
Mechanical Characterization at the Microscale of Mineralized Bone Callus after Bone Lengthening



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
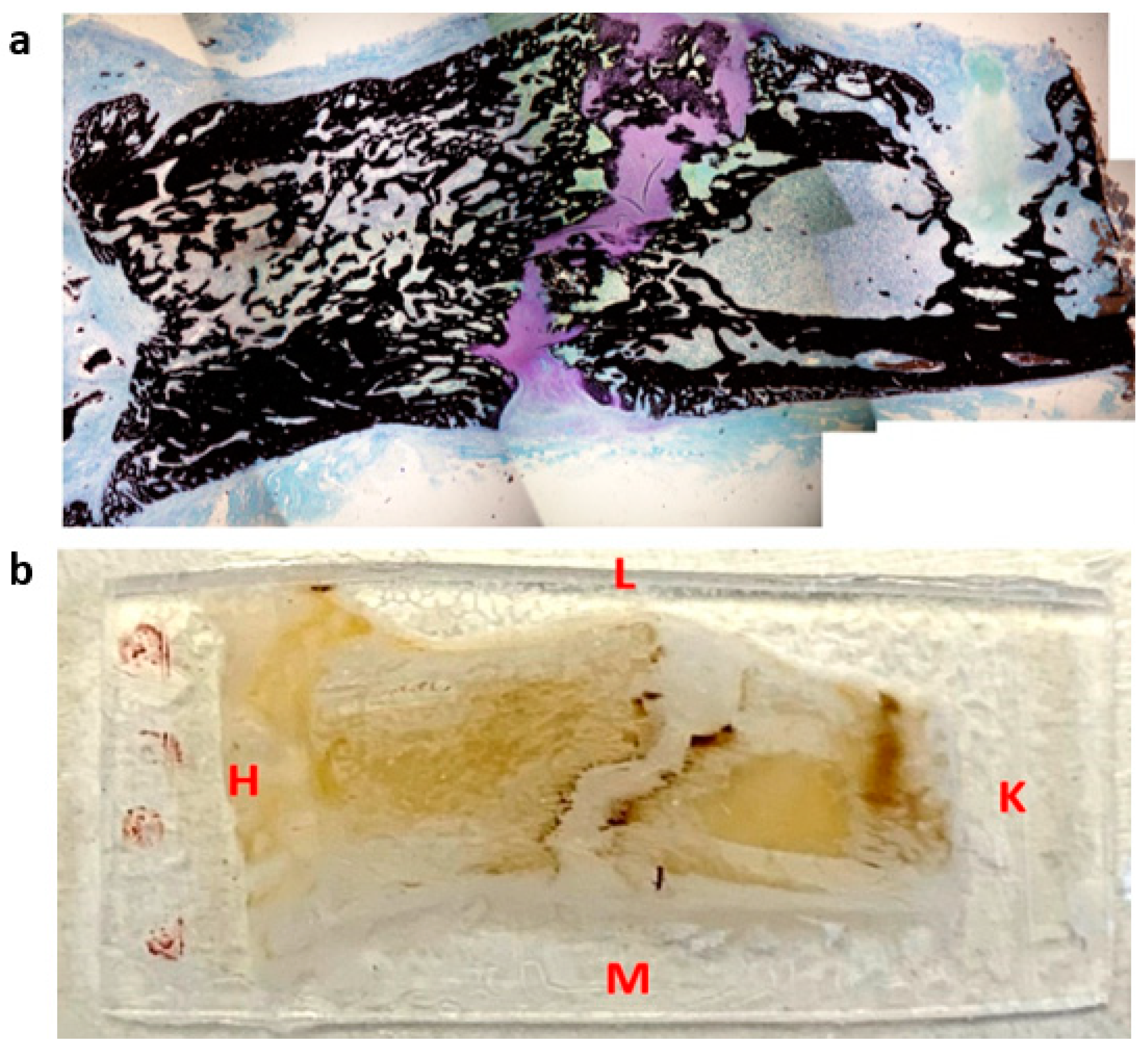
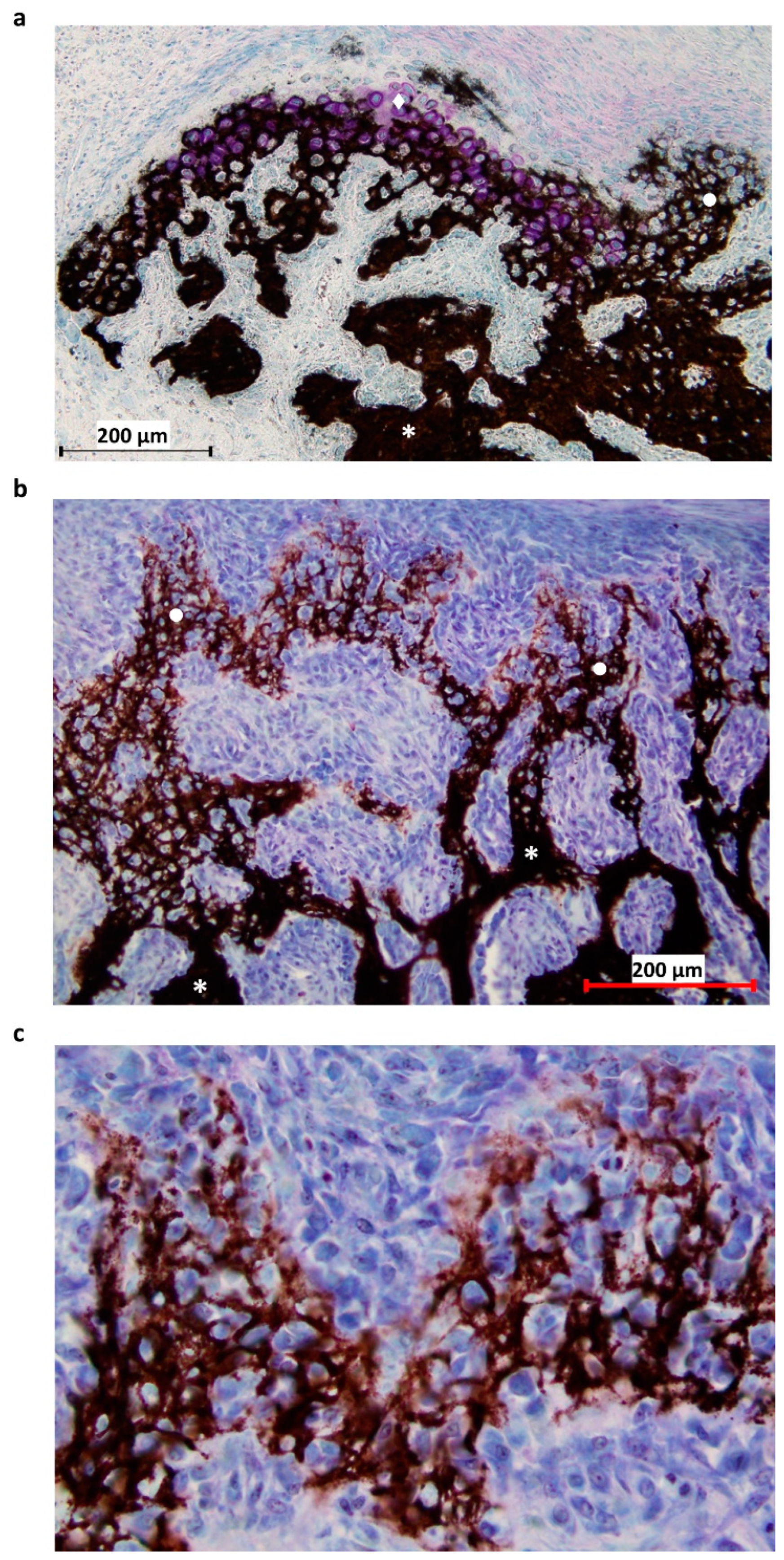
2.2. Histological Staining
2.3. Microindentation
2.3.1. Regions of Interest (ROI)
2.3.2. Tissue Type
2.4. Data Processing and Statistical Analysis
3. Results
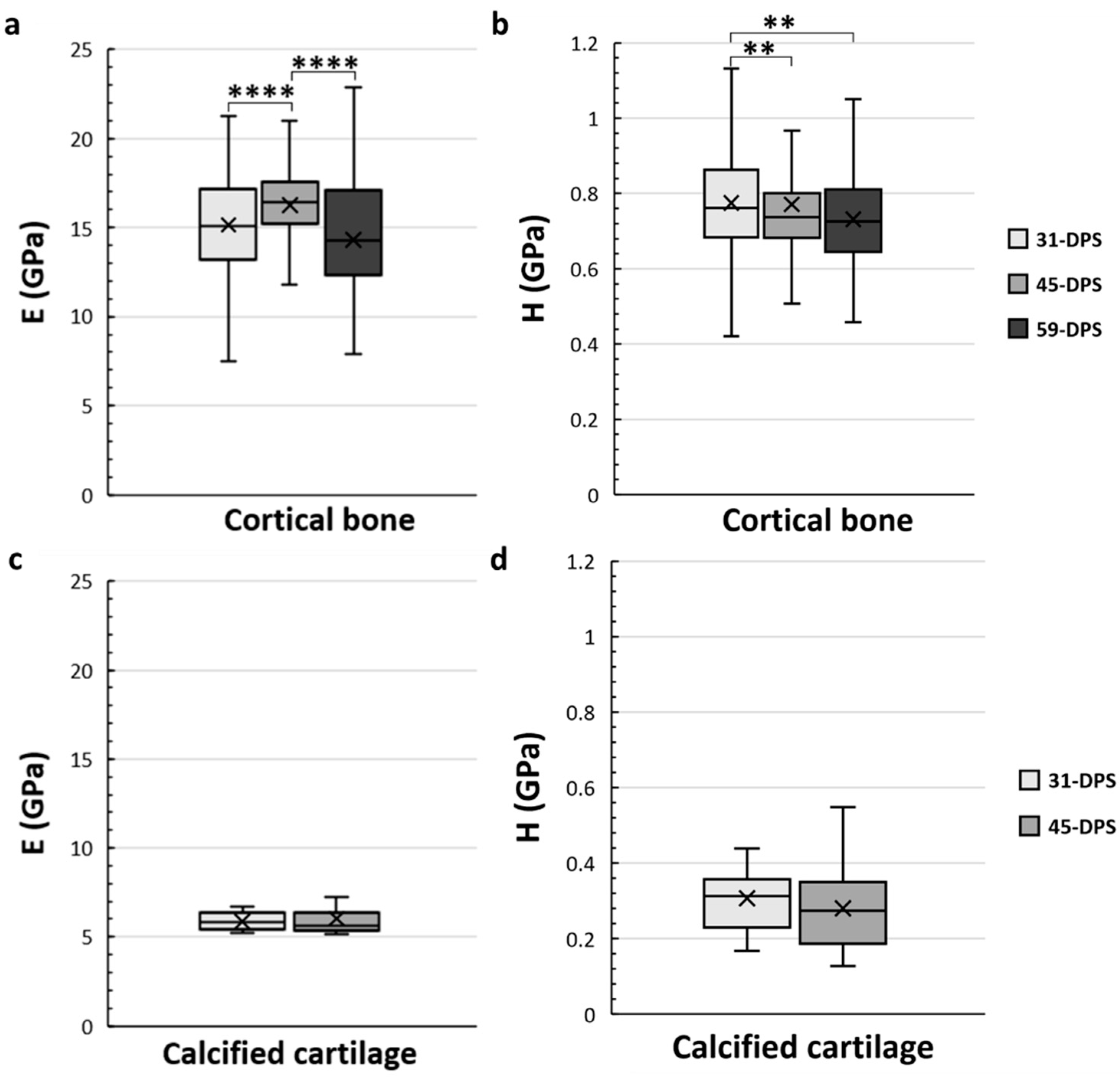
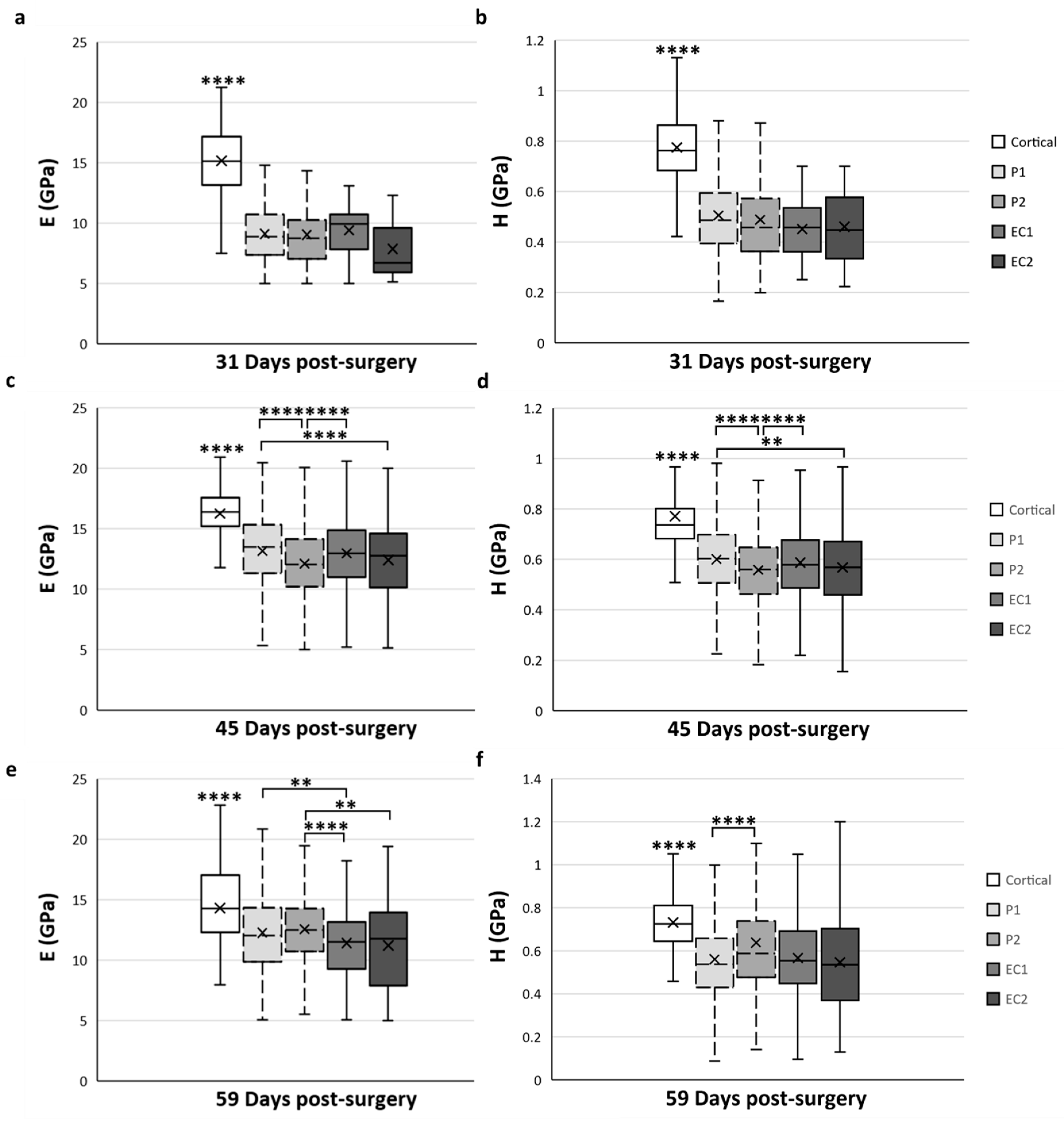
3.1. Temporal Variation of Mineralized Tissue within the Distracted Callus
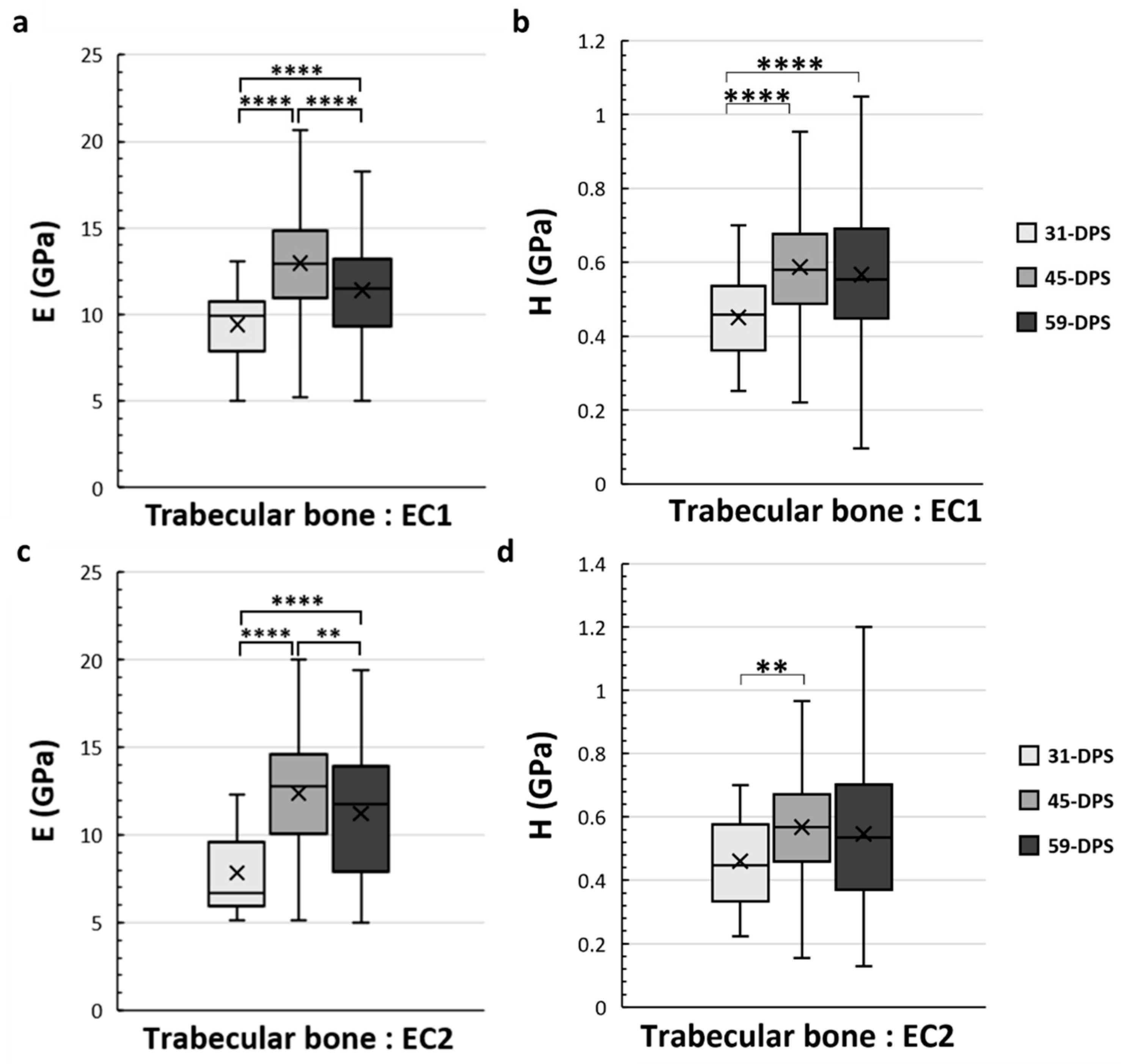
3.2. Spatial Variation of Mineralized Tissue within the Distracted Callus
4. Discussion
4.1. Temporal Variation of the Mineralized Tissues
4.2. Spatial Variation of the Mineralized Tissues
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Jauregui, J.J.; Ventimiglia, A.V.; Grieco, P.W.; Frumberg, D.B.; Herzenberg, J.E. Regenerate Bone Stimulation Following Limb Lengthening: A Meta-Analysis. BMC Musculoskelet. Disord. 2016, 17, 407. [Google Scholar] [CrossRef]
- AI-Aql, Z.S.; Alagl, A.S.; Graves, D.T.; Gerstenfeld, L.C.; Einhorn, T.A. Molecular Mechanisms Controlling Bone Formation during Fracture Healing and Distraction Osteogenesis. J. Dent. Res. 2008, 87, 107–118. [Google Scholar] [CrossRef]
- Singh, M.; Vashistha, A.; Chaudhary, M.; Kaur, G. Biological Basis of Distraction Osteogenesis—A Review. J. Oral Maxillofac. Surg. Med. Pathol. 2016, 28, 1–7. [Google Scholar] [CrossRef]
- Amir, L.R.; Everts, V.; Bronckers, A.L.J.J. Bone Regeneration during Distraction Osteogenesis. Odontology 2009, 97, 63–75. [Google Scholar] [CrossRef]
- Choi, I.H.; Chung, C.Y.; Cho, T.J.; Yoo, W.J. Angiogenesis and Mineralization During Distraction Osteogenesis. J. Korean Med. Sci. 2002, 17, 435. [Google Scholar] [CrossRef] [PubMed]
