
Biohydrogen Production from Lignocellulosic Biomass: Technology and Sustainability



Abstract
:1. Introduction
Aim of the Paper
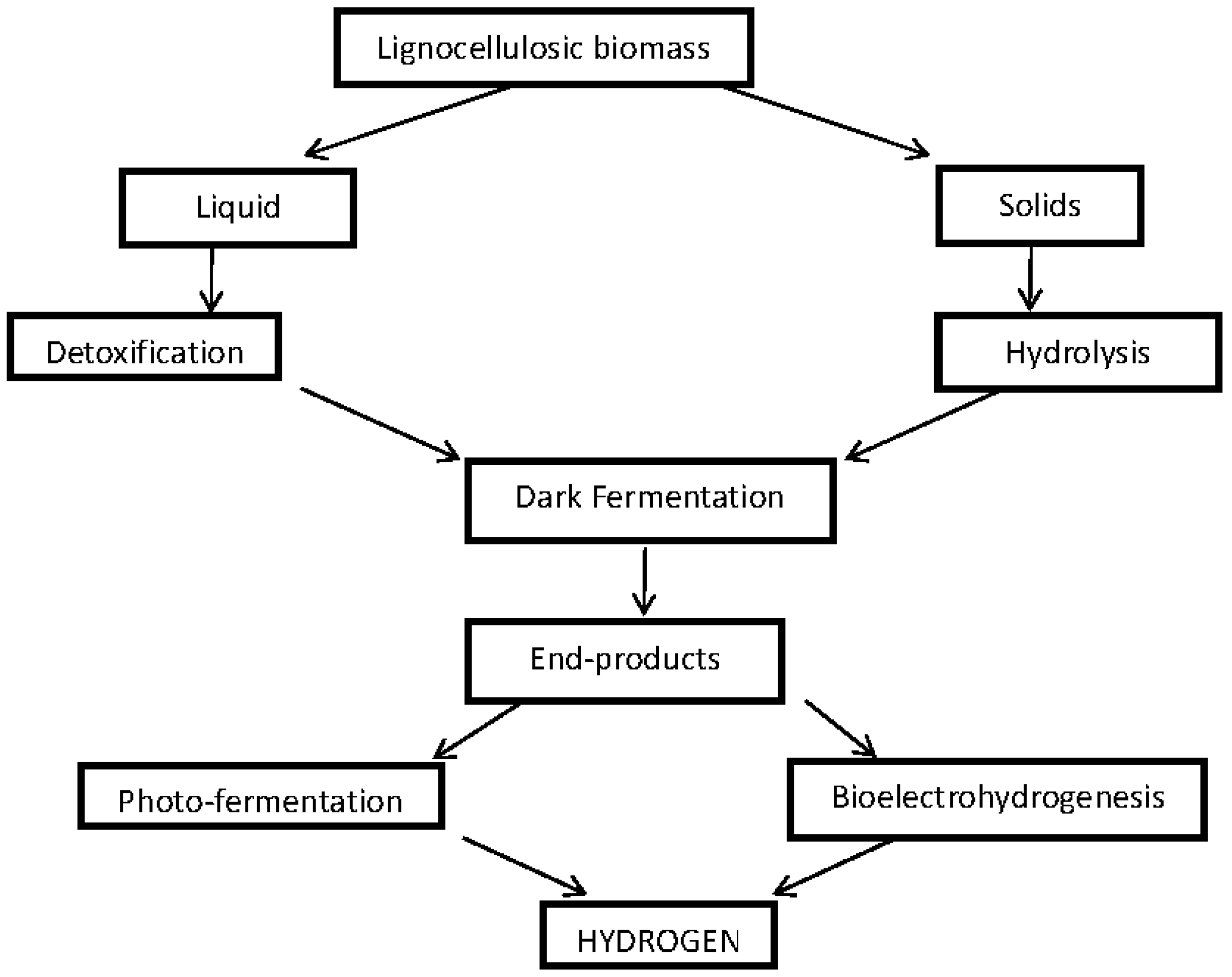
2. Feed Stock for Biohydrogen Production
3. Technology
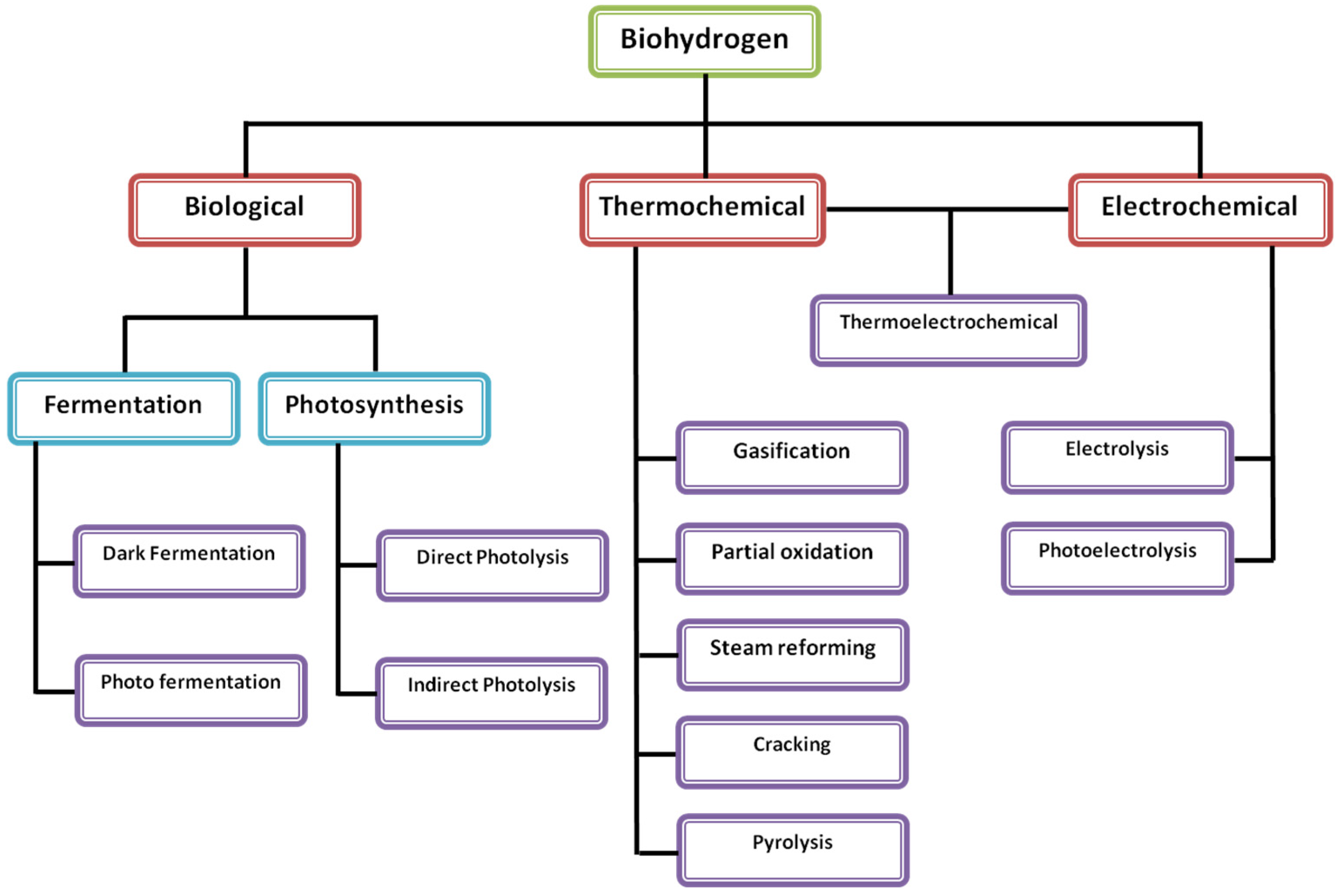
3.1. Biohydrogen Production Systems
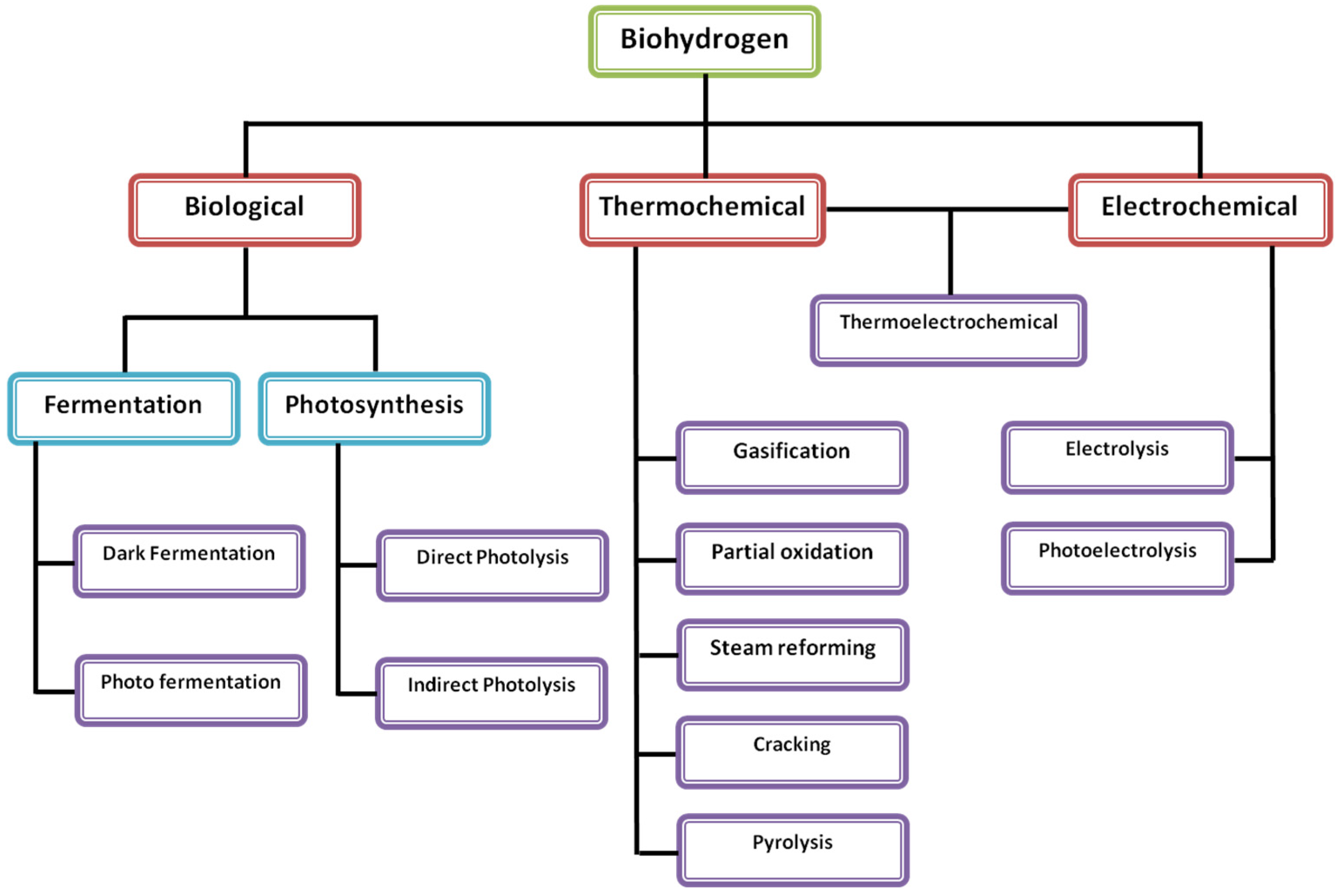
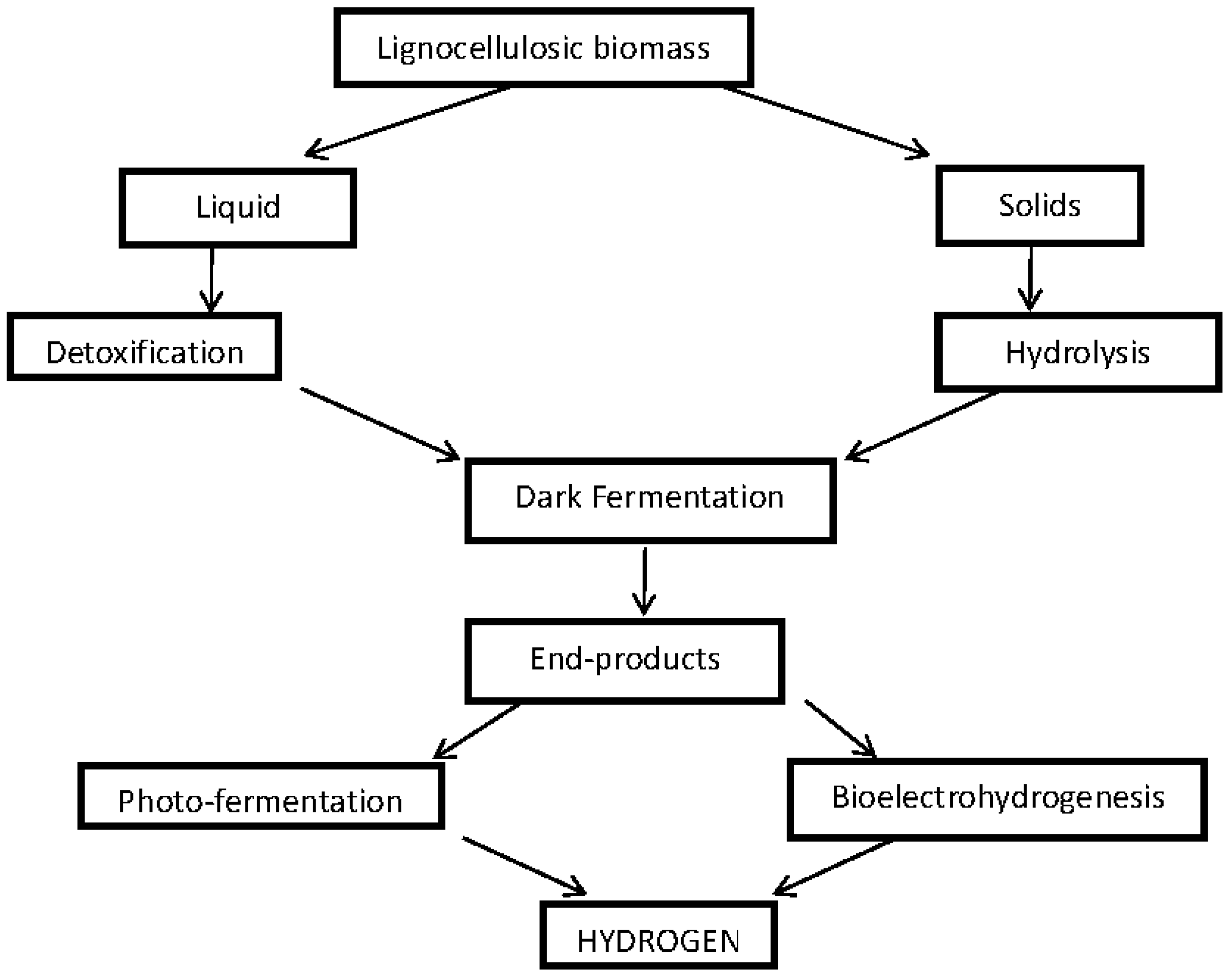
3.1.1. Dark/Anaerobic Fermentation
3.1.2. Photo Fermentation
3.1.3. Combined Biotechnologies
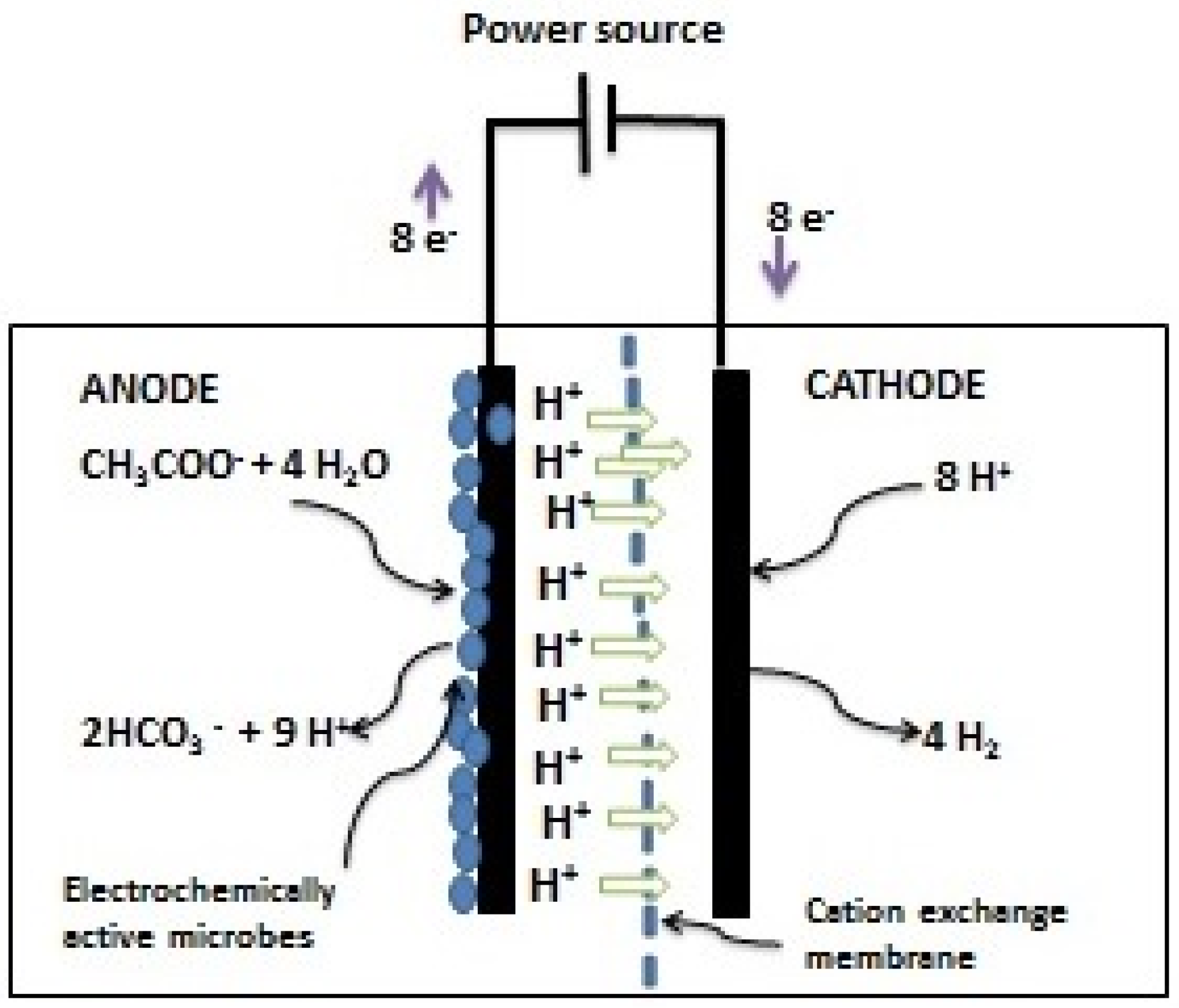
3.1.4. Bioelectrochemical Production


{kind=link}
{kind=link}
{kind=link}
| Production Routes | Main Reaction | H2 Production Rates (mmol/h·L) | Remark |
|---|---|---|---|
| Direct photolysis | 2H2O + “light energy” → 2H2 + O2 | 0.07 | Similar to the processes found in plants and algal photosynthesis. |
| Photo fermentation | C6H12O6 + 6H2O + “light energy” → 12H2 + 6CO2, ΔG0 = +3.2 kJ | 145–160 | Bacteria evolve molecular H2 catalyzed by nitrogenase under N-deficient conditions using light energy and reduced compounds (organic acids). |
| Dark fermentation | Pyruvate + CoA → acetyl-CoA + formate OR Pyruvate + CoA + 2Fd(ox) → Acetyl-CoA + CO2 + 2Fd (red) | 77 | H2 is produced by anaerobic bacteria, grown in the dark on carbohydrate rich substrate. |
| Substrate | Concentration (g/L) | Applied Voltage (V) | MEC Volume (mL) | Hydrogen Production Rate (m3 H2/m3/day) | Reference |
|---|---|---|---|---|---|
| A de-oiled refinery wastewater | 04–1 | 0.7 | 5 | 79% (Hydrogen production based on COD removal) | [60] |
| Sodium Acetate | 1 | 0.6 | 18 | 2.0 | [61] |
| Glucose | 2 | 0.6 | 26 | 0.25 ± 0.03 | [62] |
| Glucose | 2 | 0.8 | 26 | 0.37 ± 0.04 | [62] |
| Fermentation effluent | 1 | 0.6 | 26 | 1.41 | [63] |
| Sodium Acetate | 1 | 0.6 | 28 | 1.99 ± 0.02 | [64] |
| Sodium Acetate | 1 | 0.8 | 28 | 3.12 ± 0.002 | [64] |
| Sodium Acetate | 1 | 0.5 | 28 | 1.7 | [65] |
| Glucose | 1 | 0.5 | 28 | 0.83 ± 0.18 | [66] |
| Glucose | 1 | 0.9 | 28 | 1.87 ± 0.30 | [66] |
| Potato wastewater | 1.9–2.5 (COD) | 0.9 | 28 | 0.74 | [67] |
| Swine wastewater | 2 (COD) | 0.5 | 28 | 0.9–1.0 | [68] |
| Sodium Acetate | 1 | 0.6 | 48 | 0.76 | [65] |
| Sodium Acetate | 1 | 0.7 | 76 | - | [69] |
| Sodium Acetate | 1 | 0.8 | 240 | 0.0231 ± 0.003 | [70] |
| Sodium Acetate | 1 | 1 | 400 | 1.58 | [71] |
| Sodium Acetate | 2 | 0.6 | 500 | 0.53 | [72] |
| Winery wastewater | 8 | 0.9 | 1000 Lt | 0.19 ± 0.04 | [58] |
| Sodium Acetate | 1 | 0.5 | 6600 | 0.02 | [56] |
3.2. Microbiology of Biohydrogen Production
3.3. Limiting Factors in Biohydrogen Production Systems
3.4. Role of Metabolic Engineering
4. Kinetic Models for Hydrogen Production by Fermentation
5. Sustainability and Life Cycle Assessment
| Reactor | Feed Stock | Maximum H2 Yield | Reference |
|---|---|---|---|
| Fermentation | |||
| Dark fermentation | |||
| CSTR | Starch | 0.52 L/h/L and 13.2 mmol H2/g total sugar | [113] |
| Batch | Glycerol | 0.41 mol H2/mol glycerol | [114] |
| FBR | Sucrose | 4.26 mol H2/mol sucrose | [115] |
| Batch | Food waste | 593 mL H2/g carbohydrate | [116] |
| Fed-batch | Swine manure | 18.7 × 10−3 g H2 per g TVS | [117] |
| Batch | Sucrose | 4.3 mol H2/mol sucrose | [118] |
| Batch | Fructose, sorbitol, glucose | 1.27, 1.46 and 1.51 mol H2/substrate | [119] |
| Fed-batch | Starch, glucose | 465 mL H2/g starch, 3.1 mol H2/mol glucose | [120] |
| Batch | Food waste | 39.14 mL H2/g food waste (219.91 mL H2/VSadded) | [121] |
| Batch | Crude Glycerol | 64.24 mmol H2/L and 5.74 mmol H2/g COD consumed | [122] |
| Batch | Distillery wastewaters | 1 L H2/L medium | [123] |
| Batch | Cheese whey | 94.2 L H2/kgvs | [124] |
| Batch | Water hyacinth (leaves and stems) | 76.7 mL H2/g TVS was obtained at 20 g/L of water hyacinth | [125] |
| Batch | waste ground wheat solution | SHPR = 25.7 mL H2/g cells/h | [126] |
| Photo fermentation | |||
| CSTR | Sucrose | 5.81 mol H2/mol hexose | [127] |
| Fed-batch operation | Wheat starch | 201 mL H2 g/L starch | [128] |
| Batch | Molasses | 0.50 mmol H2/Lc h | [129] |
| Batch | Beet molasses | 10.5 mol H2/mol sucrose | [52] |
| Batch | Black strap | 8 mol H2/mol sucrose | [52] |
| Batch | Sucrose | 14 mol H2/mol sucrose | [52] |
| Batch | Ground wheat starch | 46 mL H2/g biomass/h, 1.23 mol H2/mol glucose | [51] |
| Batch | lignocellulose-derived organic acids | 7 mL H2/mL of the fermentation effluent | [130] |
| Photosynthesis | |||
| Direct Photolysis | |||
| Batch | Lactate | 0.07 mmol H2 (l × h) or 54 mL/h·g dry weight | [131] |
| Indirect Photolysis | |||
| Batch | arabinose and xylose | 14.55 mmol/g (arabinose); 13.73 mmol/g (xylose) | [132] |
| Thermochemical | |||
| Gasification | |||
