholar] [CrossRef]
- Enning, D.; Garrelfs, J. Corrosion of iron by sulfate-reducing bacteria: New views of an old problem. Appl. Environ. Microbiol. 2014, 80, 1226–1236. [Google Scholar] [CrossRef]
- Enning, D.; Venzlaff, H.; Garrelfs, J.; Dinh, H.T.; Meyer, V.; Mayrhofer, K.; Hassel, A.W.; Stratmann, M.; Widdel, F. Marine sulfate-reducing bacteria cause serious corrosion of iron under electroconductive biogenic mineral crust. Environ. Microbiol. 2012, 14, 1772–1787. [Google Scholar] [CrossRef]
- Dou, W.; Jia, R.; Jin, P.; Liu, J.; Chen, S.; Gu, T. Investigation of the mechanism and characteristics of copper corrosion by sulfate reducing bacteria. Corros. Sci. 2018, 144, 237–248. [Google Scholar] [CrossRef]
- Jia, R.; Tan, J.L.; Jin, P.; Blackwood, D.J.; Xu, D.; Gu, T. Effects of biogenic H2S on the microbiologically influenced corrosion of C1018 carbon steel by sulfate reducing Desulfovibrio vulgaris biofilm. Corros. Sci. 2018, 130, 1–11. [Google Scholar] [CrossRef]
- Zhang, P.; Xu, D.; Li, Y.; Yang, K.; Gu, T. Electron mediators accelerate the microbiologically influenced corrosion of 304 stainless steel by the Desulfovibrio vulgaris biofilm. Bioelectrochemistry 2015, 101, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Gu, T.; Wang, D.; Lekbach, Y.; Xu, D. Extracellular electron transfer in microbial biocorrosion. Curr. Opin. Electrochem. 2021, 29, 100763. [Google Scholar] [CrossRef]
- Xu, D.; Gu, T. Carbon source starvation triggered more aggressive corrosion against carbon steel by the Desulfovibrio vulgaris biofilm. Int. Biodeterior. Biodegrad. 2014, 91, 74–81. [Google Scholar] [CrossRef]
- Xu, D.; Gu, T.; Lovley, D.R. Microbially mediated metal corrosion. Nat. Rev. Microbiol. 2023, 21, 705–718. [Google Scholar] [CrossRef]
- Zhou, E.; Li, F.; Zhang, D.; Xu, D.; Li, Z.; Jia, R.; Jin, Y.; Song, H.; Li, H.; Wang, Q. Direct microbial electron uptake as a mechanism for stainless steel corrosion in aerobic environments. Water Res. 2022, 219, 118553. [Google Scholar] [CrossRef]
- Alrammah, F.; Xu, L.; Patel, N.; Kontis, N.; Rosado, A.; Gu, T. Conductive magnetic nanowires accelerated electron transfer between C1020 carbon steel and Desulfovibrio vulgaris biofilm. Sci. Total Environ. 2024, 925, 171763. [Google Scholar] [CrossRef]
- Kato, S. Microbial extracellular electron transfer and its relevance to iron corrosion. Microb. Biotechnol. 2016, 9, 141–148. [Google Scholar] [CrossRef]
- Zhang, T.; Xu, Z.; Wan, H.; Chen, X.; Liu, H. Dual corrosion promotion of pipeline steel in sea mud induced by sulfate reducing bacteria: Bacteria concentration cell and electronic conduction of the biofilms covered sand grains. Corros. Sci. 2024, 232, 112005. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wingender, J. Relevance of microbial extracellular polymeric substances (EPSs)-Part I: Structural and ecological aspects. Water Sci. Technol. 2001, 43, 1–8. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Beech, I.B.; Sunner, J.A.; Hiraoka, K. Microbe-surface interactions in biofouling and biocorrosion processes. Int. Microbiol. 2005, 8, 157–168. [Google Scholar]
- Clark, M.E.; Edelmann, R.E.; Duley, M.L.; Wall, J.D.; Fields, M.W. Biofilm formation in Desulfovibrio vulgaris Hildenborough is dependent upon protein filaments. Environ. Microbiol. 2007, 9, 2844–2854. [Google Scholar] [CrossRef]