- Sailhan, F. Bone Lengthening (Distraction Osteogenesis): A Literature Review. Osteoporos Int. 2011, 22, 2011–2015. [Google Scholar] [CrossRef]
- Dhaliwal, K.; Kunchur, R.; Farhadieh, R. Review of the Cellular and Biological Principles of Distraction Osteogenesis: An in Vivo Bioreactor Tissue Engineering Model. J. Plast. Reconstr. Aesthet. Surg. 2016, 69, e19–e26. [Google Scholar] [CrossRef] [PubMed]
- Bouletreau, P.J.; Warren, S.M.; Longaker, M.T. The Molecular Biology of Distraction Osteogenesis. J. Cranio-Maxillofac. Surg. 2002, 30, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Welch, R.D.; Lewis, D.D. Distraction Osteogenesis. Vet. Clin. North Am. Small Anim. Pract. 1999, 29, 1187–1205. [Google Scholar] [CrossRef]
- Li, R.; Saleh, M.; Yang, L.; Coulton, L. Radiographic Classification of Osteogenesis during Bone Distraction. J. Orthop. Res. 2006, 24, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Alam, N.; St-Arnaud, R.; As, D.L.; Rosen, V.; Hamdy, R.C. Are Endogenous BMPs Necessary for Bone Healing during Distraction Osteogenesis? Clin. Orthop. Relat. Res. 2009, 467, 3190–3198. [Google Scholar] [CrossRef]
- Aronson, J.; Harrison, B.H.; Stewart, C.L.; Harp, J.H. The Histology of Distraction Osteogenesis Using Different External Fixators. Clin. Orthop. Relat. Res. 1989, 241, 106–116. [Google Scholar] [CrossRef]
- Ekeland, A.; Engesæter, L.B.; Langeland, N. Mechanical Properties of Fractured and Intact Rat Femora Evaluated by Bending, Torsional and Tensile Tests. Acta Orthop. Scand. 1981, 52, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Abbaspour, A.; Takahashi, M.; Sairyo, K.; Takata, S.; Yukata, K.; Inui, A.; Yasui, N. Optimal Increase in Bone Mass by Continuous Local Infusion of Alendronate during Distraction Osteogenesis in Rabbits. Bone 2009, 44, 917–923. [Google Scholar] [CrossRef] [PubMed]
