
Atomic Force Microscopy Investigation of the Interactions between the MCM Helicase and DNA


 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Expression and Purification Protocol of Wild-Type MthMCM and Mutants
2.2. DNA Substrate Preparation
2.3. Sample Preparation
2.4. AFM Imaging and Image Processing
2.5. DNA End-to-End Distance Analysis
3. Results
3.1. DNA on Mica Conformational Analysis
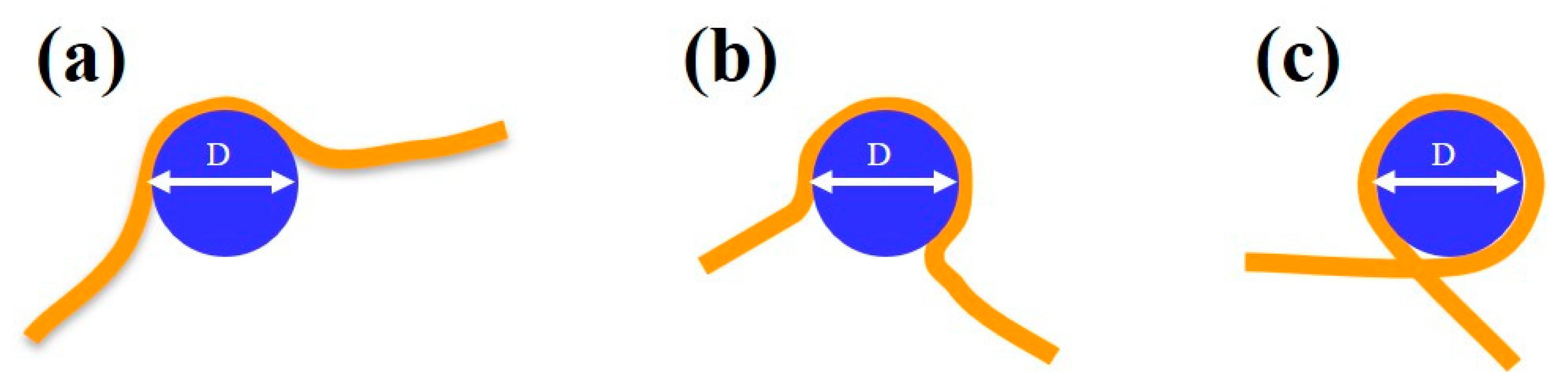
3.2. MCM Protein Complex–DNA Interaction
3.3. MCM Mutants–DNA Interaction
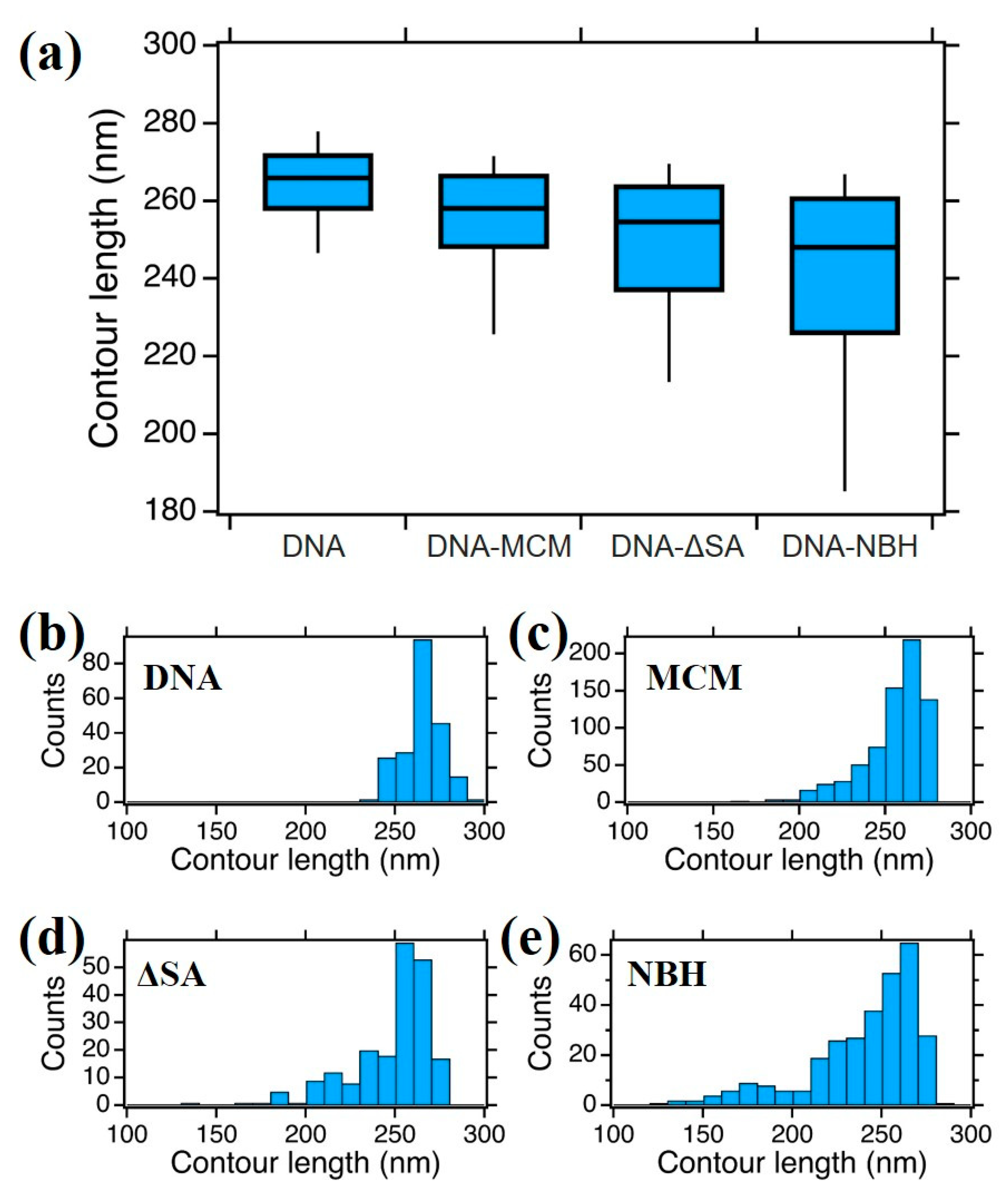
3.4. DNA Contour Length Variation
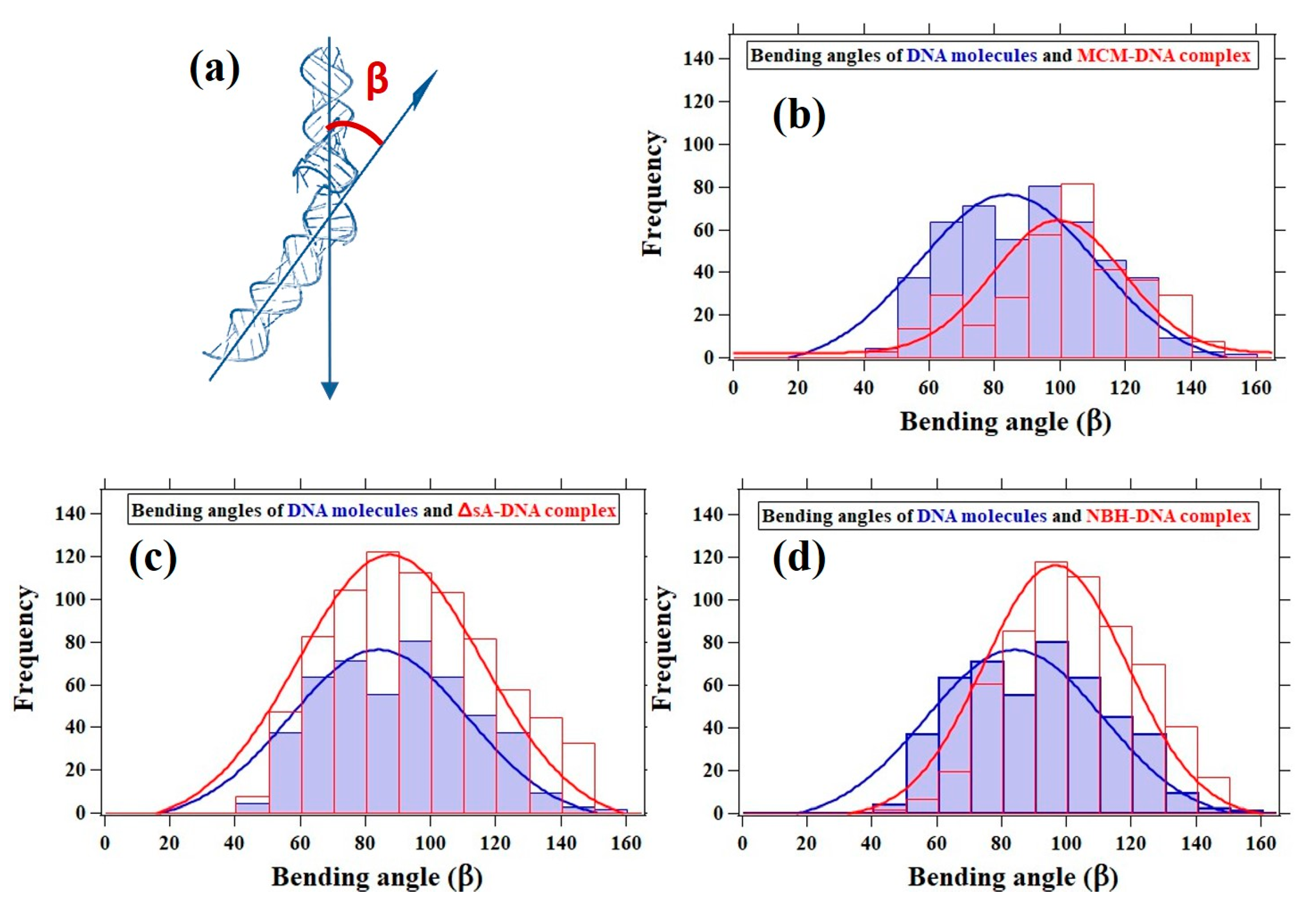
3.5. DNA Bending Angle Analysis
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bustamante, C.; Rivetti, C. Visualizing protein-nucleic acid interactions on a large scale with the scanning force microscopy. Annu. Rev. Biophys. Biomol. Struct. 1996, 25, 395–429. [Google Scholar] [CrossRef] [PubMed]
- Rivetti, C.; Guthold, M.; Bustamante, C. Wrapping of DNA around the E. coli RNA polymerase open promoter complex. EMBO J. 1999, 18, 4464–4475. [Google Scholar] [CrossRef] [PubMed]
- Gaczynska, M.; Osmulski, P.A.; Jiang, Y.; Lee, J.K.; Bermudez, V.; Hurwitz, J. Atomic force microscopic analysis of the binding of the Schizosaccharomyces pombe origin recognition complex and the spOrc4 protein with origin DNA. Proc. Natl. Acad. Sci. USA 2004, 101, 17952–17957. [Google Scholar] [CrossRef] [PubMed]
- Lyubchenko, Y.L.; Gall, A.A.; Shlyakhtenko, L.S. Visualization of DNA and protein-DNA complexes with atomic force microscopy. Methods Mol. Biol. 2014, 1117, 367–384. [Google Scholar] [PubMed]
- Doniselli, N.; Rodriguez-Aliaga, P.; Amidani, D.; Bardales, J.A.; Bustamante, C.; Guerra, D.G.; Rivetti, C. New insights into the regulatory mechanisms of ppGpp and DksA on Escherichia coli RNA polymerase–promoter complex. Nucl. Acids Res. 2015, 43, 5249–5262. [Google Scholar] [CrossRef] [PubMed]