| Continuous supercritical water gasification | glucose | 10.5–11.2 mol/mol glucose | [133] |
| Partial Oxidation | |||
| Batch | municipal sludge | Not reported the amount | [134] |
| Steam reforming | |||
| molten carbonate fuel cell (MCFC) system | ethanol | 5 mol H2/mol fed ethanol | [135] |
| Cracking | |||
| fixed-bed quartz micro reactor | Methane | 500 µmoles/min | [136] |
| Pyrolysis | |||
| stainless steel tank reactor | Biomass (redwood sawdust; cole stalk and rice husk) feed | 65.39 g/Kg biomass for redwood sawdust; 40.0 g/Kg biomass for cole stalk and rice husk | [137] |
| Thermoelectrochemical | |||
| membrane electrode assembly | sulfur dioxide | 0.4 A/cm2 at 0.835 V (H2 production rate did not reported) | [138] |
| membrane electrode assembly | anhydrous hydrogen bromide | 2.0 A/cm2 at 1.91 V (H2 production rate did not reported) | [138] |
| Electrochemical | |||
| Electrolysis | |||
| The BiOx−TiO2 electrode and stainless steel (SS, Hastelloy C-22) were used as an anode and a cathode in the electrochemical system, respectively | arsenite (As(III)) | 9.4 µmoles/min | [139] |
| Photoelectrolysis | |||
| The TiO2(ns) was prepared in the form of a sol-gel | photoelectrode system TiO2(ns)–VO2 | 6 L·h−1·m−2 for the TiO2(ns); 13.0 L·h−1·m−2 for the TiO2(ns)–VO2 photoelectrode | [140] |
6. Future Directions and Perspectives
7. Conclusions
Author Contributions
Conflicts of Interest
References
- Prasad, S.; Singh, A.; Joshi, H.C. Ethanol as an alternative fuel from agricultural, industrial and urban residues. Resour. Conserv. Recycl. 2007, 50, 1–39. [Google Scholar] [CrossRef]
- Prasad, S.; Singh, A.; Joshi, H.C. Ethanol production from sweet sorghum syrup for utilization as automotive fuel in India. Energy Fuels 2007, 21, 2415–2420. [Google Scholar] [CrossRef]
- Pant, D.; Van Bogaert, G.; Diels, L.; Vanbroekhoven, K. A review of the substrates used in microbial fuel cells (MFCs) for sustainable energy production. Bioresour. Technol. 2010, 101, 1533–1543. [Google Scholar] [CrossRef] [PubMed]
- Pant, D.; Singh, A.; van Bogaert, G.; Gallego, Y.A.; Diels, L.; Vanbroekhoven, K. An introduction to the life cycle assessment (LCA) of bioelectrochemical systems (BES) for sustainable energy and product generation: Relevance and key aspects. Renew. Sustain. Energy Rev. 2011, 15, 1305–1313. [Google Scholar] [CrossRef]
- Singh, A.; Smyth, B.M.; Murphy, J.D. A biofuel strategy for Ireland with an emphasis on production of biomethane and minimization of land-take. Renew. Sustain. Energy Rev. 2010, 14, 277–288. [Google Scholar] [CrossRef]
- Singh, A.; Pant, D.; Korres, N.E.; Nizami, A.; Prasad, S.; Murphy, J.D. Key issues in life cycle assessment of ethanol production from lignocellulosic biomass: Challenges and perspectives. Bioresour. Technol. 2010, 101, 5003–5012. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Nigam, P.S.; Murphy, J.D. Renewable fuels from algae: An answer to debatable land based fuels. Bioresour. Technol. 2011, 102, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Hallenbeck, P.C.; Ghosh, D. Advances in fermentative biohydrogen production: The way forward? Trends Biotechnol. 2009, 27, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Ball, M.; Wietschel, M. The future of hydrogen—Opportunities and challenges. Int. J. Hydrog. Energy 2009, 34, 615–627. [Google Scholar] [CrossRef]
- Perera, K.R.J.; Ketheesan, B.; Gadhamshetty, V.; Nirmalakhandan, N. Fermentative biohydrogen production: Evaluation of net energy gain. Int. J. Hydrog. Energy 2010, 35, 12224–12233. [Google Scholar] [CrossRef]
- Kovacs, K.; Maroti, G.; Rakhely, G. A novel approach for biohydrogen production. Int. J. Hydrog. Energy 2006, 31, 1460–1468. [Google Scholar] [CrossRef]
- Ren, N.; Wang, A.; Cao, G.; Xu, J.; Gao, L. Bioconversion of lignocellulosic biomass to hydrogen: Potential and challenges. Biotechnol. Adv. 2009, 27, 1051–1060. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Das, D. Enhancement of hydrogen production by Enterobacter cloacae IIT-BT 08. Process Biochem. 2000, 35, 589–593. [Google Scholar] [CrossRef]
- Chen, C.C.; Lin, C.Y.; Chang, J.S. Kinetics of hydrogen production with continuous anaerobic cultures utilizing sucrose as the limiting substrate. Appl. Microbiol. Biotechnol. 2001, 57, 56–64. [Google Scholar] [PubMed]
- Ginkel, S.V.; Sung, S.; Lay, J.J. Biohydrogen production as a function of pH and substrate concentration. Environ. Sci. Technol. 2001, 35, 4726–4730. [Google Scholar] [CrossRef] [PubMed]
- Xing, D.; Ren, N.; Li, Q.; Lin, M.; Wang, A.; Zhao, L. Ethanoligenens harbinense gen. nov., sp. nov., isolated from molasses wastewater. Int. J. Syst. Evolut. Microbiol. 2006, 56, 755–760. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhou, Q.; Li, F. Avoiding propionic acid accumulation in the anaerobic process for biohydrogen production. Biomass Bioenergy 2006, 30, 177–182. [Google Scholar] [CrossRef]