- Aleksyniene, R.; Thomsen, J.S.; Eckardt, H.; Bundgaard, K.G.; Lind, M.; Hvid, I. Parathyroid Hormone PTH(1–34) Increases the Volume, Mineral Content, and Mechanical Properties of Regenerated Mineralizing Tissue after Distraction Osteogenesis in Rabbits. Acta Orthop. 2009, 80, 716–723. [Google Scholar] [CrossRef]
- Leong, P.L.; Morgan, E.F. Measurement of Fracture Callus Material Properties via Nanoindentation. Acta Biomater. 2008, 4, 1569–1575. [Google Scholar] [CrossRef] [PubMed]
- Manjubala, I.; Liu, Y.; Epari, D.R.; Roschger, P.; Schell, H.; Fratzl, P.; Duda, G.N. Spatial and Temporal Variations of Mechanical Properties and Mineral Content of the External Callus during Bone Healing. Bone 2009, 45, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Mora-Macías, J.; Pajares, A.; Miranda, P.; Domínguez, J.; Reina-Romo, E. Mechanical Characterization via Nanoindentation of the Woven Bone Developed during Bone Transport. J. Mech. Behav. Biomed. Mater. 2017, 74, 236–244. [Google Scholar] [CrossRef]
- Boivin, G.; Farlay, D.; Bala, Y.; Doublier, A.; Meunier, P.J.; Delmas, P.D. Influence of Remodeling on the Mineralization of Bone Tissue. Osteoporos Int. 2009, 20, 1023–1026. [Google Scholar] [CrossRef]
- Boivin, G.; Bala, Y.; Doublier, A.; Farlay, D.; Ste-Marie, L.G.; Meunier, P.J.; Delmas, P.D. The Role of Mineralization and Organic Matrix in the Microhardness of Bone Tissue from Controls and Osteoporotic Patients. Bone 2008, 43, 532–538. [Google Scholar] [CrossRef]
- Pithioux, M.; Roseren, F.; Jalain, C.; Launay, F.; Charpiot, P.; Chabrand, P.; Roffino, S.; Lamy, E. An Efficient and Reproducible Protocol for Distraction Osteogenesis in a Rat Model Leading to a Functional Regenerated Femur. JoVE 2017, 128, e56433. [Google Scholar] [CrossRef] [PubMed]