- Beckwitt, E.C.; Kong, M.; Van Houten, B. Studying protein-DNA interactions using atomic force microscopy. Semin. Cell Dev. Biol. 2018, 73, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Maiorano, D.; Lutzmann, M.; Mechali, M. MCM proteins and DNA replication. Curr. Opin. Cell Biol. 2006, 18, 130–136. [Google Scholar] [CrossRef]
- Deegan, T.D.; Diffley, J.F.X. MCM: One ring to rule them all. Curr. Opin. Struct. Biol. 2016, 37, 145–151. [Google Scholar] [CrossRef]
- Abid Ali, F.; Renault, L.; Gannon, J.; Gahlon, H.L.; Kotecha, A.; Zhou, J.C.; Rueda, D.; Costa, A. Cryo-EM structures of the eukaryotic replicative helicase bound to a translocation substrate. Nat. Commun. 2016, 7, 10708. [Google Scholar] [CrossRef]
- Costa, A.; Onesti, S. Structural biology of MCM helicases. Mol. Biol. 2009, 44, 326–342. [Google Scholar] [CrossRef]
- Miller, J.M.; Enemark, E.J. Archaeal MCM proteins as an analog for the eukaryotic Mcm2–7 helicase to reveal essential features of structure and function. Archaea 2015, 2015, 305497. [Google Scholar] [CrossRef] [PubMed]
- Kelman, L.M.; O’Dell, W.B.; Kelman, Z. Unwinding 20 years of the archaeal minichromosome maintenance helicase. J. Bacteriol. 2020, 202, e00729-19. [Google Scholar] [CrossRef] [PubMed]
- Meagher, M.; Epling, L.B.; Enemark, E.J. DNA translocation mechanism of the MCM complex and implications for replication initiation. Nat. Commun. 2019, 10, 3117. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, R.J.; Bishop, B.E.; Leon, R.P.; Sclafani, R.A.; Ogata, C.M.; Chen, X.S. The structure and function of MCM from archaeal M. thermoautotrophicum. Nat. Struct. Biol. 2003, 10, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Onesti, S.; MacNeill, S.A. Structure and evolutionary origins of the CMG complex. Chromosoma 2013, 122, 47–53. [Google Scholar] [CrossRef]