- Ren, N.; Cao, G.; Guo, W.; Wang, A.; Zhu, Y.; Liu, B.; Xu, J. Biological hydrogen production from corn stover by moderately thermophile Thermoanaerobacterium thermosaccharolyticum W16. Int. J. Hydrog. Energy 2010, 35, 2708–2712. [Google Scholar] [CrossRef]
- De Vrije, T.; de Haas, G.; Tan, G.B.; Keijsers, E.R.; Claassen, P.A.M. Pretreatment of Miscanthus for hydrogen production by Thermotoga elfii. Int. J. Hydrog. Energy 2002, 27, 1381–1390. [Google Scholar] [CrossRef]
- Ntaikou, I.; Gavala, H.N.; Kornaros, M.; Lyberatos, G. Hydrogen production from sugars and sweet sorghum biomass using Ruminococcus albus. Int. J. Hydrog. Energy 2008, 33, 1153–1163. [Google Scholar] [CrossRef]
- Kapdan, I.K.; Kargi, F. Bio-hydrogen production from waste materials. Enzym. Microb. Technol. 2006, 38, 569–582. [Google Scholar] [CrossRef]
- Kalinci, Y.; Hepbasli, A.; Dincer, I. Biomass-based hydrogen production: A review and analysis. Int. J. Hydrog. Energy 2009, 34, 8799–8817. [Google Scholar] [CrossRef]
- Kraemer, J.T.; Bagley, D.M. Improving the yield from fermentative hydrogen production. Biotechnol. Lett. 2007, 29, 685–695. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wan, W. Factors influencing fermentative hydrogen production: A review. Int. J. Hydrog. Energy 2009, 34, 799–811. [Google Scholar] [CrossRef]
- Cournac, L.; Sarma, P.M.; Fontecave, M. Biohydrogen: From Basic Concepts to Technology. Int. J. Hydrog. Energy 2010, 35, 10638. [Google Scholar] [CrossRef]
- Turner, J.; Sverdrup, G.; Mann, M.K.; Maness, P.C.; Kroposki, B.; Ghirardi, M.; Evans, R.J.; Blake, D. Renewable hydrogen production. Int. J. Energy Res. 2008, 32, 379–407. [Google Scholar] [CrossRef]
- Bartacek, J.; Zabranska, J.; Lens, P.N. Developments and constraints in fermentative hydrogen production. Biofuels Bioprod. Biorefining 2007, 1, 201–214. [Google Scholar] [CrossRef]
- Nielsen, A.T.; Amandusson, H.; Bjorklund, R.; Dannetun, H.; Ejlertsson, J.; Ekedahl, L.G.; Lundström, I.; Svensson, B.H. Hydrogen production from organic waste. Int. J. Hydrog. Energy 2001, 26, 547–550. [Google Scholar] [CrossRef]
- Kim, S.H.; Shin, H.S. Effects of base-pretreatment on continuous enriched culture for hydrogen production from food waste. Int. J. Hydrog. Energy 2008, 33, 5266–5274. [Google Scholar] [CrossRef]
- Kim, D.H.; Kim, S.H.; Shin, H.S. Hydrogen fermentation of food waste without inoculum addition. Enzyme Microb. Technol. 2009, 45, 181–187. [Google Scholar] [CrossRef]
- Ren, N.; Li, J.; Li, B.; Wang, Y.; Liu, S. Biohydrogen production from molasses by anaerobic fermentation with a pilot-scale bioreactor system. Int. J. Hydrog. Energy 2006, 31, 2147–2157. [Google Scholar] [CrossRef]
- Li, J.; Li, B.; Zhu, G.; Ren, N.; Bo, L.; He, J. Hydrogen production from diluted molasses by anaerobic hydrogen producing bacteria in an anaerobic baffled reactor (ABR). Int. J. Hydrog. Energy 2007, 32, 3274–3283. [Google Scholar] [CrossRef]
- Yang, P.; Zhang, R.; Mcgarvey, J.; Benemann, J. Biohydrogen production from cheese processing wastewater by anaerobic fermentation using mixed microbial communities. Int. J. Hydrog. Energy 2007, 32, 4761–4771. [Google Scholar] [CrossRef]
- Azbar, N.; Çetinkaya Dokgöz, F.T.; Keskin, T.; Korkmaz, K.S.; Syed, H.M. Continuous fermentative hydrogen production from cheese whey wastewater under thermophilic anaerobic conditions. Int. J. Hydrog. Energy 2009, 34, 7441–7447. [Google Scholar] [CrossRef]
- Lay, J.J. Modeling and optimization of anaerobic digested sludge converting starch to hydrogen. Biotechnol. Bioeng. 2000, 63, 269–278. [Google Scholar] [CrossRef]
- Ueno, Y.; Haruta, S.; Ishii, M.; Igarashi, Y. Microbial community in anaerobic hydrogen-producing microflora enriched from sludge compost. Appl. Microbiol. Biotechnol. 2001, 57, 555–562. [Google Scholar] [PubMed]
- Lin, C.Y.; Cheng, C.H. Fermentative hydrogen production from xylose using anaerobic mixed microflora. Int. J. Hydrog. Energy 2006, 31, 832–840. [Google Scholar] [CrossRef]
- Hussy, I.; Hawkes, F.R.; Dinsdale, R.; Hawkes, D.L. Continuous fermentative hydrogen production from sucrose and sugarbeet. Int. J. Hydrog. Energy 2005, 30, 471–483. [Google Scholar] [CrossRef]
- Wang, C.H.; Lin, P.J.; Chang, J.S. Fermentative conversion of sucrose and pineapple waste into hydrogen gas in phosphate-buffered culture seeded with municipal sewage sludge. Process Biochem. 2006, 41, 1353–1358. [Google Scholar] [CrossRef]