- Klapperich, C.; Komvopoulos, K.; Pruitt, L. Nanomechanical Properties of Polymers Determined From Nanoindentation Experiments. J. Tribol. 2001, 123, 624–631. [Google Scholar] [CrossRef]
- Donnelly, E.; Baker, S.P.; Boskey, A.L. Effects of Surface Roughness and Maximum Load on the Mechanical Properties of Cancellous Bone Measured by Nanoindentation. J. Biomed. Mater. Res. A 2006, 77, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Oliver, W.C.; Pharr, G.M. An Improved Technique for Determining Hardness and Elastic Modulus Using Load and Displacement Sensing Indentation Experiments. J. Mater. Res. 1992, 7, 1564–1583. [Google Scholar] [CrossRef]
- Kan, Q.; Yan, W.; Kang, G.; Sun, Q. Oliver–Pharr Indentation Method in Determining Elastic Moduli of Shape Memory Alloys—A Phase Transformable Material. J. Mech. Phys. Solids 2013, 61, 2015–2033. [Google Scholar] [CrossRef]
- Turner, C.H.; Burr, D.B. Experimental Techniques for Bone Mechanics. In Bone Mechanics Handbook, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Djasim, U.M.; Mathot, B.J.; Wolvius, E.B.; van Neck, J.W.; van der Wal, K.G.H. Histomorphometric Comparison between Continuous and Discontinuous Distraction Osteogenesis. J. Cranio-Maxillofac. Surg. 2009, 37, 398–404. [Google Scholar] [CrossRef]
- Hvid, I.; Horn, J.; Huhnstock, S.; Steen, H. The Biology of Bone Lengthening. J. Child. Orthop. 2016, 10, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Runyan, C.M.; Gabrick, K.S. Biology of Bone Formation, Fracture Healing, and Distraction Osteogenesis. J. Craniofac. Surg. 2017, 28, 1380–1389. [Google Scholar] [CrossRef]
- Li, G.; Dickson, G.R.; Marsh, D.R.; Simpson, H. Rapid New Bone Tissue Remodeling during Distraction Osteogenesis Is Associated with Apoptosis. J. Orthop. Res. 2003, 21, 28–35. [Google Scholar] [CrossRef]