- Medagli, B.; Di Crescenzio, P.; De March, M.; Onesti, S. Structure and activity of the Cdc45-Mcm2-7-GINS (CMG) complex, the replication helicase. In The Initiation of DNA Replication in Eukaryotes; Kaplan, D., Ed.; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Abid Ali, F.; Douglas, M.E.; Locke, J.; Pye, V.E.; Nans, A.; Diffley, J.F.X.; Costa, A. Cryo-EM structure of a licensed DNA replication origin. Nat. Commun. 2017, 8, 2241. [Google Scholar] [CrossRef]
- Kasiviswanathan, R.; Shin, J.H.; Melamud, E.; Kelman, Z. Biochemical characterization of the Methanothermobacter thermautotrophicus minichromosome maintenance (MCM) helicase N-terminal domains. J. Biol. Chem. 2004, 279, 28358–28366. [Google Scholar] [CrossRef]
- Barry, E.R.; McGeoch, A.T.; Kelman, Z.; Bell, S.D. Archaeal MCM has separable processivity, substrate choice and helicase domains. Nucl. Acids Res. 2007, 35, 988–998. [Google Scholar] [CrossRef]
- Pape, T.; Meka, H.; Chen, S.; Vicentini, G.; van Heel, M.; Onesti, S. Hexameric ring structure of the full-length archaeal MCM protein complex. EMBO Rep. 2003, 4, 1079–1083. [Google Scholar] [CrossRef]
- Gomez-Llorente, Y.; Fletcher, R.J.; Chen, X.S.; Carazo, J.M.; San Martin, C. Polymorphism and double hexamer structure in the archaeal minichromosome maintenance (MCM) helicase from Methanobacterium thermoautotrophicum. J. Biol. Chem. 2005, 280, 40909–40915. [Google Scholar] [CrossRef]
- Costa, A.; Pape, T.; van Heel, M.; Brick, P.; Patwardhan, A.; Onesti, S. Structur.al basis of the Methanothermobacter thermautotrophicus MCM helicase activity. Nucleic Acids Res. 2006, 34, 5829–5838. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Pape, T.; van Heel, M.; Brick, P.; Patwardhan, A.; Onesti, S. Structural studies of the archaeal MCM complex in different functional states. J. Struct. Biol. 2006, 156, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Slaymaker, I.M.; Fu, Y.; Toso, D.B.; Ranatunga, N.; Brewster, A.; Forsburg, S.L.; Zhou, Z.H.; Chen, X.S. Mini-chromosome maintenance complexes form a filament to remodel DNA structure and topology. Nucleic Acids Res. 2013, 41, 3446–3456. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhai, Y.; Zhang, Y.; Li, W.; Yang, M.; Lei, J.; Tye, B.; Gao, N. Structure of the eukaryotic MCM complex at 3.8 Å. Nature 2015, 524, 186–191. [Google Scholar] [CrossRef]