- Fan, K.S.; Kan, N.R.; Lay, J.J. Effect of hydraulic retention time on anaerobic hydrogenesis in CSTR. Bioresour. Technol. 2006, 97, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, F.R.; Dinsdale, R.; Hawkes, D.L.; Hussy, I. Sustainable fermentative hydrogen production: Challenges for process optimisation. Int. J. Hydrog. Energy 2002, 27, 1339–1347. [Google Scholar] [CrossRef]
- Hawkes, F.R.; Hussy, I.; Kyazze, G.; Dinsdale, R.; Hawkes, D.L. Continuous dark fermentative hydrogen production by mesophilic microflora: Principles and progress. Int. J. Hydrog. Energy 2007, 32, 172–184. [Google Scholar] [CrossRef]
- Varanasi, J.L.; Roy, S.; Pandit, S.; Das, D. Improvement of energy recovery from cellobiose by thermophillic dark fermentative hydrogen production followed by microbial fuel cell. Int. J. Hydrog. Energy 2015, 40, 8311–8321. [Google Scholar] [CrossRef]
- Angenent, L.T.; Karim, K.; Al-Dahhan, M.H.; Wrenn, B.A.; Domíguez-Espinosa, R. Production of bioenergy and biochemicals from industrial and agricultural wastewater. Trends Biotechnol. 2004, 22, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Levin, D. Biohydrogen production: Prospects and limitations to practical application. Int. J. Hydrog. Energy 2004, 29, 173–185. [Google Scholar] [CrossRef]
- Dipasquale, L.; D’Ippolito, G.; Fontana, A. Capnophilic lactic fermentation and hydrogen synthesis by Thermotoga neapolitana: An unexpected deviation from the dark fermentation model. Int. J. Hydrog. Energy 2014, 39, 4857–4862. [Google Scholar] [CrossRef]
- Pradhan, N.; Dipasquale, L.; D’Ippolito, G.; Panico, A.; Lens, P.N.L.; Esposito, G.; Fontana, A. Hydrogen production by the thermophilic bacterium thermotoga neapolitana. Int. J. Mol. Sci. 2015, 16, 12578–12600. [Google Scholar] [CrossRef] [PubMed]
- Basak, N.; Das, D. The prospect of purple non-sulfur (PNS) photosynthetic bacteria for hydrogen production: The present state of the art. World J. Microbiol. Biotechnol. 2007, 23, 31–42. [Google Scholar] [CrossRef]
- Redwood, M.D.; Paterson-Beedle, M.; MacAskie, L.E. Integrating dark and light bio-hydrogen production strategies: Towards the hydrogen economy. Rev. Environ. Sci. Biotechnol. 2009, 8, 149–185. [Google Scholar] [CrossRef]
- Holladay, J.D.; Hu, J.; King, D.L.; Wang, Y. An overview of hydrogen production technologies. Catal. Today 2009, 139, 244–260. [Google Scholar] [CrossRef]
- Kapdan, I.K.; Kargi, F.; Oztekin, R.; Argun, H. Bio-hydrogen production from acid hydrolyzed wheat starch by photo-fermentation using different Rhodobacter sp. Int. J. Hydrog. Energy 2009, 34, 2201–2207. [Google Scholar] [CrossRef]
- Keskin, T.; Hallenbeck, P.C. Hydrogen production from sugar industry wastes using single-stage photofermentation. Bioresour. Technol. 2012, 112, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Nath, K.; Das, D. Improvement of fermentative hydrogen production: Various approaches. Appl. Microbiol. Biotechnol. 2004, 65, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.F.; Ren, N.Q.; Xie, G.J.; Ding, J.; Guo, W.Q.; Xing, D.F. Enhanced bio-hydrogen production by the combination of dark- and photo-fermentation in batch culture. Bioresour. Technol. 2010, 101, 5325–5329. [Google Scholar] [CrossRef] [PubMed]
- Logan, B.E. Microbial Fuel Cell; Wiley & Sons, Inc.: Hoboken, NJ, USA, 2008. [Google Scholar]
- Rozendal, R.; Hamelers, H.; Euverink, G.; Metz, S.; Buisman, C. Principle and perspectives of hydrogen production through biocatalyzed electrolysis. Int. J. Hydrog. Energy 2006, 31, 1632–1640. [Google Scholar] [CrossRef]
- Manish, S.; Banerjee, R. Comparison of biohydrogen production processes. Int. J. Hydrog. Energy 2008, 33, 279–286. [Google Scholar] [CrossRef]
- Cusick, R.D.; Bryan, B.; Parker, D.S.; Merrill, M.D.; Mehanna, M.; Kiely, P.D.; Liu, G.; Logan, B.E. Performance of a pilot-scale continuous flow microbial electrolysis cell fed winery wastewater. Appl. Microbiol. Biotechnol. 2011, 89, 2053–2063. [Google Scholar] [CrossRef] [PubMed]
- Kadier, A.; Simayi, Y.; Kalil, M.S.; Abdeshahian, P.; Hamid, A.A. A review of the substrates used in microbial electrolysis cells (MECs) for producing sustainable and clean hydrogen gas. Renew. Energy 2014, 71, 466–472. [Google Scholar] [CrossRef]
- Ren, L.; Siegert, M.; Ivanov, I.; Pisciotta, J.M.; Logan, B.E. Treatability studies on different refinery wastewater samples using high-throughput microbial electrolysis cells (MECs). Bioresour. Technol. 2013, 136, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Fan, Y.; Liu, H. Hydrogen production in single-chamber tubular microbial electrolysis cells using non-precious-metal catalysts. Int. J. Hydrog. Energy 2009, 34, 8535–8542. [Google Scholar] [CrossRef]