- Pickering, V.; Jordan, R.C.; Schmidt, B.L.; Looser, K.G.; Shah, J.P.; Strong, E.W.; Chen, T.Y.; Emrich, L.J.; Driscoll, D.L.; Slootweg, P.J.; et al. Principles of Distraction Osteogenesis. Curr. Ther. Oral Maxillofac. Surg. 2011, 38, 101. [Google Scholar]
- Forriol, F.; Denaro, L.; Longo, U.G.; Taira, H.; Maffulli, N.; Denaro, V. Bone Lengthening Osteogenesis, a Combination of Intramembranous and Endochondral Ossification: An Experimental Study in Sheep. Strateg. Trauma Limb. Reconstr. 2010, 5, 71–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Simpson, A.H.R.W.; Triffitt, J.T. The Role of Chondrocytes in Intramembranous and Endochondral Ossification During Distraction Osteogenesis in the Rabbit. Calcif. Tissue Int. 1999, 64, 310–317. [Google Scholar] [CrossRef]
- Fink, B.; Pollnau, C.; Vogel, M.; Skripitz, R.; Enderle, A. Histomorphometry of Distraction Osteogenesis During Experimental Tibial Lengthening. J. Orthop. Trauma 2003, 17, 113–118. [Google Scholar] [CrossRef]
- Yasui, N.; Sato, M.; Ochi, T.; Kimura, T.; Kawahata, H.; Kitamura, Y.; Nomura, S. Three Modes of Ossification during Distraction Osteogenesis in the Rat. Bone Jt. J. 1997, 79, 824–830. [Google Scholar] [CrossRef]
- Doblaré, M.; García, J.M.; Gómez, M.J. Modelling Bone Tissue Fracture and Healing: A Review. Eng. Fract. Mech. 2004, 71, 1809–1840. [Google Scholar] [CrossRef]
- Shapiro, F.; Wu, J.Y. Woven Bone Overview: Structural Classification Based on Its Integral Role in Developmental, Repair and Pathological Bone Formation throughout Vertebrate Groups. Eur. Cell Mater. 2019, 38, 137–167. [Google Scholar] [CrossRef] [PubMed]