- Medagli, B.; Onesti, S. Structure and mechanism of hexameric helicases. Adv. Exp. Med. Biol. 2013, 767, 75–95. [Google Scholar]
- Costa, A.; van Duinen, G.; Medagli, B.; Chong, J.; Sakakibara, N.; Kelman, Z.; Nair, S.K.; Patwardhan, A.; Onesti, S. Cryo-electron microscopy reveals a novel DNA-binding site on the MCM helicase. EMBO J. 2008, 27, 2250–2258. [Google Scholar] [CrossRef]
- Trakselis, M.A.; Seidman, M.M.; Brosh, R.M., Jr. Mechanistic insights into how CMG helicase facilitates replication past DNA roadblocks. DNA Repair. 2017, 55, 76–82. [Google Scholar] [CrossRef]
- Horcas, I.; Fernández, R.; Gómez-Rodríguez, J.M.; Colchero, J.; Gómez-Herrero, J.; Baro, A.M. WSXM: A software for scanning probe microscopy and a tool for nanotechnology. Rev. Sci. Instrum. 2007, 78, 013705. [Google Scholar] [CrossRef]
- Roiter, Y.; Minko, S. AFM single molecule experiments at the solid-liquid interface: In situ conformation of adsorbed flexible polyelectrolyte chains. J. Am. Chem. Soc. 2005, 127, 15688–15689. [Google Scholar] [CrossRef]
- Rivetti, C.; Guthold, M.; Bustamante, C. Scanning force microscopy of DNA deposited onto mica: Equilibration versus Kinetic trapping studied by statistical polymer chain analysis. J. Mol. Biol. 1996, 264, 919–932. [Google Scholar] [CrossRef]
- Rivetti, C.; Walker, C.; Bustamante, C. Polymer chain statistics and conformational analysis of DNA molecules with bends or sections of different flexibility. J. Mol. Biol. 1998, 280, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, C.; Marko, J.F.; Siggia, E.D.; Smith, S.B. Entropic elasticity of λ-phage DNA. Science 1994, 265, 1599–1600. [Google Scholar] [CrossRef] [PubMed]