- Qi, Z.; Chen, L.; Zhang, W. Comparison of transcriptional heterogeneity of eight genes between batch Desulfovibrio vulgaris biofilm and planktonic culture at a single-cell level. Front. Microbiol. 2016, 7, 597. [Google Scholar] [CrossRef]
- Clark, M.E.; He, Z.; Redding, A.M.; Joachimiak, M.P.; Keasling, J.D.; Zhou, J.Z.; Arkin, A.P.; Mukhopadhyay, A.; Fields, M.W. Transcriptomic and proteomic analyses of Desulfovibrio vulgaris biofilms: Carbon and energy flow contribute to the distinct biofilm growth state. BMC Genom. 2012, 13, 138. [Google Scholar] [CrossRef]
- Zhang, W.; Culley, D.E.; Nie, L.; Scholten, J.C. Comparative transcriptome analysis of Desulfovibrio vulgaris grown in planktonic culture and mature biofilm on a steel surface. Appl. Microbiol. Biotechnol. 2007, 76, 447–457. [Google Scholar] [CrossRef]
- Cakar, A. Bacterial communication: Quorum-sensing. Mikrobiyoloji Bul. 2004, 38, 273–284. [Google Scholar]
- Pickering, K.L.; Verbeek, C.J.; Viljoen, C. The effect of aqueous urea on the processing, structure and properties of CGM. J. Polym. Environ. 2012, 20, 335–343. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Larsson, G.; Törnkvist, M. Rapid sampling, cell inactivation and evaluation of low extracellular glucose concentrations during fed-batch cultivation. J. Biotechnol. 1996, 49, 69–82. [Google Scholar] [CrossRef]
- Burton, K. A study of the conditions and mechanism of the diphenylamine reaction for the colorimetric estimation of deoxyribonucleic acid. Biochem. J. 1956, 62, 315. [Google Scholar] [CrossRef]
- ISO 8501-1:2007; Preparation Steel Surface before Application of Paints and Related Products. International Organization for Standardization, ISO Norm: Geneva, Switzerland, 2007.
- CB, W. Extracellular DNA required foe bacterial biofilm formation. Science 2002, 295, 1487. [Google Scholar]
- Vilain, S.; Pretorius, J.M.; Theron, J.; Brözel, V.S. DNA as an adhesin: Bacillus cereus requires extracellular DNA to form biofilms. Appl. Environ. Microbiol. 2009, 75, 2861–2868. [Google Scholar] [CrossRef]
- Hlady, V.; Buijs, J. Protein adsorption on solid surfaces. Curr. Opin. Biotechnol. 1996, 7, 72–77. [Google Scholar] [CrossRef]
- Cardoso Pereira, I.; Haveman, S.; Voordouw, G. Biochemical, genetic and genomic characterization of anaerobic electron transport pathways in sulphate-reducing Delta proteobacteria. In Sulphate-Reducing Bacteria: Environmental and Engineered Systems; Cambridge University Press: Cambridge, UK, 2007; pp. 215–240. [Google Scholar]
- Bencharit, S.; Ward, M.J. Chemotactic responses to metals and anaerobic electron acceptors in Shewanella oneidensis MR-1. J. Bacteriol. 2005, 187, 5049–5053. [Google Scholar] [CrossRef]
- Childers, S.E.; Ciufo, S.; Lovley, D.R. Geobacter metallireducens accesses insoluble Fe(III) oxide by chemotaxis. Nature 2002, 416, 767–769. [Google Scholar] [CrossRef]
- Wadhams, G.H.; Armitage, J.P. Making sense of it all: Bacterial chemotaxis. Nat. Rev. Mol. Cell Biol. 2004, 5, 1024–1037. [Google Scholar] [CrossRef]
- Prabhakaran, P.; Ashraf, M.A.; Aqma, W.S. Microbial stress response to heavy metals in the environment. RSC Adv. 2016, 6, 109862–109877. [Google Scholar] [CrossRef]
- Sourjik, V. Receptor clustering and signal processing in E. coli chemotaxis. Trends Microbiol. 2004, 12, 569–576. [Google Scholar] [CrossRef]