- Lu, L.; Xing, D.; Ren, N.; Logan, B.E. Syntrophic interactions drive the hydrogen production from glucose at low temperature in microbial electrolysis cells. Bioresour. Technol. 2012, 124, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Ren, N.; Xing, D.; Logan, B.E. Hydrogen production with effluent from an ethanol-H2-coproducing fermentation reactor using a single-chamber microbial electrolysis cell. Biosens. Bioelectron. 2009, 24, 3055–3060. [Google Scholar] [CrossRef] [PubMed]
- Call, D.; Logan, B.E. Hydrogen production in a single chamber microbial electrolysis cell lacking a membrane. Environ. Sci. Technol. 2008, 42, 3401–3406. [Google Scholar] [CrossRef] [PubMed]
- Call, D.F.; Merrill, M.D.; Logan, B.E. High surface area stainless steel brushes as cathodes in microbial electrolysis cells. Environ. Sci. Technol. 2009, 43, 2179–2183. [Google Scholar] [CrossRef] [PubMed]
- Selembo, P.A.; Perez, J.M.; Lloyd, W.A.; Logan, B.E. High hydrogen production from glycerol or glucose by electrohydrogenesis using microbial electrolysis cells. Int. J. Hydrog. Energy 2009, 34, 5373–5381. [Google Scholar] [CrossRef]
- Wang, X.; Cheng, S.; Feng, Y.; Merrill, M.D.; Saito, T.; Logan, B.E. Use of carbon mesh anodes and the effect of different pretreatment methods on power production in microbial fuel cells. Environ. Sci. Technol. 2009, 43, 6870–6874. [Google Scholar] [CrossRef] [PubMed]
- Wagner, R.C.; Regan, J.M.; Oh, S.-E.; Zuo, Y.; Logan, B.E. Hydrogen and methane production from swine wastewater using microbial electrolysis cells. Water Res. 2009, 43, 1480–1488. [Google Scholar] [CrossRef] [PubMed]
- Pisciotta, J.M.; Zaybak, Z.; Call, D.F.; Nam, J.Y.; Logan, B.E. Enrichment of microbial electrolysis cell biocathodes from sediment microbial fuel cell bioanodes. Appl. Environ. Microbiol. 2012, 78, 5212–5219. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Wen, Z.; Ci, S.; Chen, J.; He, Z. Carbon/iron-based nanorod catalysts for hydrogen production in microbial electrolysis cells. Nano Energy 2012, 1, 751–756. [Google Scholar] [CrossRef]
- Guo, K.; Tang, X.; Du, Z.; Li, H. Hydrogen production from acetate in a cathode-on-top single-chamber microbial electrolysis cell with a mipor cathode. Biochem. Eng. J. 2010, 51, 48–52. [Google Scholar] [CrossRef]
- Hu, H.; Fan, Y.; Liu, H. Hydrogen production using single-chamber membrane-free microbial electrolysis cells. Water Res. 2008, 42, 4172–4178. [Google Scholar] [CrossRef] [PubMed]
- Van Groenestijn, J.W.; Geelhoed, J.S.; Goorissen, H.P.; Meesters, K.P.; Stams, A.J.; Claassen, P.A. Performance and population analysis of a non-sterile trickle bed reactor inoculated with caldicellulosiruptor saccharolyticus, a thermophilic hydrogen producer. Biotechnol. Bioeng. 2009, 102, 1361–1367. [Google Scholar] [CrossRef] [PubMed]
- Kleerebezem, R.; van Loosdrecht, M.C. Mixed culture biotechnology for bioenergy production. Curr. Opin. Biotechnol. 2007, 18, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Kengen, S.W.M.; Goorissen, H.P.; Verhaart, M.; Stams, A.J.M.; van Niel, E.W.J.; Claassen, P.A.M. Biological Hydrogen Production by Anaerobic Microorganisms. In Biofuels; John Wiley & Sons, Ltd.: Chichester, UK, 2009; pp. 197–221. [Google Scholar]
- Kotsopoulos, T.A.; Zeng, R.J.; Angelidaki, I. Biohydrogen production in granular up-flow anaerobic sludge blanket (UASB) reactors with mixed cultures under hyper-thermophilic temperature (70 C). Biotechnol. Bioeng. 2006, 94, 296–302. [Google Scholar] [CrossRef] [PubMed]
- De Vrije, T.; Bakker, R.R.; Budde, M.A.; Lai, M.H.; Mars, A.E.; Claassen, P.A. Efficient hydrogen production from the lignocellulosic energy crop Miscanthus by the extreme thermophilic bacteria Caldicellulosiruptor saccharolyticus and Thermotoga neapolitana. Biotechnol. Biofuels 2009, 2, 12. [Google Scholar] [CrossRef] [PubMed]
- Van Niel, E.W.; Budde, M.A.; De Haas, G.; Van der Wal, F.J.; Claassen, P.A.; Stams, A.J. Distinctive properties of high hydrogen producing extreme thermophiles, Caldicellulosiruptor saccharolyticus and Thermotoga elfii. Int. J. Hydrog. Energy 2002, 27, 1391–1398. [Google Scholar] [CrossRef]
- Thauer, R.K.; Jungermann, K.; Decker, K. Energy conservation in chemotrophic anaerobic bacteria. Bacteriol. Rev. 1977, 41, 100–180. [Google Scholar] [PubMed]
- Vanginkel, S.; Oh, S.; Logan, B. Biohydrogen gas production from food processing and domestic wastewaters. Int. J. Hydrog. Energy 2005, 30, 1535–1542. [Google Scholar] [CrossRef]