- Bockelmann, U.; Essevaz-Roulet, B.; Heslot, F. DNA strand separation studied by single molecule force measurements. Phys. Rev. E 1998, 58, 2386–2394. [Google Scholar] [CrossRef]
- Lu, Y.; Weers, B.; Stellwagen, N.C. DNA persistence length revisited. Biopolymers 2002, 61, 261–275. [Google Scholar] [CrossRef]
- Graham, B.W.; Schauer, G.D.; Leuba, S.H.; Trakselis, M.A. Steric exclusion and wrapping of the excluded DNA strand occurs along discrete external binding paths during MCM helicase unwinding. Nucleic Acids Res. 2011, 39, 6585–6595. [Google Scholar] [CrossRef] [PubMed]
- Kasiviswanathan, R.; Shin, J.H.; Kelman, Z. DNA Binding by the Methanothermobacter thermautotrophicus Cdc6 protein is inhibited by the Minichromosome maintenance helicase. J. Bacteriol. 2006, 188, 4577–4580. [Google Scholar] [CrossRef] [PubMed]
- Georgescu, R.; Yuan, Z.; Bai, L.; de Luna Almeida Santos, R.; Sun, J.; Zhang, D.; Yurieva, O.; Li, H.; O’Donnell, M.E. Structure of eukaryotic CMG helicase at a replication fork and implications to replisome architecture and origin initiation. Proc. Natl. Acad. Sci. USA 2017, 114, E697–E706. [Google Scholar] [CrossRef] [PubMed]
- Eickhoff, P.; Kose, H.B.; Martino, F.; Petojevic, T.; Abid Ali, F.; Locke, J.; Tamberg, N.; Nans, A.; Berger, J.M.; Botchan, M.R.; et al. Molecular basis for ATP-hydrolysis-driven DNA translocation by the CMG helicase of the eukaryotic replisome. Cell Rep. 2019, 28, 2673–2688. [Google Scholar] [CrossRef]
- Baretić, D.; Jenkyn-Bedford, M.; Aria, V.; Cannone, G.; Skehel, M.; Yeeles, J.T.P. Cryo-EM structure of the fork protection complex bound to CMG at a replication fork. Mol. Cell. 2020, 78, 926–940.e13. [Google Scholar] [CrossRef]
- Rzechorzek, N.J.; Hardwick, S.W.; Jatikusumo, V.A.; Chirgadze, D.Y.; Pellegrini, L. CryoEM structures of human CMG-ATPγS-DNA and CMG-AND-1 complexes. Nucleic Acids Res. 2020, 48, 6980–6995. [Google Scholar] [CrossRef]