- Biemann, H.P.; Koshland, D.E. Aspartate receptors of escherichia-coli and salmonella-typhimurium bind ligand with negative and half-of-the-sites cooperativity. Biochemistry 1994, 33, 629–634. [Google Scholar] [CrossRef]
- Li, E.; Wu, J.; Zhang, D.; Wang, P.; Zhu, L.; Li, C.; Sun, Z.; Gao, Y. Effect of autoinducer-2 on corrosion of Q235 carbon steel caused by sulfate reducing bacteria. Corros. Sci. 2022, 200, 110220. [Google Scholar] [CrossRef]
- Berg, H.C. The rotary motor of bacterial flagella. Annu. Rev. Biochem. 2003, 72, 19–54. [Google Scholar] [CrossRef]
- Hengge, R. Principles of c-di-GMP signalling in bacteria. Nat. Rev. Microbiol. 2009, 7, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Jenal, U.; Malone, J. Mechanisms of cyclic-di-GMP signaling in bacteria. Annu. Rev. Genet. 2006, 40, 385–407. [Google Scholar] [CrossRef] [PubMed]
- Schirmer, T.; Jenal, U. Structural and mechanistic determinants of c-di-GMP signalling. Nat. Rev. Microbiol. 2009, 7, 724–735. [Google Scholar] [CrossRef] [PubMed]
- Xuan, T.-F.; Wang, Z.-Q.; Liu, J.; Yu, H.-T.; Lin, Q.-W.; Chen, W.-M.; Lin, J. Design and synthesis of novel c-di-GMP G-quadruplex inducers as bacterial biofilm inhibitors. J. Med. Chem. 2021, 64, 11074–11089. [Google Scholar] [CrossRef] [PubMed]
- Hee, C.-S.; Habazettl, J.; Schmutz, C.; Schirmer, T.; Jenal, U.; Grzesiek, S. Intercepting second-messenger signaling by rationally designed peptides sequestering c-di-GMP. Proc. Natl. Acad. Sci. USA 2020, 117, 17211–17220. [Google Scholar] [CrossRef]
- Furukawa, K.; Gu, H.; Sudarsan, N.; Hayakawa, Y.; Hyodo, M.; Breaker, R.R. Identification of ligand analogues that control c-di-GMP riboswitches. ACS Chem. Biol. 2012, 7, 1436–1443. [Google Scholar] [CrossRef]
- Huang, T.; Peng, Z.; Hu, M.; Xiao, Y.-S.; Liu, Z.-G.; Guan, Q.-Q.; Xie, M.-Y.; Tao, X. Interactions between Lactobacillus plantarum NCU116 and its environments based on extracellular proteins and polysaccharides prediction by comparative analysis. Genomics 2020, 112, 3579–3587. [Google Scholar] [CrossRef]
- Flower, A.M. The SecY translocation complex: Convergence of genetics and structure. TRENDS Microbiol. 2007, 15, 203–210. [Google Scholar] [CrossRef]
- Tsirigotaki, A.; De Geyter, J.; Economou, A.; Karamanou, S. Protein export through the bacterial Sec pathway. Nat. Rev. Microbiol. 2017, 15, 21–36. [Google Scholar] [CrossRef]
- Beck, B.J.; Downs, D.M. The apbE gene encodes a lipoprotein involved in thiamine synthesis in Salmonella typhimurium. J. Bacteriol. 1998, 180, 885–891. [Google Scholar] [CrossRef]
- Beck, B.J.; Downs, D.M. A periplasmic location is essential for the role of the ApbE lipoprotein in thiamine synthesis in Salmonella typhimurium. J. Bacteriol. 1999, 181, 7285–7290. [Google Scholar] [CrossRef] [PubMed]
- Skovran, E.; Downs, D.M. Lack of the ApbC or ApbE protein results in a defect in Fe-S cluster metabolism in Salmonella enterica serovar Typhimurium. J. Bacteriol. 2003, 185, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Dunsmore, B.; Whitfield, T.; Lawson, P.; Collins, M. Corrosion by sulfate reducing bacteria that utilize nitrate. Presented at the CORROSION 2004, New Orleans, LA, USA, 28 March–1 April 2004. [Google Scholar]