- Roohani, I.; Yeo, G.C.; Mithieux, S.M.; Weiss, A.S. Emerging Concepts in Bone Repair and the Premise of Soft Materials. Curr. Opin. Biotechnol. 2022, 74, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Gupta, H.S.; Schratter, S.; Tesch, W.; Roschger, P.; Berzlanovich, A.; Schoeberl, T.; Klaushofer, K.; Fratzl, P. Two Different Correlations between Nanoindentation Modulus and Mineral Content in the Bone–Cartilage Interface. J. Struct. Biol. 2005, 149, 138–148. [Google Scholar] [CrossRef]
- Boskey, A.L. Matrix Proteins and Mineralization: An Overview. Connect. Tissue Res. 1996, 35, 357–363. [Google Scholar] [CrossRef]
- Sun, L.; Fan, Y.; Li, D.; Zhao, F.; Xie, T.; Yang, X.; Gu, Z. Evaluation of the Mechanical Properties of Rat Bone under Simulated Microgravity Using Nanoindentation. Acta Biomater. 2009, 5, 3506–3511. [Google Scholar] [CrossRef]
- Amir, L.R.; Becking, A.G.; Jovanovic, A.; Perdijk, F.B.T.; Everts, V.; Bronckers, A.L.J.J. Vertical Distraction Osteogenesis in the Human Mandible: A Prospective Morphometric Study. Clin. Oral Implant. Res. 2006, 17, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Hargrave-Thomas, E.; van Sloun, F.; Dickinson, M.; Broom, N.; Thambyah, A. Multi-Scalar Mechanical Testing of the Calcified Cartilage and Subchondral Bone Comparing Healthy vs Early Degenerative States. Osteoarthr. Cartil. 2015, 23, 1755–1762. [Google Scholar] [CrossRef]
- Duer, M.J.; Friščić, T.; Murray, R.C.; Reid, D.G.; Wise, E.R. The Mineral Phase of Calcified Cartilage: Its Molecular Structure and Interface with the Organic Matrix. Biophys. J. 2009, 96, 3372–3378. [Google Scholar] [CrossRef] [PubMed]
- Golub, E.E. Biomineralization and Matrix Vesicles in Biology and Pathology. Semin. Immunopathol. 2011, 33, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Touaitahuata, H.; Cres, G.; de Rossi, S.; Vives, V.; Blangy, A. The Mineral Dissolution Function of Osteoclasts Is Dispensable for Hypertrophic Cartilage Degradation during Long Bone Development and Growth. Dev. Biol. 2014, 393, 57–70. [Google Scholar] [CrossRef]