- Liu, D.; Liu, D.; Zeng, R.J.; Angelidaki, I. Hydrogen and methane production from household solid waste in the two-stage fermentation process. Water Res. 2006, 40, 2230–2236. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.-E.; van Ginkel, S.; Logan, B.E. The relative effectiveness of pH control and heat treatment for enhancing biohydrogen gas production. Environ. Sci. Technol. 2003, 37, 5186–5190. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.M.; Cheng, S.S.; Wu, K.L. Behavioral study on hydrogen fermentation reactor installed with silicone rubber membrane. Int. J. Hydrog. Energy 2002, 27, 1157–1165. [Google Scholar] [CrossRef]
- Van Groenestijn, J.W.; Hazewinkel, J.H.; Nienoord, M.; Bussmann, P.J. Energy aspects of biological hydrogen production in high rate bioreactors operated in the thermophilic temperature range. Int. J. Hydrog. Energy 2002, 27, 1141–1147. [Google Scholar] [CrossRef]
- Kim, D.H.; Han, S.K.; Kim, S.H.; Shin, H.S. Effect of gas sparging on continuous fermentative hydrogen production. Int. J. Hydrog. Energy 2006, 31, 2158–2169. [Google Scholar] [CrossRef]
- Koku, H. Aspects of the metabolism of hydrogen production by Rhodobacter sphaeroides. Int. J. Hydrog. Energy 2002, 27, 1315–1329. [Google Scholar] [CrossRef]
- Beckers, L.; Masset, J.; Hamilton, C.; Delvigne, F.; Toye, D.; Crine, M.; Thonart, P.; Hiligsmann, S. Investigation of the links between mass transfer conditions, dissolved hydrogen concentration and biohydrogen production by the pure strain Clostridium butyricum CWBI1009. Biochem. Eng. J. 2015, 98, 18–28. [Google Scholar] [CrossRef]
- Mizuno, O.; Dinsdale, R.; Hawkes, F.R.; Hawkes, D.L.; Noike, T. Enhancement of hydrogen production from glucose by nitrogen gas sparging. Bioresour. Technol. 2000, 73, 59–65. [Google Scholar] [CrossRef]
- Chou, C.; Wang, C.; Huang, C.; Lay, J. Pilot study of the influence of stirring and pH on anaerobes converting high-solid organic wastes to hydrogen. Int. J. Hydrog. Energy 2008, 33, 1550–1558. [Google Scholar] [CrossRef]
- Fontes Lima, D.M.; Zaiat, M. The influence of the degree of back-mixing on hydrogen production in an anaerobic fixed-bed reactor. Int. J. Hydrog. Energy 2012, 37, 9630–9635. [Google Scholar] [CrossRef]
- Montiel-Corona, V.; Revah, S.; Morales, M. Hydrogen production by an enriched photoheterotrophic culture using dark fermentation effluent as substrate: Effect of flushing method, bicarbonate addition, and outdoor–indoor conditions. Int. J. Hydrog. Energy 2015, 40, 9096–9105. [Google Scholar] [CrossRef]
- Clark, I.C.; Zhang, R.H.; Upadhyaya, S.K. The effect of low pressure and mixing on biological hydrogen production via anaerobic fermentation. Int. J. Hydrog. Energy 2012, 37, 11504–11513. [Google Scholar] [CrossRef]
- Kim, D.-H.; Shin, H.-S.; Kim, S.-H. Enhanced H2 fermentation of organic waste by CO2 sparging. Int. J. Hydrog. Energy 2012, 37, 15563–15568. [Google Scholar] [CrossRef]
- Gómez, X.; Morán, A.; Cuetos, M.J.; Sánchez, M.E. The production of hydrogen by dark fermentation of municipal solid wastes and slaughterhouse waste: A two-phase process. J. Power Sources 2006, 157, 727–732. [Google Scholar] [CrossRef]
- Clauwaert, P.; Verstraete, W. Methanogenesis in membraneless microbial electrolysis cells. Appl. Microbiol. Biotechnol. 2009, 82, 829–836. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Xing, D.; Call, D.F.; Logan, B.E. Direct biological conversion of electrical current into methane by electromethanogenesis. Environ. Sci. Technol. 2009, 43, 3953–3958. [Google Scholar] [CrossRef] [PubMed]
- Pant, D.; van Bogaert, G.; de Smet, M.; Diels, L.; Vanbroekhoven, K. Use of novel permeable membrane and air cathodes in acetate microbial fuel cells. Electrochimica Acta 2010, 55, 7710–7716. [Google Scholar] [CrossRef]
- Selembo, P.A.; Merrill, M.D.; Logan, B.E. The use of stainless steel and nickel alloys as low-cost cathodes in microbial electrolysis cells. J. Power Sour. 2009, 190, 271–278. [Google Scholar] [CrossRef]
- Maeda, T.; Sanchez-Torres, V.; Wood, T.K. Metabolic engineering to enhance bacterial hydrogen production. Microb. Biotechnol. 2008, 1, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, K.; Kimura, T.; Sakka, K.; Ohmiya, K. Overexpression of a hydrogenase gene in Clostridium paraputrificum to enhance hydrogen gas production. FEMS Microbiol. Lett. 2005, 246, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Vardar-Schara, G.; Maeda, T.; Wood, T.K. Metabolically engineered bacteria for producing hydrogen via fermentation. Microb. Biotechnol. 2008, 1, 107–125. [Google Scholar] [CrossRef] [PubMed]
- Hallenbeck, P.C.; Ghosh, D. Improvements in fermentative biological hydrogen production through metabolic engineering. J. Environ. Manag. 2012, 95, S360–S364. [Google Scholar] [CrossRef] [PubMed]
- Ryu, M.-H.; Hull, N.C.; Gomelsky, M. Metabolic engineering of Rhodobacter sphaeroides for improved hydrogen production. Int. J. Hydrog. Energy 2014, 39, 6384–6390. [Google Scholar] [CrossRef]