- Flemming, H.-C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Thauer, R.K.; Stackebrandt, E.; Hamilton, W.A. Energy metabolism and phylogenetic diversity of sulphate-reducing bacteria. In Sulphate-Reducing Bacteria: Environmental and Engineered Systems; Cambridge University Press: Cambridge, UK, 2007; pp. 1–37. [Google Scholar]
- Dawson, M.P.; Humphrey, B.A.; Marshall, K.C. Adhesion: A tactic in the survival strategy of a marine vibrio during starvation. Curr. Microbiol. 1981, 6, 195–199. [Google Scholar] [CrossRef]
- Kjelleberg, S.; Humphrey, B.A.; Marshall, K.C. Effect of interfaces on small, starved marine bacteria. Appl. Environ. Microbiol. 1982, 43, 1166–1172. [Google Scholar] [CrossRef]
- Saini, G.; Nasholm, N.; Wood, B.D. Implications of growth and starvation conditions in bacterial adhesion and transport. J. Adhes. Sci. Technol. 2011, 25, 2281–2297. [Google Scholar] [CrossRef]
- Kjelleberg, S.; Humphrey, B.A.; Marshall, K.C. Initial phases of starvation and activity of bacteria at surfaces. Appl. Environ. Microbiol. 1983, 46, 978–984. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, I.W. Biofilm exopolysaccharides: A strong and sticky framework. Microbiology 2001, 147, 3–9. [Google Scholar] [CrossRef]
- Odom, J.; Peck, H., Jr. Hydrogen cycling as a general mechanism for energy coupling in the sulfate-reducing bacteria, Desulfovibrio sp. FEMS Microbiol. Lett. 1981, 12, 47–50. [Google Scholar] [CrossRef]
- Voordouw, G. Carbon monoxide cycling by Desulfovibrio vulgaris Hildenborough. J. Bacteriol. 2002, 184, 5903–5911. [Google Scholar] [CrossRef]
- Heidelberg, J.F.; Seshadri, R.; Haveman, S.A.; Hemme, C.L.; Paulsen, I.T.; Kolonay, J.F.; Eisen, J.A.; Ward, N.; Methe, B.; Brinkac, L.M. The genome sequence of the anaerobic, sulfate-reducing bacterium Desulfovibrio vulgaris Hildenborough. Nat. Biotechnol. 2004, 22, 554–559. [Google Scholar] [CrossRef]
- Müller, V.; Imkamp, F.; Biegel, E.; Schmidt, S.; Dilling, S. Discovery of a ferredoxin: NAD+-oxidoreductase (Rnf) in Acetobacterium woodii: A novel potential coupling site in acetogens. Ann. N. Y. Acad. Sci. 2008, 1125, 137–146. [Google Scholar] [CrossRef]
- Schmehl, M.; Jahn, A.; Meyer zu Vilsendorf, A.; Hennecke, S.; Masepohl, B.; Schuppler, M.; Marxer, M.; Oelze, J.; Klipp, W. Identification of a new class of nitrogen fixation genes in Rhodobacter capsalatus: A putative membrane complex involved in electron transport to nitrogenase. Mol. Gen. Genet. MGG 1993, 241, 602–615. [Google Scholar] [CrossRef]
- Herrmann, G.; Jayamani, E.; Mai, G.; Buckel, W. Energy conservation via electron-transferring flavoprotein in anaerobic bacteria. J. Bacteriol. 2008, 190, 784–791. [Google Scholar] [CrossRef]
- Thauer, R.K.; Jungermann, K.; Decker, K. Energy conservation in chemotrophic anaerobic bacteria. Bacteriol. Rev. 1977, 41, 100–180. [Google Scholar] [CrossRef]
- Jungermann, K.; Kirchniawy, H.; Thauer, R. Ferredoxin dependent CO2 reduction to formate in clostridium pasteurianum. Biochem. Biophys. Res. Commun. 1970, 41, 682–689. [Google Scholar] [CrossRef]
- Ragsdale, S.W.; Ljungdahl, L.G. Hydrogenase from Acetobacterium woodii. Arch. Microbiol. 1984, 139, 361–365. [Google Scholar] [CrossRef]
- Shanmugasundaram, T.; Wood, H. Interaction of ferredoxin with carbon monoxide dehydrogenase from Clostridium thermoaceticum. J. Biol. Chem. 1992, 267, 897–900. [Google Scholar] [CrossRef]