- Noguchi, Y.; Yuan, Z.; Bai, L.; Schneider, S.; Zhao, G.; Stillman, B.; Speck, C.; Li, H. Cryo-EM structure of Mcm2-7 double hexamer on DNA suggests a lagging-strand DNA extrusion model. Proc. Natl. Acad. Sci. USA 2017, 114, E9529–E9538. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Onesti, S. The MCM complex: (Just) a replicative helicase? Biochem. Soc. Trans. 2008, 36, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Graham, B.W.; Tao, Y.; Dodge, K.L.; Thaxton, C.T.; Olaso, D.; Young, N.L.; Marshall, A.G.; Trakselis, M.A. DNA interactions probed by hydrogen-deuterium exchange (HDX) fourier transform ion cyclotron resonance mass spectrometry confirm external binding sites on the minichromosomal maintenance (MCM) helicase. J. Biol. Chem. 2016, 291, 12467–12480. [Google Scholar] [CrossRef]
- Graham, B.W.; Bougoulias, M.E.; Dodge, K.L.; Thaxton, C.T.; Olaso, D.; Tao, Y.; Young, N.L.; Marshall, A.G.; Trakselis, M.A. Control of hexamerization, assembly, and excluded strand specificity for the sulfolobus solfataricus MCM helicase. Biochemistry 2018, 57, 5672–5682. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohammed Khalid, A.A.; Parisse, P.; Medagli, B.; Onesti, S.; Casalis, L. Atomic Force Microscopy Investigation of the Interactions between the MCM Helicase and DNA. Materials 2021, 14, 687. https://doi.org/10.3390/ma14030687
Mohammed Khalid AA, Parisse P, Medagli B, Onesti S, Casalis L. Atomic Force Microscopy Investigation of the Interactions between the MCM Helicase and DNA. Materials. 2021; 14(3):687. https://doi.org/10.3390/ma14030687
Chicago/Turabian StyleMohammed Khalid, Amna Abdalla, Pietro Parisse, Barbara Medagli, Silvia Onesti, and Loredana Casalis. 2021. "Atomic Force Microscopy Investigation of the Interactions between the MCM Helicase and DNA" Materials 14, no. 3: 687. https://doi.org/10.3390/ma14030687
APA StyleMohammed Khalid, A. A., Parisse, P., Medagli, B., Onesti, S., & Casalis, L. (2021). Atomic Force Microscopy Investigation of the Interactions between the MCM Helicase and DNA. Materials, 14(3), 687. https://doi.org/10.3390/ma14030687