- Boivin, G. Minéralisation et minéralité osseuses. Therapies 2003, 58, 409–413. [Google Scholar] [CrossRef]
- Compton, J.; Fragomen, A.; Rozbruch, S.R. Skeletal Repair in Distraction Osteogenesis: Mechanisms and Enhancements. JBJS Rev. 2015, 3, e2. [Google Scholar] [CrossRef]
- Lacroix, D.; Prendergast, P.J. A Mechano-Regulation Model for Tissue Differentiation during Fracture Healing: Analysis of Gap Size and Loading. J. Biomech. 2002, 35, 1163–1171. [Google Scholar] [CrossRef]
- Isaksson, H.; Comas, O.; van Donkelaar, C.C.; Mediavilla, J.; Wilson, W.; Huiskes, R.; Ito, K. Bone Regeneration during Distraction Osteogenesis: Mechano-Regulation by Shear Strain and Fluid Velocity. J. Biomech. 2007, 40, 2002–2011. [Google Scholar] [CrossRef] [PubMed]
- Claes, L.; Meyers, N.; Schülke, J.; Reitmaier, S.; Klose, S.; Ignatius, A. The Mode of Interfragmentary Movement Affects Bone Formation and Revascularization after Callus Distraction. PLoS ONE 2018, 13, e0202702. [Google Scholar] [CrossRef]
- Hasler, C.C.; Krieg, A.H. Current Concepts of Leg Lengthening. J. Child. Orthop. 2012, 6, 89–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, M.; Kozloff, K.M.; Goulet, J.A.; Goldstein, S.A. Increased Distraction Rates Influence Precursor Tissue Composition without Affecting Bone Regeneration. J. Bone Miner. Res. 2000, 15, 982–989. [Google Scholar] [CrossRef]
- Nowazesh Ali, M.; Ejiri, S.; Kobayashi, T.; Anwar, R.B.; Oda, K.; Ohshima, H.; Saito, C. Histologic Study of the Cellular Events during Rat Mandibular Distraction Osteogenesis. Oral Maxillofac. Surg. 2009, 107, 325–335. [Google Scholar] [CrossRef]