- Wang, J.; Wan, W. Kinetic models for fermentative hydrogen production: A review. Int. J. Hydrog. Energy 2009, 34, 3313–3323. [Google Scholar] [CrossRef]
- Dincer, I. Hydrogen and Fuel Cell Technologies for Sustainable Future. Jordan J. Mech. Ind. Eng. 2008, 2, 1–14. [Google Scholar]
- Rathore, D.; Singh, A. Biohydrogen production from microalgae. In Biofuels Technologies Recent Developments; Gupta, V., Tuohy, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 317–333. [Google Scholar]
- Kwak, H.Y.; Lee, H.S.; Jung, J.Y.; Jeon, J.S.; Park, D.R. Exergetic and thermoeconomic analysis of a 200-kW phosphoric acid fuel cell plant. Fuel 2004, 83, 2087–2094. [Google Scholar] [CrossRef]
- Rubio Rodríguez, M.A.; de Ruyck, J.; Díaz, P.R.; Verma, V.K.; Bram, S. An LCA based indicator for evaluation of alternative energy routes. Appl. Energy 2011, 88, 630–635. [Google Scholar] [CrossRef]
- Romagnoli, F.; Blumberga, D.; Pilicka, I. Life cycle assessment of biohydrogen production in photosynthetic processes. Int. J. Hydrog. Energy 2011, 36, 7866–7871. [Google Scholar] [CrossRef]
- Djomo, S.N.; Humbert, S. Dagnija Blumberga Life cycle assessment of hydrogen produced from potato steam peels. Int. J. Hydrog. Energy 2008, 33, 3067–3072. [Google Scholar] [CrossRef]
- Djomo, S.N.; Blumberga, D. Comparative life cycle assessment of three biohydrogen pathways. Bioresour. Technol. 2011, 102, 2684–2694. [Google Scholar] [CrossRef] [PubMed]
- Ochs, D.; Wukovits, W.; Ahrer, W. Life cycle inventory analysis of biological hydrogen production by thermophilic and photo fermentation of potato steam peels (PSP). J. Clean. Prod. 2010, 18, S88–S94. [Google Scholar] [CrossRef]
- Chen, S.D.; Lo, Y.C.; Lee, K.S.; Huang, T.I.; Chang, J.S. Sequencing batch reactor enhances bacterial hydrolysis of starch promoting continuous bio-hydrogen production from starch feedstock. Int. J. Hydrog. Energy 2009, 34, 8549–8557. [Google Scholar] [CrossRef]
- Seifert, K.; Waligorska, M.; Wojtowski, M.; Laniecki, M. Hydrogen generation from glycerol in batch fermentation process. Int. J. Hydrog. Energy 2009, 34, 3671–3678. [Google Scholar] [CrossRef]
- Lin, C.N.; Wu, S.Y.; Chang, J.S.; Chang, J.S. Biohydrogen production in a three-phase fluidized bed bioreactor using sewage sludge immobilized by ethylene-vinyl acetate copolymer. Bioresour. Technol. 2009, 100, 3298–3301. [Google Scholar] [CrossRef] [PubMed]
- Yasin Nazlina, H.M.; Aini, R.; Ismail, F.; Zulkhairi, M.; Hassan, M.A. Effect of different temperature, initial pH and substrate composition on biohydrogen production from food waste in batch fermentation. Asian J. Biotechnol. 2009, 1, 42–50. [Google Scholar] [CrossRef]
- Zhu, J.; Li, Y.; Wu, X.; Miller, C.; Chen, P.; Ruan, R. Swine manure fermentation for hydrogen production. Bioresour. Technol. 2009, 100, 5472–5477. [Google Scholar] [CrossRef] [PubMed]
- Gadhamshetty, V.; Johnson, D.C.; Nirmalakhandan, N.; Smith, G.B.; Deng, S. Feasibility of biohydrogen production at low temperatures in unbuffered reactors. Int. J. Hydrog. Energy 2009, 34, 1233–1243. [Google Scholar] [CrossRef]
- Ghosh, D.; Hallenbeck, P.C. Fermentative hydrogen yields from different sugars by batch cultures of metabolically engineered Escherichia coli DJT135. Int. J. Hydrog. Energy 2009, 34, 7979–7982. [Google Scholar] [CrossRef]
- Kargi, F.; Pamukoglu, M.Y. Dark fermentation of ground wheat starch for bio-hydrogen production by fed-batch operation. Int. J. Hydrog. Energy 2009, 34, 2940–2946. [Google Scholar] [CrossRef]
- Han, W.; Ye, M.; Zhu, A.J.; Zhao, H.T.; Li, Y.F. Batch dark fermentation from enzymatic hydrolyzed food waste for hydrogen production. Bioresour. Technol. 2015, 191, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Chookaew, T.; O-Thong, S.; Prasertsan, P. Biohydrogen production from crude glycerol by two stage of dark and photo fermentation. Int. J. Hydrog. Energy 2015, 40, 7433–7438. [Google Scholar] [CrossRef]
- Wicher, E.; Seifert, K.; Zagrodnik, R.; Pietrzyk, B.; Laniecki, M. Hydrogen gas production from distillery wastewater by dark fermentation. Int. J. Hydrog. Energy 2013, 38, 7767–7773. [Google Scholar] [CrossRef]
- Moreno, R.; Escapa, A.; Cara, J.; Carracedo, B.; Gómez, X. A two-stage process for hydrogen production from cheese whey: Integration of dark fermentation and biocatalyzed electrolysis. Int. J. Hydrog. Energy 2015, 40, 168–175. [Google Scholar] [CrossRef]
- Su, H.; Cheng, J.; Zhou, J.; Song, W.; Cen, K. Hydrogen production from water hyacinth through dark- and photo- fermentation. Int. J. Hydrog. Energy 2010, 35, 8929–8937. [Google Scholar] [CrossRef]
- Argun, H.; Kargi, F. Effects of sludge pre-treatment method on bio-hydrogen production by dark fermentation of waste ground wheat. Int. J. Hydrog. Energy 2009, 34, 8543–8548. [Google Scholar] [CrossRef]