- Li, F.; Hinderberger, J.; Seedorf, H.; Zhang, J.; Buckel, W.; Thauer, R.K. Coupled ferredoxin and crotonyl coenzyme A (CoA) reduction with NADH catalyzed by the butyryl-CoA dehydrogenase/Etf complex from Clostridium kluyveri. J. Bacteriol. 2008, 190, 843–850. [Google Scholar] [CrossRef]
- Dörries, M.; Wöhlbrand, L.; Rabus, R. Differential proteomic analysis of the metabolic network of the marine sulfate-reducer Desulfobacterium autotrophicum HRM2. Proteomics 2016, 16, 2878–2893. [Google Scholar] [CrossRef]
- Pereira, I.A.C.; Ramos, A.R.; Grein, F.; Marques, M.C.; da Silva, S.M.; Venceslau, S.S. A comparative genomic analysis of energy metabolism in sulfate reducing bacteria and archaea. Front. Microbiol. 2011, 2, 69. [Google Scholar] [CrossRef]
- Marietou, A.; Kjeldsen, K.U.; Glombitza, C.; Jørgensen, B.B. Response to substrate limitation by a marine sulfate-reducing bacterium. ISME J. 2022, 16, 200–210. [Google Scholar] [CrossRef]
- Meyer, B.; Kuehl, J.V.; Price, M.N.; Ray, J.; Deutschbauer, A.M.; Arkin, A.P.; Stahl, D.A. The energy-conserving electron transfer system used by Desulfovibrio alaskensis strain G 20 during pyruvate fermentation involves reduction of endogenously formed fumarate and cytoplasmic and membrane-bound complexes, Hdr-Flox and Rnf. Environ. Microbiol. 2014, 16, 3463–3486. [Google Scholar] [CrossRef]
- Rabus, R.; Hansen, T.A.; Widdel, F. Dissimilatory sulfate-and sulfur-reducing prokaryotes. Prokaryotes 2006, 2, 659–768. [Google Scholar]
- Adav, S.S.; Lee, D.-J.; Tay, J.-H. Extracellular polymeric substances and structural stability of aerobic granule. Water Res. 2008, 42, 1644–1650. [Google Scholar] [CrossRef]
- Wang, X.; An, Q.; Zhao, B.; Guo, J.S.; Huang, Y.S.; Tian, M. Auto-aggregation properties of a novel aerobic denitrifier Enterobacter sp. strain FL. Appl. Microbiol. Biotechnol. 2018, 102, 2019–2030. [Google Scholar] [CrossRef]
- Ding, X.S.; Zhao, B.; An, Q.; Tian, M.; Guo, J.S. Role of extracellular polymeric substances in biofilm formation by Pseudomonas stutzeri strain XL-2. Appl. Microbiol. Biotechnol. 2019, 103, 9169–9180. [Google Scholar] [CrossRef]
- Wu, B.; Ni, B.-J.; Horvat, K.; Song, L.; Chai, X.; Dai, X.; Mahajan, D. Occurrence state and molecular structure analysis of extracellular proteins with implications on the dewaterability of waste-activated sludge. Environ. Sci. Technol. 2017, 51, 9235–9243. [Google Scholar] [CrossRef]
- Guo, Z.; Ma, L.; Dai, Q.; Liu, Y.; Zhang, D.; Ao, R. Behavior of heavy metals in municipal sludge during dewatering: The role of conditioners and extracellular polymeric substances. J. Environ. Manag. 2023, 342, 118287. [Google Scholar] [CrossRef]
- Dong, Z.H.; Liu, T.; Liu, H.F. Influence of EPS isolated from thermophilic sulphate-reducing bacteria on carbon steel corrosion. Biofouling 2011, 27, 487–495. [Google Scholar] [CrossRef]
- Gillette, M.A.; Satpathy, S.; Cao, S.; Dhanasekaran, S.M.; Vasaikar, S.V.; Krug, K.; Petralia, F.; Li, Y.; Liang, W.-W.; Reva, B.; et al. Proteogenomic characterization reveals therapeutic vulnerabilities in lung adenocarcinoma. Cell 2020, 182, 200–225.e35. [Google Scholar] [CrossRef] [PubMed]
- Kachuk, C.; Stephen, K.; Doucette, A. Comparison of sodium dodecyl sulfate depletion techniques for proteome analysis by mass spectrometry. J. Chromatogr. A 2015, 1418, 158–166. [Google Scholar] [CrossRef]