- Kojimoto, H.; Yasui, N.; Goto, T.; Matsuda, S.; Shimomura, Y. Bone Lengthening in Rabbits by Callus Distraction. The Role of Periosteum and Endosteum. J. Bone Jt. Surgery. Br. Vol. 1988, 70-B, 543–549. [Google Scholar] [CrossRef]
- Chang, F.; Mishima, H.; Ishii, T.; Yanai, T.; Akaogi, H.; Sakai, S.; Yoshioka, T.; Ochiai, N. Stimulation of EP4 Receptor Enhanced Bone Consolidation during Distraction Osteogenesis. J. Orthop. Res. 2007, 25, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Loboa, E.G.; Fang, T.D.; Parker, D.W.; Warren, S.M.; Fong, K.D.; Longaker, M.T.; Carter, D.R. Mechanobiology of Mandibular Distraction Osteogenesis: Finite Element Analyses with a Rat Model. J. Orthop. Res. 2005, 23, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Florez, N.; Oyen, M.L.; Shefelbine, S.J. Insight into Differences in Nanoindentation Properties of Bone. J. Mech. Behav. Biomed. Mater. 2013, 18, 90–99. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roseren, F.; Roffino, S.; Pithioux, M. Mechanical Characterization at the Microscale of Mineralized Bone Callus after Bone Lengthening. Materials 2022, 15, 6207. https://doi.org/10.3390/ma15186207
Roseren F, Roffino S, Pithioux M. Mechanical Characterization at the Microscale of Mineralized Bone Callus after Bone Lengthening. Materials. 2022; 15(18):6207. https://doi.org/10.3390/ma15186207
Chicago/Turabian StyleRoseren, Flavy, Sandrine Roffino, and Martine Pithioux. 2022. "Mechanical Characterization at the Microscale of Mineralized Bone Callus after Bone Lengthening" Materials 15, no. 18: 6207. https://doi.org/10.3390/ma15186207
APA StyleRoseren, F., Roffino, S., & Pithioux, M. (2022). Mechanical Characterization at the Microscale of Mineralized Bone Callus after Bone Lengthening. Materials, 15(18), 6207. https://doi.org/10.3390/ma15186207