- Laurinavichene, T.V.; Belokopytov, B.F.; Laurinavichius, K.S.; Tekucheva, D.N.; Seibert, M.; Tsygankov, A.A. Towards the integration of dark- and photo-fermentative waste treatment. 3. Potato as substrate for sequential dark fermentation and light-driven H2 production. Int. J. Hydrog. Energy 2010, 35, 8536–8543. [Google Scholar] [CrossRef]
- Ozmihci, S.; Kargi, F. Effects of starch loading rate on performance of combined fed-batch fermentation of ground wheat for bio-hydrogen production. Int. J. Hydrog. Energy 2010, 35, 1106–1111. [Google Scholar] [CrossRef]
- Avcioglu, S.G.; Ozgur, E.; Eroglu, I.; Yucel, M.; Gunduz, U. Biohydrogen production in an outdoor panel photobioreactor on dark fermentation effluent of molasses. Int. J. Hydrog. Energy 2011, 36, 11360–11368. [Google Scholar] [CrossRef]
- Zhu, Z.; Shi, J.; Zhou, Z.; Hu, F.; Bao, J. Photo-fermentation of Rhodobacter sphaeroides for hydrogen production using lignocellulose-derived organic acids. Process Biochem. 2010, 45, 1894–1898. [Google Scholar] [CrossRef]
- Francou, N.; Vignais, P.M. Hydrogen production by Rhodopseudomonas capsulata cells entrapped in carrageenan beads. Biotechnol. Lett. 1984, 6, 639–644. [Google Scholar] [CrossRef]
- Taguchi, F.; Mizukami, N.; Hasegawa, K.; Saito-Taki, T. Microbial conversion of arabinose and xylose to hydrogen by a newly isolated clostridium sp. No. 2. Can. J. Microbiol. 1994, 40, 228–233. [Google Scholar] [CrossRef]
- Susanti, R.F.; Dianningrum, L.W.; Yum, T.; Kim, Y.; Lee, B.G.; Kim, J. High-yield hydrogen production from glucose by supercritical water gasification without added catalyst. Int. J. Hydrog. Energy 2012, 37, 11677–11690. [Google Scholar] [CrossRef]
- Qinming, Z.; Shuzhong, W.; Liang, W.; Donghai, X. Catalytic Hydrogen Production from Municipal Sludge in Supercritical Water with Partial Oxidation. Chall. Power Eng. Environ. 2007, 1, 1252–1255. [Google Scholar]
- Frusteri, F.; Freni, S.; Chiodo, V.; Spadaro, L.; Bonura, G.; Cavallaro, S. Potassium improved stability of Ni/MgO in the steam reforming of ethanol for the production of hydrogen for MCFC. J. Power Sour. 2004, 132, 139–144. [Google Scholar] [CrossRef]
- Aiello, R.; Fiscus, J.E.; Loye, H.; Amiridis, M.D. Hydrogen production via the direct cracking of methane over Ni/SiO2: Catalyst deactivation and regeneration. Appl. Catal. A Gen. 2000, 192, 227–234. [Google Scholar] [CrossRef]
- Deng, W.; Jiang, H.; Wu, Y.; Fan, H.; Ji, J. Hydrogen production from biomass pyrolysis in molten alkali. AASRI Procedia 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Sivasubramanian, P.; Ramasamy, R.P.; Freire, F.J.; Holland, C.E.; Weidner, J.W. Electrochemical hydrogen production from thermochemical cycles using a proton exchange membrane electrolyzer. Int. J. Hydrog. Energy 2007, 32, 463–468. [Google Scholar] [CrossRef]
- Kim, J.; Kwon, D.; Kim, K.; Hoffmann, M.R. Electrochemical Production of Hydrogen Coupled with the Oxidation of Arsenite. Environ. Sci. Technol. 2014, 48, 2059–2066. [Google Scholar] [CrossRef] [PubMed]
- Karn, R.K.; Srivastava, O.N. On the synthesis and photochemical studies of nanostructured TiO 1 and TiO 1 admixed VO 1 photoelectrodes in regard to hydrogen production through photoelectrolysis. Int. J. Hydrog. Energy 1999, 24, 965–971. [Google Scholar] [CrossRef]
- Perlack, R.D.; Wright, L.L.; Turhollow, A.F.; Graham, R.L.; Stokes, B.J.; Erbach, D.C. Biomass as Feedstock For a Bioenergy and Bioproducts Industry: The Technical Feasibility of a Billion-Ton Annual Supply; U.S. Department of Energy: Oak Ridge, TN, USA, 2005. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, A.; Sevda, S.; Abu Reesh, I.M.; Vanbroekhoven, K.; Rathore, D.; Pant, D. Biohydrogen Production from Lignocellulosic Biomass: Technology and Sustainability. Energies 2015, 8, 13062-13080. https://doi.org/10.3390/en81112357
Singh A, Sevda S, Abu Reesh IM, Vanbroekhoven K, Rathore D, Pant D. Biohydrogen Production from Lignocellulosic Biomass: Technology and Sustainability. Energies. 2015; 8(11):13062-13080. https://doi.org/10.3390/en81112357
Chicago/Turabian StyleSingh, Anoop, Surajbhan Sevda, Ibrahim M. Abu Reesh, Karolien Vanbroekhoven, Dheeraj Rathore, and Deepak Pant. 2015. "Biohydrogen Production from Lignocellulosic Biomass: Technology and Sustainability" Energies 8, no. 11: 13062-13080. https://doi.org/10.3390/en81112357
APA StyleSingh, A., Sevda, S., Abu Reesh, I. M., Vanbroekhoven, K., Rathore, D., & Pant, D. (2015). Biohydrogen Production from Lignocellulosic Biomass: Technology and Sustainability. Energies, 8(11), 13062-13080. https://doi.org/10.3390/en81112357