- Marx, H.; Minogue, C.E.; Jayaraman, D.; Richards, A.L.; Kwiecien, N.W.; Siahpirani, A.F.; Rajasekar, S.; Maeda, J.; Garcia, K.; Del Valle-Echevarria, A.R.; et al. A proteomic atlas of the legume Medicago truncatula and its nitrogen-fixing endosymbiont Sinorhizobium meliloti. Nat. Biotechnol. 2016, 34, 1198–1205. [Google Scholar] [CrossRef]
- Niu, L.; Zhang, H.; Wu, Z.; Wang, Y.; Liu, H.; Wu, X.; Wang, W. Modified TCA/acetone precipitation of plant proteins for proteomic analysis. PLoS ONE 2018, 13, e0202238. [Google Scholar] [CrossRef] [PubMed]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Xiong, E.; Wang, W.; Scali, M.; Cresti, M. Universal sample preparation method integrating trichloroacetic acid/acetone precipitation with phenol extraction for crop proteomic analysis. Nat. Protoc. 2014, 9, 362–374. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, T.; Zhang, Z.; Payne, S.H.; Zhang, B.; McDermott, J.E.; Zhou, J.-Y.; Petyuk, V.A.; Chen, L.; Ray, D.; et al. Integrated proteogenomic characterization of human high-grade serous ovarian cancer. Cell 2016, 166, 755–765. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cells | Element | Binding Energy (eV) | Relatives Content | Proposed Components |
|---|---|---|---|---|
| Plankton | C 1s | 284.4 | 23.50% | Aromatic carbon |
| 284.8 | 14.30% | C-(C,H) | ||
| 285.1 | 25.30% | C-N | ||
| 286.1 | 23.90% | C-(O,N) | ||
| 288.1 | 13% | N-C=O | ||
| O 1s | 531.3 | 50.70% | C=O/O-C=O | |
| 532.3 | 49.30% | C-O-H/C-O-C | ||
| 533 | 0% | OH-C=O | ||
| Biofilm | C 1s | 284.4 | 23% | Aromatic carbon |
| 284.8 | 13.10% | C-(C,H) | ||
| 285.1 | 21.80% | C-N | ||
| 286.1 | 25.10% | C-(O,N) | ||
| 288.1 | 17.10% | N-C=O | ||
| O 1s | 531.3 | 71.90% | C=O/O-C=O | |
| 532.3 | 11.90% | C-O-H/C-O-C | ||
| 533 | 12.90% | OH-C=O | ||
| 535.8 | 3.30% | H2O |
| Secondary Structures | Wavenumber (cm−1) | Planktonic | Biofilm |
|---|---|---|---|
| Unknown | 1605–1600 | 0 | 2.3 |
| Aggregated strands | 1625–1610 | 8.3 | 6.6 |
| β-sheet extend | 1640–1630 | 24.6 | 0 |
| β-sheet | 1645–1640 | 0 | 29.5 |
| α-helix | 1657–1648 | 26.6 | 18.3 |
| 3-Turn helix | 1676–1659 | 26 | 35.6 |
| Antiparallel β-sheet/aggregated strands | 1695–1680 | 14.5 | 7.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhang, R.; Mathivanan, K.; Zhang, Y.; Yang, L.; Guan, F.; Duan, J. Proteomics and EPS Compositional Analysis Reveals Desulfovibrio bisertensis SY-1 Induced Corrosion on Q235 Steel by Biofilm Formation. Materials 2024, 17, 5060. https://doi.org/10.3390/ma17205060
Wang Y, Zhang R, Mathivanan K, Zhang Y, Yang L, Guan F, Duan J. Proteomics and EPS Compositional Analysis Reveals Desulfovibrio bisertensis SY-1 Induced Corrosion on Q235 Steel by Biofilm Formation. Materials. 2024; 17(20):5060. https://doi.org/10.3390/ma17205060
Chicago/Turabian StyleWang, Yanan, Ruiyong Zhang, Krishnamurthy Mathivanan, Yimeng Zhang, Luhua Yang, Fang Guan, and Jizhou Duan. 2024. "Proteomics and EPS Compositional Analysis Reveals Desulfovibrio bisertensis SY-1 Induced Corrosion on Q235 Steel by Biofilm Formation" Materials 17, no. 20: 5060. https://doi.org/10.3390/ma17205060
APA StyleWang, Y., Zhang, R., Mathivanan, K., Zhang, Y., Yang, L., Guan, F., & Duan, J. (2024). Proteomics and EPS Compositional Analysis Reveals Desulfovibrio bisertensis SY-1 Induced Corrosion on Q235 Steel by Biofilm Formation. Materials, 17(20), 5060. https://doi.org/10.3390/ma17205060